
YBUF5和YBUF7菌株对人参锈腐病的防病作用及几种防御酶活性的影响
2021-09-29 04:50颜尘栋金春吉李熙英
延边大学农学学报 2021年3期
颜尘栋,金春吉,金 辉,王 薇,王 伟,李熙英*
(1.延边大学农学院,吉林 延吉 133002;2.和龙市农业科技教育中心,吉林 和龙 133500)
人参(PanaxginsengC. A. Mey )是一种名贵的中药材,含人参皂苷、氨基酸等多种成分,可增强人体的抵抗力,具有调节神经功能以及促进物质代谢等功能[1-2]。
人参人工栽培过程中常发生各种病害严重危害人参产量及品质。国内外已有描述的人参病害有40多种,其中人参锈腐病(Cylindrocarpondestructans)发生普遍而严重,一般发病率20%~30%,严重地块发病率70%以上,严重影响人参产业发展。目前,人参锈腐病的防治主要采用农业防治、化学防治等。农业防治一般效果偏低,作用较慢;化学防治常导致“3R”问题以及环境污染、药害、人畜中毒、农田生态平衡与生物多样性的破坏等一系列社会问题[3]。化学药剂对环境的污染和对人畜的危害,对环境友好安全的生物防治越来越被重视[4]。
目前,用于人参病害生物防治的微生物种类繁多,但主要集中在生防细菌[5]和生防真菌[6]的研究方面。人参锈腐病是对人参根部危害最大的 1 种土传病害,对人参产量和品质影响极大,常年发病率为24.9%~44.1%,目前仍没有较好的解决办法。近年来,用化学防治措施来防治人参锈腐病的结果远远未达到人们预想的效果。有关拮抗放线菌利用于人参病害防治方面主要集中在拮抗放线菌的分离筛选及鉴定等[7]。
众多研究证明了放线菌能促进小麦,西瓜,棉花等重要经济作物的生长。并有效抑制多种病原菌的生长[8]。经过Zhao等[9]的研究证明了用放线菌HD-087处理幼苗发现,幼苗中PAL、POD的活性大大增强。表明放线菌可以有效的抑制病原菌的生长,促进植物体内防御酶的活性并且提高了植物生长的速度。
该研究利用拮抗放线菌菌株YBUF5和YBUF7进行了田间防病试验和人参植株体内几种防御酶活性的影响研究,为拮抗放线菌菌株田间实际应用以及探明生防机理等方面提供依据。
1 材料与方法
1.1 供试菌株
供试拮抗放线菌为YBUF5菌株和YBUF7菌株,YBUF5菌株为生暗灰链霉菌(Streptomycescaniferus),对人参锈腐病菌的室内抑菌率为82.20%;YBUF7菌株为湿链霉菌(Streptomyceshumidus),对人参锈腐病菌的室内抑菌率为78.60%[10];供试病原菌为人参锈腐病菌(Cylindrocarpondestructans)由延边大学农学院植物病理研究室保管。
1.2 供试人参
2年生人参苗,由延边农科院提供。
1.3 菌悬液的制备
1) 病原菌菌悬液:取活化好的人参锈腐病原菌4培养皿,用手持式高速匀浆机机械捣碎后,加入蒸馏水定容至1 000 mL。
2) 拮抗菌菌悬液:取活化好的YBUF5和YBUF7菌株4培养皿,用手持式高速匀浆机机械捣碎后,加入蒸馏水定容至1 000 mL。
1.4 YBUF5、YBUF7及混合菌株对人参锈腐病的防病作用
该试验在延边大学农学院实验基地进行。春季移栽的人参出苗后,采用菌悬液灌根的方法进行拮抗放线菌菌株对人参锈腐病的防病试验。试验设置YBUF5菌悬液10 mL+无菌水10 mL、YBUF7菌悬液10 mL+无菌水10 mL、YBUF5和YBUF7菌悬液各5 mL+无菌水10 mL、人参锈腐病菌菌悬液10 mL+YBUF5菌悬液10 mL、人参锈腐病菌菌悬液10 mL+YBUF7菌悬液10 mL、人参锈腐病原菌悬液10 mL+YBUF5和YBUF7菌悬液各5 mL等6种处理;对照组设置为先加人参锈腐病菌菌悬液10 mL+无菌水10 mL、只加20 mL无菌水2种。每处理设置50株,2次重复。秋季,调查成活株数、病株数,并测根径、根长、单根重、产量。人参锈腐病病情分级标准参考周淑香等[11]方法。公式如下:
成活率/%=(平均成活株数/定植株数)×100
1.5 YBUF5、YBUF7及混合菌株对人参组织内POD、CAT、SOD、PAL活性的影响
试验处理同上。菌悬液处理后的第30、40、50天测定各处理组人参根茎和叶片的POD、CAT、SOD、PAL活性,具体方法参照王学奎[12]、宋喜霞[13]和赵淑玲等[14]的方法测定。
2 结果与分析
2.1 YBUF5、YBUF7及混合菌株对人参锈腐病的防病作用
由表1可见,没接锈腐病菌只接YBUF5、YBUF7及混合菌株处理的人参植株均无发病,其参苗成活率均在90%以上,并与无接菌对照间无显著性差异。说明YBUF5、YBUF7菌株及混合菌株对人参植株是安全的。在人工接锈腐病菌的条件下,YBUF5、YBUF7及混合菌株处理的人参植株成活株数显著高于只接锈腐病菌的对照,其中YBUF5菌株处理的成活率最高、病情指数最低,其防效为69.88%;其次为YBUF7处理的人参植株成活率较高、病情指数较低,其防效为60.24;混合菌株处理成活率较低,病情指数最高,其防效为51.81%(表1)。

表1 YBUF5、YBUF7及混合菌株对人参植株成活及锈腐病的防病作用
由表2可知,YBUF5、YBUF7单菌株处理的人参根茎、根长及产量等明显高于无菌对照,但YBUF5菌株和YBUF7菌株之间没有显著性差异;混合菌株与无菌对照之间也没有显著性差异。YBUF5菌株和YBUF7菌株处理的人参单根重明显优于无菌对照,其单根增产率分别为113.59%和112.08%。
人工接锈腐病菌的条件下,用不同放线菌菌株处理的人参产量指标及产量显著高于只接锈腐病菌的对照。根径方面,YBUF5菌株处理组的根径最宽,其次是YBUF7菌株处理组;再次是混合菌株处理;根长方面,YBUF5菌株处理、YBUF7菌株处理、混合菌株处理的根长之间无显著性差异。产量方面,YBUF5和YBUF7菌株单独处理的产量最高,但它们之间无显著性差异,其次为混合菌株处理的人参平均产量,不同处理的平均增产率分别为只接锈腐病菌的235.16%、223.87%和181.95%。人参单根重方面,YBUF5菌株处理的单根重最高,单根增产率为130.59%,其次是YBUF7菌株处理和混合菌株处理的单根重,但它们之间无显著性差异,其单根增产率分别为119.41%和117.58%(表2)。
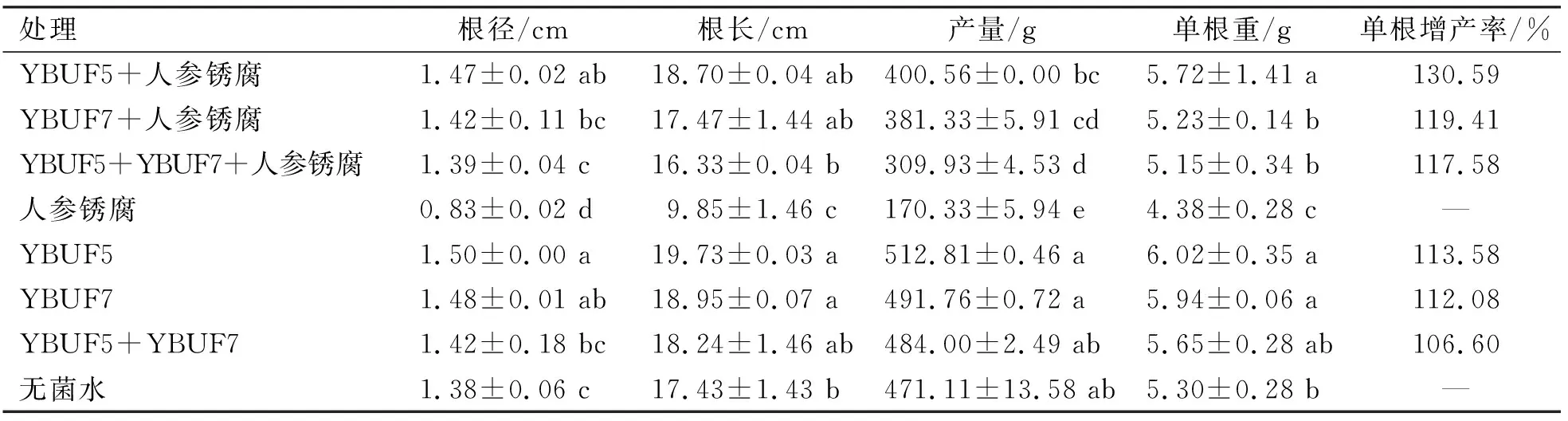
表2 不同处理对人参产量指标的影响
2.2 YBUF5、YBUF7和混合菌株处理对人参叶和根内POD酶活性的影响
由图1,2可见,不同处理的人参叶和根组织内POD活性显著高于无菌对照,并随着时间有降低的趋势。其中,只接锈腐病菌的POD酶活性最高;其次为放线菌+锈腐病菌处理POD酶活性;再次为只用YBUF5、YBUF7处理的POD酶活性。只放线菌处理或放线菌+锈腐病菌处理均为YBUF5处理的POD酶活性较高,其次为YBUF7处理的,混合菌株处理的POD酶活性较低。不同组织中,参叶中的POD活性明显高于参根中的POD活性。
2.3 YBUF5、YBUF7和混合菌株处理对人参叶和根内CAT酶活性的影响
由图3,4可见,不同处理的人参叶和根组织内CAT活性显著高于无菌对照,并随着时间有降低的趋势。其中,只接锈腐病菌的CAT酶活性最高;其次为放线菌+锈腐病菌处理的CAT酶活性;再次为只用放线菌处理的CAT酶活性。人参根中,只放线菌处理或放线菌+锈腐病菌处理中YBUF5、YBUF7以及混合菌株处理间的CAT酶活性差异不显著。人参叶中,只放线菌处理或放线菌+锈腐病菌处理中YBUF5、YBUF7处理的CAT酶活性较高,混合菌株处理的CAT酶活性较低。不同组织中,参叶中的CAT活性和参根中CAT活性间差异不大。
2.4 YBUF5、YBUF7和混合菌株处理对人参叶和根内SOD酶活性的影响
由图5,6可见,不同处理的人参叶和根组织内SOD活性显著高于无菌对照,并随着时间有升高的趋势。其中,只接锈腐病菌的SOD酶活性最高;其次为放线菌+锈腐病菌处理的SOD酶活性;再次为只用放线菌处理的SOD酶活性。人参根和叶中,只放线菌处理或放线菌+锈腐病菌处理中YBUF5、YBUF7以及混合菌株处理间的SOD酶活性差异不大。不同组织中,参叶中的SOD活性明显高于参根中的SOD活性。
2.5 YBUF5、YBUF7和混合菌株处理对人参叶和根内PAL酶活性的影响
由图7,8可见,不同处理的人参叶和根组织内PAL活性显著高于无菌对照,并随着时间有降低的趋势。其中,只接锈腐病菌的PAL酶活性最高;其次为放线菌+锈腐病菌处理的PAL酶活性;再次为只用放线菌处理的PAL酶活性。人参根中,只放线菌处理或放线菌+锈腐病菌处理中接菌50 d时YBUF5处理的PAL酶活性显著高于YBUF7和混合菌株处理的PAL酶活性外,其它时间段不同处理间无显著性差异。人参叶中,接菌30 d时放线菌+锈腐病菌处理间无显著性差异;只接放线菌中YBUF5处理的PAL酶活性高于YBUF7和混合菌株处理的PAL酶活性;接菌40和50 d时,只放线菌处理或放线菌+锈腐病菌处理中YBUF5和YBUF7处理的PAL酶活性高于混合菌剂处理的PAL酶活性。不同组织中,参叶中的PAL活性明显高于参根中的PAL活性。
3 讨论与结论
田间试验中,YBUF5菌株、YBUF7菌株、两者混合菌株以及对照间成活率无显著性差异,其成活率均在90%以上,并对人参植株生长有一定的促进作用。说明YBUF5菌株、YBUF7菌株以及两者混合菌株对人参植株是安全的。
田间防病试验中,YBUF5菌株、YBUF7菌株以及混合菌株对人参锈腐病具有明显的防治效果,其中,YBUF5菌株处理的防效最高,其防效为69.88%;其次为YBUF7处理,其防效为60.24%;混合菌株处理成活率较低,其防效为51.81%。YBUF5菌株的抑菌率高于张阿桃等[15]人选择的放线菌链霉菌属绿色类群1014-5防治人参锈腐病的61.9%。低于马廷会等[16]人选择的链霉菌属中与Streptomycesnaraensis相似的MS32放线菌对人参锈腐病的抑制率76.45%。
人工接锈腐病菌的条件下,用放线菌菌株处理的人参产量指标及产量显著高于只接锈腐病菌的对照。其中,YBUF5和YBUF7菌株单独处理的产量最高,但它们之间无显著性差异,其次为混合菌株处理的人参平均产量。人参单根重方面,YBUF5菌株处理的单根重最高,单根增产率为130.59%;其次是YBUF7菌株处理和混合菌株处理的单根重,但它们之间无显著性差异,其单根增产率分别为119.41%和117.58%。这可能因为YBUF5和YBUF7菌株在一定程度上改善了人参的生长环境,使得人参的产量相对提高。这些结果与金海强等[17]利用解淀粉芽孢杆菌防治人参锈腐病防治试验研究结果基本吻合。
植物体的主要防御酶(POD、CAT、SOD、PAL)活性与植物的抗性有密切的关系,在遇到各种不利的生长因素时,防御酶的活性都会有所上升[18]。POD、CAT、SOD 3者都对植物体内的活性氧清除有重要的作用,SOD能够消除植物体由于新陈代谢而产生的超氧自由基[19],PAL对木质素等次生物质代谢有重要意义[20]。YBUF5和YBUF7菌悬液处理健康人参后人参组织内的主要防御酶活性增加,说明YBUF5和YBUF7菌株可诱导人参组织内主要防御酶活性的增高,使人参植株提前激活自身防御系统来抑制锈腐病菌的侵入。
田间试验中,受人参锈腐病菌胁迫的人参参根及叶片内的主要防御酶(POD、CAT、SOD、PAL)活性都有明显增高,这与宋喜霞等[13]人以及孙嘉曼等[21]人的研究结果一致。接种YBUF5、YBUF7菌株以及混合菌株远高于无菌水处理,说明菌株可以诱导防御酶活性增强,这与郭双双[22],李书强等[23]人的研究结果一致。在面对锈腐病菌侵染的时候,植物体自身会产生防御酶来抵抗病菌,而随着时间的推移,病害加深,植物体自身的防御无法防御时,防御酶的活性就开始降低,这与刘延硕等[24]人的研究结果一致。由于拮抗放线菌的加入,防御酶的活性相对只接病原菌的较低,这也说明了这2种拮抗放线菌能抑制人参锈腐病菌的增殖,这与周东兴[25]对番茄枯萎病防治所得出的结论相同。
单株处理的防御酶活性普遍高于混合菌株处理的防御酶活性。时间上,POD、CAT、PAL酶活性在第30天时最高,而SOD酶活性却在第50天时最高。原因可能是拮抗放线菌对前3种防御酶活性诱导较快,对SOD酶活性诱导较慢。说明这几种酶的活性变化不是同步的,这与王勇[26],车喜庆[27]等人的研究结果一致。生防微生物的生防机制不一定是单一的,往往是2种或更多生防机制协同发挥作用[28]。该试验中YBUF5、YBUF7菌株的生防机制等有待进一步研究。
猜你喜欢
现代园艺(2022年19期)2022-09-21
实用手外科杂志(2022年2期)2022-08-31
中外文摘(2022年12期)2022-08-02
护理与康复(2021年4期)2021-12-07
小读者(2021年19期)2021-11-07
健康之家(2021年19期)2021-05-23
智慧健康(2020年5期)2020-03-24
建材发展导向(2019年10期)2019-08-24
作文评点报·中考版(2019年4期)2019-02-18
课外语文·中(2018年10期)2018-11-16
