
不同氮效率油菜种质苗期氮吸收转运与利用差异研究
2021-09-26 08:34璐王建强2韩配配李银水顾炽明胡小加谢立华
作物杂志 2021年3期
秦 璐王建强,2韩配配,3 李银水 顾炽明 胡小加 谢立华 廖 星
(1中国农业科学院油料作物研究所/农业农村部油料作物生物学与遗传育种重点开放实验室,430062,湖北武汉;2青岛清原化合物有限公司,266000,山东青岛;3江苏沿海地区农业科学研究所,224002,江苏盐城)
油菜是我国重要的油料作物之一,菜籽油占国产油料作物产油量的57%以上,是第一大食用植物油[1]。油菜的高产依赖于氮肥的大量施用,但随着施氮量的增加油菜产量并不会持续同步增长。目前,我国油菜生产的氮肥施用量高达200~330kg/hm2,是一般禾本科作物的3倍[2],但其表观利用率仅为34.0%[3]。这表示每生产100kg油菜籽,需要吸收纯氮9~12kg[4],氮肥利用率低已成为限制我国油菜产业发展的一个重要因素。在油菜化肥减施增效的大环境下,通过生物学手段挖掘油菜种质资源氮高效潜力是提升氮效率的有效方法之一。前人已经对多种作物氮高效种质进行评价与筛选[5-8],开展了关于玉米[9]、小麦[10]和水稻[11]氮高效机理的研究,为油菜氮高效机理研究提供了很好的借鉴。但是氮高效机理涉及的途径复杂多样,前人研究结果也有差异。钟思荣等[12]研究了不同基因型烟草氮吸收效率与利用效率的差异,结果表明氮吸收效率对氮效率的直接作用均大于氮利用效率,氮高效型烟草氮吸收效率显著高于氮低效型烟草,而氮利用效率无显著差异。陈颖等[13]从氮高效吸收和利用角度对氮高效机理进行了阐述,着重强调了根系对水稻氮高效吸收的重要意义。熊淑萍等[14]对不同氮效率小麦品种苗期氮吸收同化能力差异的研究表明,氮高效型小麦品种对NO3-和NH4+的吸收和同化能力均显著高于氮低效型小麦品种。田飞[15]从氮代谢的角度研究了氮效率差异与油菜种质氮同化差异,发现氮高效油菜种质叶片具有较强的硝酸盐同化能力。
氮高效是作物自身遗传特性与复杂环境共同作用的结果。Moll等[16]将氮效率分为吸收效率和利用效率,氮吸收、转运、同化、利用与再利用等因素均会影响氮效率的最终表现。目前,对不同氮效率油菜氮吸收、转运和利用等还缺乏系统的研究。本研究以不同氮效率油菜为试验材料,在不同氮浓度处理下,系统地研究不同氮效率油菜种质苗期氮吸收、转运和利用效率的差异,以期为油菜氮高效机理的深入研究提供理论依据。
1 材料与方法
1.1 材料与设计
1.1.1 供试材料 材料为2个低氮胁迫处理下氮效率差异的甘蓝型油菜种质:氮高效种质6号(H6)和氮低效种质18号(L18),以油菜植株生物量、养分累积量和籽粒产量为筛选标准,2个种质是经过2次水培和2次大田试验从350份甘蓝型油菜种质资源中筛选获得[17]。前期研究结果表明,在低氮胁迫处理下2个油菜种质氮效率存在明显差异,主要体现在氮利用效率。
1.1.2 试验设计 以改良后的1/2浓度Hoagland营养液配方为基础[18],通过水培营养液设置正常氮(CK)和低氮(LN)2个氮浓度处理。CK的氮浓度为9500μmol/L(pH 5.8~6.0),LN氮浓度为CK浓度的1/50,即为190μmol/L。其他养分浓度相同,即LN处理时去掉的KNO3和Ca(NO3)2里的钾和钙,分别用相应浓度的K2SO4和CaCl2补充,具体配方见表1。

表1 供试营养液配方Table 1 The formula of nutrient solution
水培容器为32cm×24cm×12cm规格的蓝色聚乙烯塑料面包箱。试验在植物生长间进行,光照时长为12h/12h(白天/黑夜),温度24℃。油菜种子消毒后,用改良的1/4浓度Hoagland营养液催芽,每个处理4次重复。
具体操作如下:催芽前将育苗杯和水培用的周转箱和定植板用0.037%稀盐酸浸泡,防止青苔生长,浸泡24h后用清水清洗多遍,防止盐酸残留。选取饱满一致的油菜种子,用蒸馏水浸泡30min后,用5%的NaClO溶液消毒20min,并且将种子用纯水冲洗多次后,均匀播于盛有1/4浓度完全营养液的育苗杯上育苗,遮光催芽2d后,见光生长5~7d。待幼苗适宜移栽时,选取长势一致的幼苗移栽到盛有不同氮浓度营养液的周转箱上进行培养。每周更换1次营养液,换营养液时适当分根,防止根系缠绕在一起,同时保证不能伤根。处理14d后,植株出现明显缺氮症状时,选取生长较为一致的幼苗,观察植株表型,测定植株生物量以及氮相关指标。
1.2 测定项目与方法
1.2.1 根系表面积 利用扫描仪(EPSON J221A)获取油菜根系二维图片,用WinRHIZO根系分析软件(Pro 2012b)计算根系表面积。
1.2.2 氮含量 从不同处理中各选取4株生长较为一致的植株,将根系和地上部分别装于牛皮纸袋并编号;在105℃下杀青35min后在75℃下烘干至恒重,并称重。烘干的植株样品研磨后,称取0.1500g置于消化管中,加入少量蒸馏水润湿样品,加入5mL浓硫酸摇匀后放置过夜。次日加入3mL(30%)双氧水,180℃消煮30min左右;待温度达270℃时,重复上步操作;待温度达到360℃时,再次加入3mL(30%)双氧水,继续消煮直至消化管中液体变透明;冷却后,过滤定容至50mL,参照李朝英等[19]的方法用凯氏定氮仪(KDY-9820)测定氮含量。
依据Moll等[16]、徐子先[17]和周健民等[20]的方法测定油菜苗期氮营养效率指标,按以下公式计算:
氮累积量=地上部氮含量×地上部干重+根系氮含量×根系干重;氮生理利用效率=生物量/氮累积量;氮效率系数=LN生物量/CK生物量;转运系数=地上部氮含量/根系氮含量;根系干物质利用效率(单位干物质形成的根系表面积)=根系表面积/根系干重;根系氮利用效率(单位氮形成的根系表面积)=根系表面积/根系氮累积量。
1.3 数据处理和统计分析
采用Microsoft Office Excel 2016对数据进行整理;用IBM SPSS Statistics 20.0(LSD法)进行数据分析。
2 结果与分析
2.1 不同氮浓度处理下氮效率差异油菜种质的生物量比较
通过二因素方差分析(LSD法)对不同氮浓度处理下氮效率差异油菜种质生物量进行分析,结果(表2)表明,不同氮浓度处理对不同氮效率油菜种质苗期生物量的影响大于基因型的影响。氮高效种质H6和氮低效种质L18在CK处理培养14d时,植株干物质累积量无显著差异,但是在LN处理下H6生物量显著大于L18(P<0.05),为L18的2.07倍;其中H6地上部干重和根干重分别为L18的2.28和1.73倍。此外,H6氮效率系数(LN/CK)为0.42,而L18仅为0.21,表明H6在LN处理下的整体生长优于L18,对低氮胁迫的耐受性强,表现出低氮胁迫下氮高效的特征。另一方面,二者在CK处理下根冠比无显著差异,而在LN处理下根冠比均显著增加,这表明LN处理下植株把更多的干物质分配至根系,供其生长所需;然而LN处理下L18根冠比增加显著大于H6(P<0.05),为H6的1.27倍,而地上部干重仅为H6的43.75%,这表明LN处理下,氮低效种质分配到根系的干物质增加,相对增加了根系的生长,导致分配到地上部的干物质减少;相比之下,氮高效种质地上部和根系的干物质分配更为合理,不仅促进了根系生长,而且相对维持了地上部的生长。

表2 不同氮浓度处理下氮效率差异油菜种质生物量及根冠比比较Table 2 Comparison of biomass and root-shoot ratio in rapeseed germplasms with contrasting N efficiency at different N levels
2.2 不同氮浓度处理下氮效率差异油菜种质氮吸收累积量比较
氮效率包括氮的吸收效率和利用效率,众多研究表明,氮吸收效率在氮高效中扮演着重要的角色。基于培养介质供氮量一致,氮吸收效率表征植物对氮吸收和累积的能力大小,因此,用植株氮累积量来衡量不同油菜种质的氮吸收效率。结果(图1)表明,不同氮浓度处理对不同氮效率油菜种质苗期氮累积量的影响大于基因型。在CK处理下,培养14d时,H6植株整株氮累积量比L18高8.13%,且LN处理加大了二者氮累积量的差异。与L18相比,H6植株总氮累积量显著增加42.11%。由此可见,氮高效油菜种质在低氮胁迫下对氮的吸收累积能力表现更加突出。从根系氮累积量(图1)可以看出,在CK处理下,H6根系的氮累积量显著高于L18,但是地上部无显著差异;而在LN处理下,油菜种质之间根系氮累积量无显著差异。同时,LN处理下H6地上部氮累积量为L18的1.88倍,因此,H6分配或转运到地上部的氮相比L18增加更为明显。综上所述,H6在正常氮处理和低氮胁迫下均表现出相对较高的氮吸收累积能力,尤其是LN处理下,显著高于L18,同时结果显示,氮充足时,H6根系吸收能力较高,而氮缺乏时,H6地上部积累的氮更多,说明除了吸收能力外,氮高效种质对氮的转运效率可能相对更高。

图1 不同氮浓度处理下氮效率差异油菜种质氮吸收累积量比较Fig.1 Comparison of N accumulation in rapeseed germplasms with contrasting N efficiency at different N levels
2.3 不同氮浓度处理下氮效率差异油菜种质氮转运系数比较
植物吸收的氮主要由叶片同化,进而被植株利用,所以氮的高效转运对于氮效率提升具有重要意义。从表3可以看出,在CK处理下培养14d,H6植株氮含量高于L18(P<0.05),整株氮含量相比L18增加8.38%。而LN处理14d时,H6植株氮含量显著小于L18,整株氮含量为L18的68.74%,其地上部和根系氮含量均显著小于L18,分别是L18的84.57%和62.08%,但是,H6的氮转运系数为L18的1.37倍,说明H6虽然单位干物质氮含量低,但是氮从根系向地上部的转运能力更强。
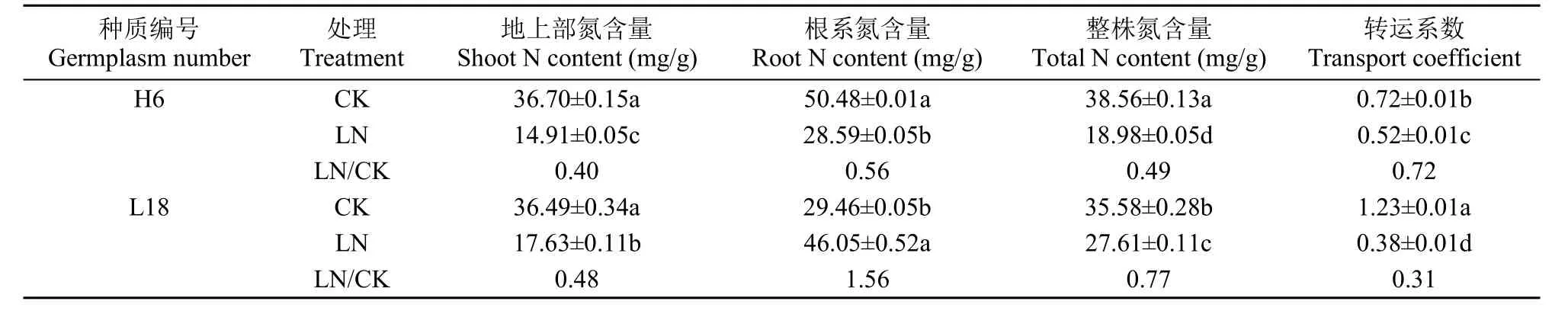
表3 不同氮浓度处理下氮效率差异油菜种质氮含量和转运系数比较Table 3 Comparison of N content and transport coefficient in rapeseed germplasms with contrasting N efficiency at different N levels
2.4 不同氮浓度处理下氮效率差异油菜种质氮生理利用效率比较
氮生理利用效率表示植物单位吸氮量所产生的干物质或产量,可以直接反映植株对所吸收氮的利用能力。从图2看出,在CK条件下,不同氮效率油菜种质氮生理利用效率无显著差异;在LN处理下,不同氮效率油菜种质氮生理利用率均显著提高,平均提高61.1%;相比而言,H6提高的幅度更为突出,提高了2.02倍;但是L18根系氮生理利用率显著降低,单位氮形成的根系干物质量相对较低,仅为CK的63.98%。进一步分析2个油菜种质地上部和根系氮生理利用率发现,LN处理下H6地上部和根系的氮生理利用效率均显著大于CK处理,分别为CK处理的2.46倍和1.76倍;并且H6地上部和根系的氮生理利用效率也显著大于L18,分别是其1.19倍和1.61倍。综上所述,与CK处理相比,LN处理氮效率差异种质H6和L18地上部氮生理利用率均显著提高,且H6根系氮生理利用率也显著提高,而L18根系氮生理利用率显著降低。因此,氮高效油菜种质根系氮的高效利用可能是其氮高效的原因之一。
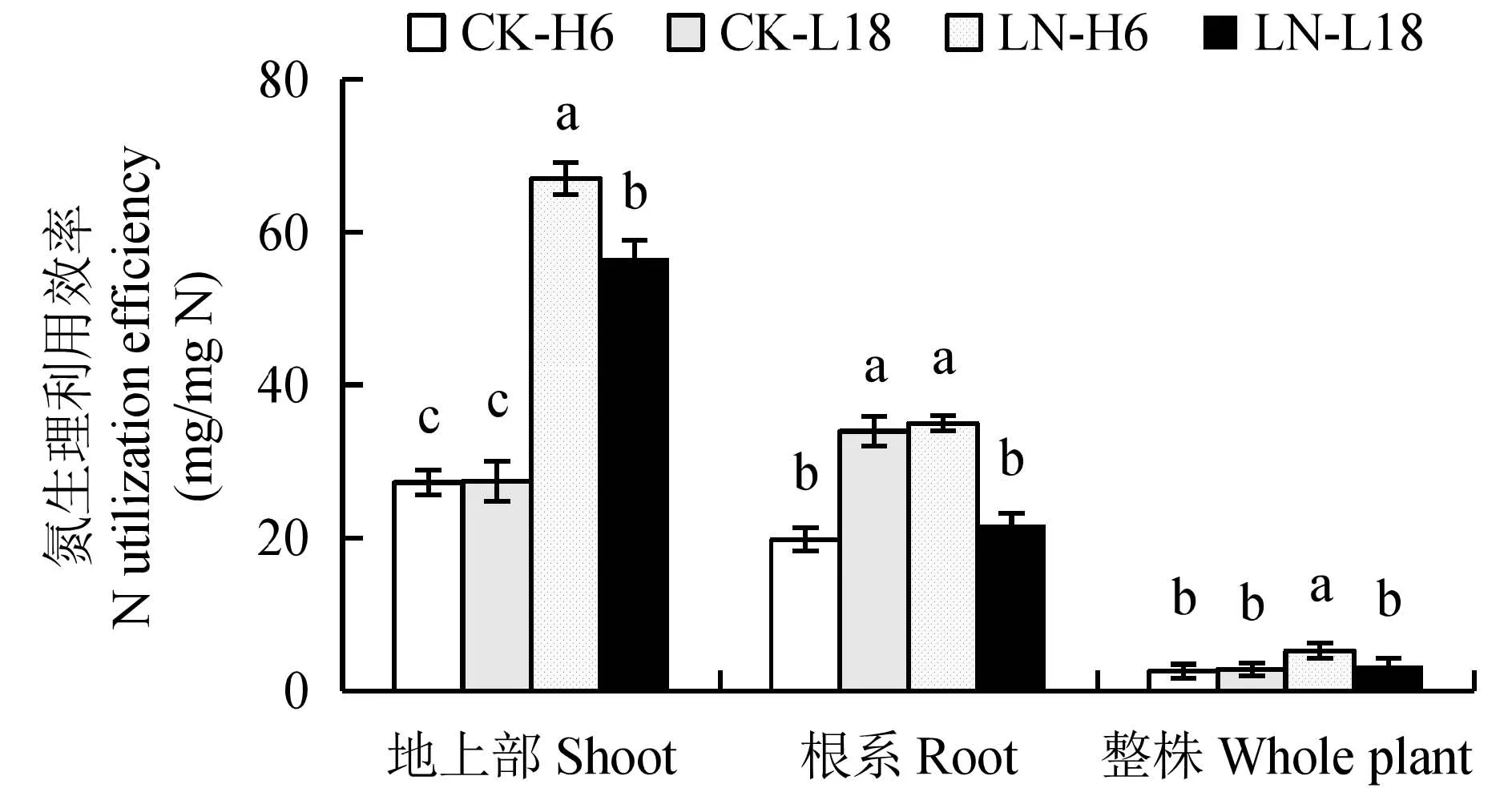
图2 不同氮浓度处理下氮效率差异油菜种质氮生理利用效率比较Fig.2 Comparison of nitrogen utilization efficiency in rapeseed germplasms with contrasting N efficiency at different N levels
对油菜种质根系干物质利用率和氮利用率分析(表4)可以看出,CK处理下,氮效率差异种质H6和L18根系干物质利用率无显著差异,而LN处理下H6根系干物质利用率显著大于L18(P<0.05),为L18的1.22倍;CK处理下,H6和L18根系氮利用率存在显著差异(P<0.05),这可能与种质自身遗传特性有关。在LN处理下,H6根系氮利用率显著大于CK处理,为CK处理的2.24倍,而L18根系氮利用率仅为CK处理的63.57%。综上所述,H6 LN处理下单位干物质和单位氮可形成较大的根系吸收面积,使根系具有较高的养分吸收能力。因此,LN处理下氮高效种质一方面可通过提高根系碳氮利用能力,形成发达的根系;另一方面以有限的碳氮养分形成较大的根系吸收表面积来提升根系养分吸收能力,这可能是油菜实现氮高效的关键原因之一。

表4 不同氮浓度处理下氮效率差异油菜种质根系干物质利用效率和氮利用效率比较Table 4 Comparison of dry weight utilization and N utilization in roots of rapeseed germplasms with contrasting N efficiency at different N levels
3 讨论
植物氮效率是一个非常复杂的性状,涉及植物对氮的吸收、同化、运输、利用和再利用等方面,是自身遗传和环境因子共同作用的结果[21]。陈二影等[22]研究了不同基因型谷子氮吸收利用基因型差异,发现不同生态类型谷子品种的氮吸收和利用效率差异显著,华北夏谷类型品种之间的氮利用效率差异最大。匡艺等[23]研究结果表明,在低氮和正常供氮条件下,小黑麦的氮利用效率都有较大的基因型差异。不同氮效率小麦品种的研究进一步表明氮高效型小麦品种根系对NO3-和NH4+的吸收和同化能力均显著高于氮低效型小麦品种[14]。本研究发现在正常氮浓度培养条件下,氮效率差异油菜根系氮吸收累积量和根系氮生理利用率同样存在一定差异,这可能与种质自身遗传特性有关,前人研究[16]也表明,在氮充足的情况下,植物氮效率的差异主要表现在吸收效率,而根系对氮的吸收有着重要作用,而氮缺乏时,氮效率的差异主要在利用效率。本研究同样表明,低氮胁迫加大了二者氮效率的差异,尤其是对油菜氮的转运与利用产生了更为显著的影响。
根系是植物吸收养分和水分的主要器官,也是最先感知根际环境条件变化的器官,为了适应外界环境变化根系会表现出极强的可塑性[24-25]。本研究发现,氮效率差异种质根系生物量表现出不同的变化趋势,氮高效种质根系生物量在正常氮和低氮处理间无显著差异,而氮低效种质根系生物量在低氮胁迫处理下显著降低。熊淑萍等[14]对小麦根系吸收特性与氮利用差异的分析研究表明,较高的根系生物量和根系总吸收面积能促进根系对氮的吸收,是氮高效的基础,合理的根冠比能促进作物对氮的高效利用,是氮高效的关键,这与本研究结果类似。王艳等[26]研究表明,玉米苗期根系形态直接与氮吸收效率相关,根系形态对氮的高效吸收具有重要作用,尤其是氮胁迫条件下。崔文芳等[27]对杂交玉米氮高效的研究中进行了氮吸收效率和利用效率对氮效率贡献的通径分析,结果表明氮吸收效率对氮效率的贡献大于氮利用效率,更加突出了氮吸收效率的重要性。在本研究中氮效率差异油菜种质在低氮胁迫处理下根冠比均显著增加,同样说明油菜低氮胁迫下优先促进根系生长,增强养分吸收能力,这与前人的研究结果较为一致。
此外,从本研究结果可以看出,与正常氮浓度相比,不同氮效率油菜在低氮胁迫处理下植株氮含量均不同程度地降低,但是氮高效种质氮转运系数显著增加,而氮低效种质显著减小,这表明低氮胁迫下,氮高效种质氮由根系转运到地上部的能力提高,而氮低效种质氮转运能力相对较差。韩永亮[28]对不同氮效率油菜硝态氮长距离运输和短途分配差异研究表明,正常供氮水平下,氮高效型油菜有更多的硝态氮被运输到植物的地上部分参与同化。同时Hajari等[29]对甘蔗氮高效研究同样发现,氮高效型甘蔗中的硝态氮从根部到地上部的转移率较高。因此,高效的氮素转运能力可能是低氮胁迫下油菜氮高效的一个重要原因。
本研究结果显示,与CK相比,LN处理下氮高效油菜种质根系干物质利用率、根系氮利用率和氮生理利用率均显著提高,而氮低效油菜种质根系氮生理利用率显著降低,说明氮高效油菜种质对吸收的氮具有高效的同化、分配和利用能力。前人研究[30-31]也发现氮高效作物叶片谷氨酰胺合成酶具有较高的活性,有利于氨基酸和蛋白质的合成与转化。Hirel等[32]研究表明,超表达谷氨酰胺合成酶基因(GS)可以提高小麦的籽粒产量。这也说明氮高效作物在氮的利用方面具有一定优势。因此,低氮胁迫下高效的氮同化和利用对于油菜氮高效同样起着较为重要的作用。目前,对油菜氮高效机理研究而言,氮吸收效率和氮利用效率对氮效率的决定作用尚无统一的结论,需要进行更加细致深入的研究。
4 结论
油菜氮高效是自身遗传潜力与环境因素共同作用的结果,在正常氮浓度条件下供试油菜种质苗期氮效率存在一定的差异,表现为氮高效油菜种质根系氮吸收累积量大于氮低效种质,而低氮胁迫加大了不同氮效率油菜种质在氮吸收累积、转运及利用方面的差异。二因素方差分析(LSD法)表明,不同氮浓度处理对苗期油菜氮效率的影响大于基因型本身,低氮胁迫下除去种质自身遗传因素的影响,氮效率差异种质氮吸收能力均降低,而氮转运和利用能力提高。但是,低氮胁迫下氮高效油菜种质的氮吸收累积量、转运系数和利用效率均显著大于氮低效种质。综上所述,低氮胁迫下,氮高效种质在苗期可通过提升氮的吸收、转运和利用效率综合来实现油菜氮高效。
猜你喜欢
今日农业(2022年13期)2022-09-15
今日农业(2021年21期)2021-11-26
今日农业(2021年14期)2021-10-14
今日农业(2019年14期)2019-01-04
中国麻业科学(2018年6期)2018-04-09
娃娃乐园·综合智能(2018年3期)2018-03-22
数学小灵通(1-2年级)(2017年10期)2017-11-08
水利规划与设计(2016年9期)2017-01-15
西南农业学报(2016年5期)2016-05-17
广西林业科学(2016年3期)2016-03-16
