
外源物质干预对逆境胁迫下植物生长代谢的影响研究进展
2021-09-26 02:10任延靖青海大学农林科学院青海省蔬菜遗传与生理重点实验室青海西宁810016青海大学三江源生态和高原农牧业国家重点实验室青海西宁810016青海省农业技术推广总站青海西宁810000
青海农技推广 2021年3期
任延靖 柳 红(1.青海大学农林科学院青海省蔬菜遗传与生理重点实验室,青海 西宁 810016) (2.青海大学三江源生态和高原农牧业国家重点实验室,青海 西宁 810016) (.青海省农业技术推广总站,青海 西宁 810000)
逆境胁迫是造成植物减产、品质下降等最主要的因素之一[1]。逆境胁迫包括生物胁迫和非生物胁迫,生物胁迫主要包括病虫危害、杂草危害等;非生物胁迫主要包括寒冷、高温、干旱、盐度、洪水和养分缺乏[2],这两种胁迫均是影响作物产量和野生植物物种分布的限制因素。
目前对植物危害较严重的两种非生物胁迫因子是干旱和盐碱。干旱是一个重大的全球性问题,影响到1%~3%的陆地面积[3],预计到2090年,这一数字将增加到10倍以上[4];土壤盐分是影响植物生长性能和农业生产力的主要问题之一[5],已成为影响3.97×108hm2陆地的世界性问题,高盐灌溉水对约20%的可耕地产生了负面影响,预计到21世纪中叶,这一趋势将上升到50%[6]。
利用外源物质缓解逆境胁迫对植物的危害的研究已成为国内外研究的热门,目前干预植物逆境胁迫的外源物质主要有细胞分裂素、吲哚乙酸等生长调节剂,褪黑素、亚精胺等外源有机物和金属离子、氮磷钾等外源无机物等。近年来,国内外很多学者分别进行了外源物质处理下的作物抗逆研究,但研究较为分散,缺乏系统性的总结。因此本文围绕外源物质干预下对植物遭受非生物胁迫时的生长代谢的影响进行了综述,分析当前研究工作中存在的不足,并提出未来应当加强的方面,以期为后续相关研究提供参考。
1 植物生长调节剂对植物抗逆的影响
农业生产过程中,农作物不可避免地遭受低温、干旱、盐碱、涝灾及病虫害等不良因素地影响,在这些逆境条件下,农作物生长受到抑制,幼苗素质下降,植株抗病能力、营养生长和生殖生长均受到严重损害,最终影响农作物的产量。而植物生长调节剂在这种情况下可以有效地提高作物抗逆能力,减轻这种负面影响,从而促进作物的生长发育。
1.1 细胞分裂素影响逆境胁迫下的植物内含物变化
细胞分裂素(CTK)是一类能够调节植物生长和发育进程的物质[7]。最近地研究发现CTK能够提高植物体内的可溶性糖含量,通过可溶性糖的渗透作用提升植物对渗透胁迫的适应性,进而提高植物的生长和产量。还有研究表明CTK能够直接或间接地清除自由基,减少脂质过氧化作用,提高过氧化氢酶(CAT)等膜保护酶的活性,改变膜脂过氧化产物、膜脂肪酸组成的比例,保护细胞膜,促进植物在遭受逆境胁迫后的生长。分子生物学研究表明植物中较高含量的CTK可使非生物胁迫下激活的异常转录程序回归正常,从而促进植物对渗透胁迫的适应,进而促进植物的良好生长和产量的提高。Prerostova等[8]通过CKX和IPT两个基因的过表达转化拟南芥试验,也证实了CTK在干旱驯化中具有重要作用。因此有针对性地控制植物中CTK地含量是培育耐旱植物的有效措施。
1.2 吲哚乙酸提高逆境胁迫下植物对重金属的耐受性
吲哚乙酸(IAA)是一类不饱和芳香环的(UAR)小分子有机化合物,不仅能够参与调节植物的生长发育,还可以提高植物对重金属的耐受性,抑制植物对重金属的吸收。在向日葵的研究中发现施用外源IAA能够促进植物生长、减少铅和锌的毒性作用,还能够增强向日葵的抗逆性;在对香草的研究中发现施用外源IAA可以降低香草体内丙二醛(MDA)含量,有效减轻镉胁迫对香草生长的影响;对砷胁迫的凤尾蕨施用20mol/L IAA,发现凤尾蕨不仅能够正常生长,还能够提高植物内砷的富集;Zhou等[9]研究了IAA对香樟镉毒性的影响,结果表明外源IAA可提高香樟的净光合速率、增加渗透调节物质浓度、去除活性氧自由基,改善香樟的光合性能和生长环境,从而降低镉对香樟的毒性作用。
1.3 脱落酸通过调节植物体内酶活性提高植物的耐受性
脱落酸(ABA)是一种可以介导植物脱水应激反应信号级联的植物激素[10],在逆境胁迫下植物会启动脱落酸的合成系统,合成大量脱落酸,进而加快植物气孔地关闭,促进水分吸收,减少水分运输的途径,降低叶片伸展率;并且可以诱导植物抗旱特异性蛋白质合成,调整保卫细胞离子通道,诱导ABA反应基因改变相关基因的表达,从而增强植物抵御逆境胁迫的能力。Salvi等[11]研究了鹰嘴豆幼苗在脱水胁迫和ABA处理后的变化,结果表明肌醇半乳糖苷合酶在提高植物脱水胁迫耐受性方面发挥着重要作用,且肌醇半乳糖苷合酶通过保护膜损伤免受ROS攻击来介导脱水应激耐受性。植物体内的脱落酸编码清除活性氧以及渗透液的生物合成也是由ABA诱导的,它协助确保了大分子的正常功能,并保持了细胞的完整性,以避免逆境胁迫对植物存活的不利影响。
1.4 乙烯通过调节植物体内的酶活性调控植物对逆境胁迫的耐受性
乙烯(C2H4)被认为是一种应激激素,参与了许多非生物应激反应。乙烯的生物合成途径包括通过ACC合成酶将S-腺苷甲硫氨酸转化为氨基环丙烷羧酸(ACC),然后通过ACC氧化酶(ACO)将ACC转化为乙烯,生成ACC通常是乙烯产生的限速步骤。一般来说,种子萌发受到激素的严格控制。研究发现,乙烯可以减轻中度盐胁迫对种子发芽的抑制。Lin等[12]分析了拟南芥种子在不同盐浓度条件下萌发过程中乙烯与一氧化氮(NO)的关系,结果表明外源1-氨基环丙烷-1-羧酸盐(ACC,乙烯生物合成的前体)或硝普钠(SNP,NO供体)的应用在很大程度上克服了盐对种子发芽的抑制作用。Zhu等[13]研究发现在外源施用乙烯可以减轻采后冷害过程对桃细胞壁和脂质代谢的影响。作为气态分子,正常情况下植物组织内的乙烯维持在一个极低的水平,胁迫刺激后产生的乙烯可以很快地向邻近组织扩散,使得乙烯可以快速、灵敏地应答逆境胁迫,将信号传递到效应部位引起一系列应激反应;盐、冷胁迫下乙烯可通过调节植物体内的CAT等各种酶活性进而提高植物的耐受性。
1.5 水杨酸通过提高植物抗氧化性提高植物耐受性
水杨酸(SA)是一种植物内源信号物质和新的植物激素,能够提高植物抗氧化系统的效率和对氧化胁迫的耐受性。对不同植物模型的大量研究表明,SA作为信号分子在局部抗性反应和防御相关基因表达中起着重要作用。在非生物胁迫下,SA可诱导植物中脯氨酸的积累,促进抗坏血酸—谷胱甘肽(AsA-GSH)循环,提高过氧化氢酶(CAT)、过氧化物酶(POD)、超氧化物歧化酶(SOD)、抗坏血酸(AsA)和金属解毒系统等抗氧化剂的功效[14]。Han等[15]研究了外源水杨酸对安娜鸢尾遭受镉毒性的缓解作用,结果表明施用1μmol/L SA预处理六角鸢尾幼苗的根,可提高其干重、光合速率、抗氧化酶活性,保护植物免受镉引起的氧化应激损伤。在植物遭受逆境胁迫时加入外源SA能促进不溶性糖和蛋白的水解,从而增加细胞可溶性糖SS)、可溶性蛋白(SP)及游离氨基酸(PRO)含量,继而发挥上述物质对植物细胞的渗透调节作用,通过这种方式,维持气孔开放和光合作用,进而保持植物在逆境胁迫中继续生长。
2 外源有机物对植物抗逆的影响
2.1 褪黑素可调控多种逆境因子胁迫下植物的耐受性
褪黑素(MT)是一种兼具亲水性和亲脂性的新型生物调节因子[16],在植物发育和抗逆性中起着重要作用。外源褪黑素可以降低植物细胞膜脂过氧化,缓解低温对植物细胞的损伤。植物具备在线粒体和叶绿体中合成MT的能力[17]。据报道,在胁迫条件下,MT能促进植物生长、延缓衰老、调节光周期反应和根系结构[18]。MT还可以保护植物免受诸如热[19]、冷[20]、盐[21]、干旱和重金属等非生物胁迫,通过植物体内酶促与非酶促系统对活性氧的清除从而增强了植物的抗逆性[22]。相关报道表明,分别采用50μmoL/L外源MT应用于菊苣和100μmol/L外源MT处理水生植物牛膝菊,均能够通过改善植物生长、光合作用和抗氧化系统来减轻镉的毒性[23]。Namdjoyan等[24]研究探讨了褪黑素作为信号分子在改善铅(Pb)胁迫下的红花幼苗氧化损伤中的可能作用,结果表明MT可以通过降低植物对Pb的吸收和根系到芽的迁移来减轻Pb对植物的毒性,同时调节不同的抗氧化系统。结果还表明,尽管褪黑素对提高Pb植物修复潜力的作用不大,但信号分子的应用可通过刺激抗氧化防御机制来提高红花在Pb污染土壤中的存活能力。
相关报道还表明,外源MT可减轻盐胁迫下小麦和玉米[25]的水分损失。在干旱条件下的番茄叶片中,MT通过促进角质蜡层厚度的增加来恢复叶面水分损失[26],因此,MT可以通过提高叶面水势、减少水分损失和维持植物新陈代谢来保护植物免受水分胁迫。外源MT通过减少番茄[27]和水稻[28]在低温胁迫下的活性氧和丙二醛的积累以及电解质渗漏来维持氧化平衡。早期的研究表明,外源MT激活了抗氧化酶,促进了非酶抗氧化剂的积累,从而抵消了环境应激所造成的损害[29]。Li等[30]报道了褪黑素在最佳剂量下的应用,无论是在香茅叶片上还是在根上,不仅在应用部位诱导了冷应激耐受性,而且在未经处理的远处也系统地诱导了冷耐受性。褪黑素的研究对全球生态环境的改善有促进作用,褪黑素的研究是具有可行性、经济性、重要性、应用性的,必将会极大地降低投入在环境保护方面的成本。
2.2 亚精胺是与植物抗逆性关系最密切的一类次生代谢产物
亚精胺(SPD)属于多胺的一种,是生物生命活动过程中产生的一类次生代谢物,在植物中具有调节基础代谢、影响生长发育、控制形态建成、延缓衰老等功能,且亚精胺与植物抗逆性的关系最为密切。外源SPD可有效缓解植物幼苗在逆境胁迫下自由基对细胞膜的损伤程度,主要表现为:SOD、POD和抗坏血酸过氧化物酶(APX)的活性增强,抗坏血酸(AsA)和类胡萝卜素含量的增多,植株的抗氧化能力相对提高;在逆境胁迫下外源SPD处理的植物叶片中脯氨酸含量逐渐升高,促进了维持氮素代谢和细胞质酸碱平衡从而达到提高植物面临逆境下的耐受性。利用亚精胺作为外源物质调节植物抗逆性的研究已有很多报道,如张毅[31]研究了亚精胺缓解盐碱胁迫对番茄幼苗的伤害的效应及其调控机理,结果表明外源SPD参与调节了盐碱胁迫下番茄幼苗体内多胺代谢,从而提高了植株的耐盐碱性;Banerjee等[32]研究了SPD对长期氟胁迫下易感籼稻品种IR-64的改善作用,结果表明通过限制阴离子通过氯化物通道,SPD处理大大减少了籼稻体内氟化物的生物累积。随着SPD生物活性的不断被发掘,其在植物抗逆生长过程中将不断发挥积极作用。
2.3 γ-氨基丁酸影响植物生长发育且可提高盐胁迫下的植物耐受性
γ-氨基丁酸(GABA)是一种以自由态形式广泛存在于原核生物和真核生物的四碳非蛋白质氨基酸,能够影响植物的生长发育[33],在植物中,GABA通过三羧酸(TCA)循环中的两个酶促反应过程合成和代谢。具体而言,是先由α-酮戊二酸转化为谷氨酸,然后通过谷氨酸脱羧酶(GAD)脱羧形成GABA,随后,GABA转氨酶和琥珀半醛脱氢酶分别转化为琥珀半醛和琥珀酸。GABA是三羧酸循环中琥珀酸的重要来源,GABA有利于植物体内的碳氮平衡,调节细胞质pH,抵御氧化胁迫。在植物中,GABA不仅可以作为代谢物,而且可以作为信号分子。最近发现了一种GABA受体,即铝激活的苹果酸转运体(ALMT),提供了GABA作为信号分子的证据[34]。最近的遗传和生理研究结果表明,GABA不仅是对诸如盐胁迫等非生物胁迫的反应所必需的[35]。
2.4 脯氨酸通过调节植物抗氧化酶提升植物耐逆性
脯氨酸(PRO)是一种氨基酸,它揭示了胁迫下植物的有益特性,远远超出了其作为相容溶质的功能[36]。由于脯氨酸其独特的螯合特性、参与细胞信号机制以及积极参与植物抗氧化系统的非酶成分,其在提高植物对各种非生物胁迫的耐受性方面的潜力已逐渐得到承认。
2.5 l-肉碱与植物对非生物胁迫的耐受性有关
L-肉碱(LC)是一种内源性氨化合物,在动物、细菌、一些酵母菌和真菌及植物自然合成的L-赖氨酸和L-蛋氨酸氨基酸中L-肉碱是一种基本的铵化合物,负责所有生物体的能量代谢;除代谢功能外,L-肉碱在细胞中还具有解毒和抗氧化作用、能量代谢、激素作用、适应压力和解毒功能[37]。ONEY-BIROL[38]采用施加外源L-肉碱的方法研究了其缓解盐胁迫下大麦的遗传毒性损伤,结果表明外源1mol LC通过增加有丝分裂和减少氧化应激对大麦幼苗造成DNA损伤来减轻盐胁迫的有害影响。许多类型的植物,如谷物和豆类均含有左旋肉碱,存在于植物的叶子以及干燥和发芽的种子中。此外,L-肉碱的水平与植物对非生物胁迫的耐受性有关。
2.6 海藻糖植物在胁迫下渗透调节的重要渗透保护剂之一
海藻糖(TRE)是一种非还原性二糖,是植物在胁迫下渗透调节的重要渗透保护剂之一。在干燥条件下它不仅是一种能源,而且还是脱水酶、蛋白质和脂膜以及其他生物结构的有效稳定剂。它也是作为涉及应激反应和解毒的基因的激活剂的信号和抗氧化剂分子。但是植物体内的海藻糖的合成不足以防止逆境下的不利影响,因此有报道称外源海藻糖是通过调节抗氧化剂来提高耐盐性[39];另外通过海藻糖的离子调节作用还可提高作物对高盐度的抗性。Rohman等[40]研究了海藻糖减轻玉米在盐度和低磷下的氧化应激中的作用,结果表明海藻糖通过调节抗氧化剂和乙醛酶系统,降低Na+/k+、ROS、MDA和Mg含量,进而促进了玉米幼苗的生长。
2.7 P-香豆酸可影响逆境胁迫下植物的生长代谢
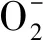
2.8 胺类在植物非生物胁迫中起着重要作用
多胺(PAs)是所有活细胞中普遍存在的天然多态阳离子,对细胞的发育、生长和存活至关重要[44]。多胺不仅有助于植物的生化生理及细胞代谢,如离子通道调节和维持染色质结构和细胞膜,还具有调节酶的功能[45],是调节DNA复制、转录和翻译所必需的物质。特别是在植物中,PAs及其代谢产物起着细胞信号分子的作用,可以增强植物对病原菌和大量的非生物胁迫的耐受性,包括果实的抗衰老作用等[46-47]。
多巴胺(DA)是一种水溶性抗氧化剂,在植物非生物胁迫中起着重要作用。DA不仅可以调节植物细胞的各种代谢活动,如清除活性氧过程,植物糖代谢,调节离子通透性,叶绿体的磷酸化,它还能与植物激素相互作用,影响植物的生长发育[48]。Jiao等[49]研究了外源多巴胺促进苹果幼苗的耐碱性,结果表明外源DA的应用通过提高苹果幼苗绿原酸的水平,进而提高了碱胁迫下苹果幼苗的抗氧化能力。
2.9 果聚糖是调控植物逆境胁迫的渗透调节剂之一
果聚糖是β-D-呋喃果糖的多聚体,是由果糖聚合而生成的多糖的总称,在地球的温带和冷区植物中主要积累果糖聚合物,称为果聚糖。12%到15%的被子植物含有果聚糖。它们产生于单子叶植物和双子叶植物中,分布于茎、叶、花序和种子中。果糖聚合度(DP)也与抗氧化能力有关。例如,在遭受干旱的小麦幼苗中,高DP果聚糖的积累确保了快速消除抗氧化损伤[50]。低温但不冻结的条件下,在植物生长发育时期内,果糖的积累往往与耐冻性的增强有关。果聚糖还是渗透调节剂,受干旱环境的调节,干旱对果糖代谢的影响通常包括水解活性的提高、果糖DP的降低和渗透调节能力的提高[51]。因此在今后研究富含果聚糖的植物抗逆方面,可重点关注果聚糖变化规律。
2.10 谷胱甘肽作为植物螯合素的前体调控植物抗逆性
谷胱甘肽(GSH)是一种具有多种生理功能的生物活性三肽。据报道,细胞中GSH水平影响氧化还原状态和氧化还原敏感信号通路,如光合活性。此外,糖代谢酶—醛缩酶和三糖—磷酸异构酶是蛋白质s-谷胱甘肽化的靶点,这些酶活性被认为是由GSH控制的,这种控制被认为是为了防止戊糖和己糖代谢物在细胞中过度积累,这些实验结果表明,GSH在糖代谢的调节中起着重要的作用;GSH还参与过氧化物代谢和氧化还原信号传递并且可以在各种酶的参与下调节细胞中的氧化还原状态;GSH还通过谷胱甘肽s-转移酶催化的机制解毒,诸如除草剂等。据报道,GSH是植物螯合素的前体,通过酶聚合GSH来响应重金属元素如根系中的镉而合成,并参与镉元素的长距离迁移。Xiang等[52]研究了增强内源和外源GSH如何影响拟南芥的镉运动和分布的,结果表明在野生型和转基因拟南芥植物中,只有外源GSH能显著抑制镉从根系到芽的迁移。
3 外源无机物对植物抗逆性的影响
3.1 金属离子
3.1.1钙对逆境胁迫下植物的生长发育具有积极作用
对植物来说,钙(Ca)不仅是一种大量的营养元素,还作为一种生理活动的信号物质,还是偶联胞外信号与胞内生理生化反应的第二信使,是植物代谢和发育的主要调控因素之一。钙在植物生长中起着不可或缺的作用,是各种细胞内和细胞外生理活动的信号物质。它具有感知外部环境压力和保持细胞膜的选择性和完整性的功能。当植物受到外界环境的刺激时,植物细胞中Ca2+的浓度会出现特定的变化,并有一系列的保护性生理反应。前期研究表明在植物适应酸中毒、有毒金属、铵中毒、干旱和寒冷过程中,Ca2+可以发挥积极作用,适量的钙可以减轻盐度和干旱等各种胁迫对植物的负面影响,也能有效地增加作物对养分的吸收[53]。Li等[54]研究了外源Ca2+(5mol/L CaCl2)对大豆在不同生长阶段(幼苗、开花期和灌浆期)以及模拟酸雨(pH4.5或pH3.0)胁迫下大豆产量和籽粒品质的影响,结果发现Ca2+的应用可以调节质膜H+-ATP酶的活性,以减轻其作用。Xu等[55]研究了钙对植物镉毒性浓度的影响,结果表明钙在镉胁迫下的植物生长、重金属积累和迁移、光合作用、氧化损伤和信号转导中起着重要作用。
3.1.2磷缺乏可阻碍植物光合作用系统的形成
植物体中磷(P)是作物植物细胞的重要组成部分,P不足一方面将导致细胞缺失完整性,另一方面可以阻碍植物光合作用系统的形成,另外P的增加可以通过改变和使抗氧化系统失活来促进活性氧的生成[56]或者通过阻碍植物的光合作用系统[57]。Wang等[58]研究了在缺P条件下燕麦根系的转录和代谢反应,结果表明燕麦根在缺P四周后表现出柠檬酸盐的增加以及有苹果酸的渗出,在缺P10d后,有9371个差异表达的转录本有两倍或更大的变化。Jia等[59]探讨了外源P影响多年生黑麦草的Cd防御机制,结果表明,外源性P的施加部分减轻了脂质过氧化影响,降低了超氧化物歧化酶(SOD)、过氧化氢酶(CAT)和过氧化物酶(POD)抗氧化酶的水平,降低了促肾上腺皮质激素清除活性,总的来说,黑麦草植物的Cd解毒可以通过外源P输入来刺激,从而促进螯合介导的Cd解毒过程。
3.2 一氧化氮对植物的耐盐性有促进作用
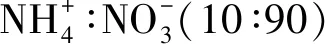
Dai等[65]研究结果表明外源NO可减轻高硒胁迫对水稻幼苗根系形态、生长、光合作用和抗氧化能力的影响,降低MDA、H2O2和脯氨酸含量,且外源NO能有效地降低高硒处理水稻的毒性。Soliman等[66]外源性NO在改善镍(Ni)介导的茄子氧化作用中的作用,揭示了外源性硝普钠(SNP)通过上调抗氧化和乙醛酶系统来提高Ni应激耐受性的效率。Zhang等[67]研究了外源NO和P胁迫对金缕梅幼苗的菌根化、植物生长及相关微生物的影响,结果表明SNP处理和磷胁迫对Tuberindicum的定植率有显著影响,最高水平为仅用100μmol/L SNP和5μmol/L P处理下就可提高T.indicum的定植率,并且对接种后不同时间幼苗的株高、茎围、生物量、根冠比和根活性也有积极影响。He等[68]对NO对大麦根系镉胁迫的耐受性进行了研究,结果表明在Cd胁迫下,两个大麦品种丙二醛含量和电解质水平均有增加。
3.3 其他外源无机物对植物抗逆的影响
硫化氢(H2S)是生物体内继NO和CO后的第3种气体信号分子,在生物生长发育及非生物应答胁迫等方面具有重要作用,已成为生物学研究的热点。已有的研究表明,H2S可以促进种子萌发,根的发生,调节光合作用、气孔运动,并参与抵抗重金属、盐、干旱、高温等胁迫[69]。Kaya等[70]研究了硫化氢部分涉及的硫胺素对草莓植物镉毒性的耐受性,结果表明添加H2S供体硫化钠(0.2mmol/L NaHS)进一步提高了草莓对镉胁迫的耐受性。
4 总结与展望
逆境胁迫因子间是相互关联的,不是单一存在的(图1)。植物在受NaCl影响的土壤上生长后,其对镉的吸收量也会增加;虽然土壤中铁的丰度很高,但其生物有效性非常低,特别是在碱性石灰性土壤上,高pH和高碳酸盐含量严重降低了土壤中铁的有效性。
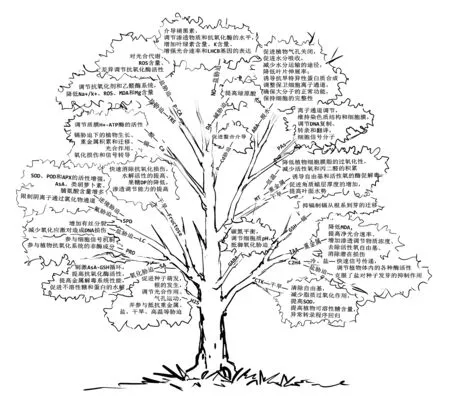
图1 外源物质干预对逆境胁迫下植物生长代谢的影响
在逆境胁迫下植物体内的激素也存在相关调节的机制,如在CK和ABA代谢之间存在一种相互调节的机制,CK和ABA之间的代谢关系目前也已经很清晰,即当CK水平升高时抑制了黄嘌呤脱氢酶活性,黄嘌呤脱氢酶是ABA生物合成的关键酶之一;干旱胁迫下的高浓度CK还可作为乙烯诱导衰老的拮抗剂。研究表明转基因烟草中IPT基因的过表达导致烟草植株中CK浓度的增加进而导致ABA生物合成基因的抑制。在过去的十年中,虽然对CK介导的干旱驯化进行了大量的研究,但其确切的分子机制尚不清楚。随着CRISPR/CAS等新的生物技术手段的出现和全基因组序列的可利用性,未来提高非生物抗逆性的遗传操作将不断提高,未来的研究应集中在发掘新的干旱应答基因和蛋白质,以及细胞分裂素对抗旱性影响的分子机制还应加强与植物抗逆相关基因家族的研究[71]。另外,在外源有机物之间且与植物激素之间同样也存在着相关作用关系,如一定浓度的褪黑素可以提高种子的IAA含量,帮助种子萌发;外源SPD可使逆境胁迫下植物幼苗体内产生超氧阴离子的速率降低,H2O2和MDA的含量下降,提高SOD、POD、CAT、APX、谷胱甘肽还原酶(GR)、AsA和谷胱甘肽(GSH)的活性,抑制ROS的积累和膜脂过氧化,进而提高植株的抗氧化能力和抗逆性。
阐明植物—微生物—环境相互作用的分子基础在植物科学发展中具有重要意义。随着个体应激信号通路的发展,最近的研究开始揭示生物和非生物应激反应之间的分子交叉,以及联合应激反应中的调节原则[2]。与动物不同,植物在面临压力时不能移动。然而,植物已经建立了复杂的信号网络来应对压力,在逆境胁迫作用下引起一系列的反应,以防止或尽量减少损害,同时逆境会诱发植物体内基因表达模式的改变,进而改变相关代谢途径,产生适应胁迫的生理变化[72]。目前对植物胁迫反应的研究已经达到分子水平,但是植物对干旱和其他胁迫的适应取决于抗逆相关基因的表达,也取决于胁迫诱导的各种信号通路的调控,后续可以结合遗传学,深入探索胁迫反应机制,以改善植物在胁迫条件下的生长。
猜你喜欢
舰船科学技术(2022年11期)2022-07-15
中国土壤与肥料(2021年5期)2021-12-02
农业资源与环境学报(2021年5期)2021-10-06
现代农业科技(2016年20期)2016-12-20
现代养生·下半月(2015年6期)2015-09-07
现代养生·下半月(2015年2期)2015-06-15
现代农业科技(2009年19期)2009-03-20
中学生数理化·八年级数学华师大版(2008年3期)2008-08-26
中国新闻周刊(2004年11期)2004-04-07
