
人和猕猴Broca区跨物种解剖连接差异分析
2021-09-21 08:27任凯欣王千山夏潇鸾邓红霞李海芳
太原理工大学学报 2021年5期
任凯欣,王千山,夏潇鸾,邓红霞,李海芳
(1.太原理工大学 信息与计算机学院,山西 晋中 030600;2.澳门大学 认知与脑科学中心,澳门 999078)
Broca区位于人脑的额叶下回,大量的临床和实验研究证实该区主管语言信息的处理和话语的产生,与语义和单词含义的理解,语音和发音方式的形成都有密切的联系[1]。除此之外,该区对于语言重复、手势产生和句子语法的流畅性也起到了必不可少的作用。KELLER et al[2]从形态学的角度描述了Broca区,他们指出,在人脑中,Broca区主要包括两个区域,即额下回的岛盖部(pars opercularis)和三角部(pars triangularis).在20世纪初期,BRODMANN[3]首次系统地描述了Broca区的细胞结构,将该区分为两个子区域,即Brodmann 44区(BA44)和Brodmann 45区(BA45).
尽管研究人员从各种损伤研究、单细胞电生理学研究和示踪剂研究中获得了广泛的信息[4-5],但伦理学禁止使用这种侵入式方法在人脑中进行研究。幸运的是,比较神经科学与磁共振技术的发展为非侵入式大脑研究带来了希望。PETRIDES et al[6]在报告中提出,猕猴腹侧前运动皮层(区域6)前面的区域是与人类BA44同源的皮质区域,他们的研究还表明,与人类BA44同源的猕猴区域前面的皮质区是与人类BA45同源的皮质区域,这块皮质区域又可以进一步细分为头部(rostral areas,45A)和尾部(caudal area,45B)两个子区域。尽管已有研究对Broca区的功能特性进行了不同角度的探讨分析,但很少有人从连接结构的角度对Broca区的白质纤维束数量进行跨物种差异性分析,本文弥补了这一缺失。
本文从大脑连接结构的角度对与语言发展密切相关的Broca区进行了跨物种一致性分析。通过使用概率纤维束跟踪技术,分别构建了人和猕猴Broca同源区与10个感兴趣脑区(region of interests,ROI)之间的连接性指纹图,同时也构建了Broca同源区与39条同源纤维束之间的连接性指纹图。此外通过运用统计学的分析方法,从样本总体均值的水平对Broca区与每一个感兴趣脑区之间的跨物种差异性进行了研究,对实验结果的可信度进行了分析。结果表明在解剖学上同源的Broca区在连接结构方面也具有较高的跨物种一致性。
由于大脑的功能与其连接结构密切相关[7],因此本研究为将猴脑作为人脑Broca区的临床研究对象提供了依据[8],对猕猴大脑Broca区的研究对于理解人脑的工作机制至关重要。
1 材料和方法
1.1 人类数据
本文从HCP数据中心发布的WU-Minn-1 200名受试者中选择了19名被试(22~35岁)[9],并从中挑选出这些被试经过数据预处理操作以后的结构像(structure magnetic resonance imaging,sMRI)数据和弥散像(diffusion magnetic resonance imaging,dMRI)数据。这批数据具有相对较高的ROI配准精度,其中,sMRI数据包括0.7 mm高分辨率各向同性T1加权(T1-weighted,T1w)和T2加权(T2-weight,T2w)图像数据;dMRI数据包括多梯度和多扩散加权的1.25 mm各向同性数据。
1.2 猕猴数据
实验中使用的猕猴数据集是来自加利福尼亚大学戴维斯分校(UC-Davis)的17只雌性猕猴(18.5~22.5岁;7.28~14.95 kg).这批猕猴数据[10]是使用Siemens Skyra 3T的4通道翻转线圈扫描收集的。所有猕猴都已进食并符合UC-Davis IACUC道德认证。在扫描之前,将猕猴用异氟烷进行麻醉,麻醉以后猕猴立即被放入扫描仪中,然后用立体定位框架固定猕猴的头部。在扫描过程中,实时监控了猴子的各种生理指标。
1.3 人类数据预处理
本文所使用的人类数据是由HCP预处理管道[11]进行预处理以后的数据。具体包括空间伪影的消除、变形矫正、皮质表面生成、交叉模式配准以及标准空间对齐等操作。在结束预处理操作以后,需要将人脑脑图谱配准到每一个被试大脑上,在被试个体脑图谱上对感兴趣脑区进行提取。图1是利用脑图谱配准法生成的被试个体大脑图谱图像。

图1 人脑图集配准到b0像结果图Fig.1 Register ration of the human brain atlas to the b0 image
1.4 猕猴数据预处理
在GLASSER et al[12]的论文中,作者概述了使来自其他来源的数据与HCP预处理管道兼容的方法。这是因为HCP预处理管道对原始数据的收集有一定的要求。其中,HCP预处理管道对dMRI数据采集的最低要求是:1) 在左右方向上对大脑进行多波段加速扫描。2) 使用Stejskal-Tanner(单极)扩散编码方案在图像中执行相位编码左右方向,即执行反向编码。3) 大量扩散梯度方向(≥128)。4) 扩散权重高达3 000 s/mm2。5) 高维空间各向同性分辨率(≤1.5 mm).
对于来自UC-Davis的猕猴数据来讲,其扩散权重只能达到1 600 s/mm2,体素分辨率则为1.4×1.4×1.4 mm.因此,HCP预处理管道不能用于对猕猴数据进行预处理操作,本文将手动对猕猴数据进行预处理。对于猕猴dMRI数据来讲,对其进行了涡流校正和非脑组织切除等预处理,下面详细介绍预处理中的脑图谱配准操作。
在配准操作之前,需要先将每一个猕猴被试的b0像从被试弥散像中提取出来,然后利用掩膜文件对其进行去除非脑组织的操作。研究表明[13],被试的T1结构像对于弥散像有很好的畸变矫正和伪影去除方面的作用,因此,本文摒弃了传统的使用FSL中的BET工具来生成b0像掩膜的思想,通过将被试的T1结构像掩膜文件(T1_brain_mask)配准到b0像上来生成b0像掩膜(b0_brain_mask)。随后,将生成的b0掩膜文件和现有的T1结构像掩膜文件配回到被试的b0像和T1像上去,分别生成了去除了非脑组织和进行了畸变矫正以后的个体T1结构像(T1_brain)和b0像(nodif_brain)。图2选取了4个被试对b0像掩膜文件的生成过程进行了展示,其中,第1列为被试T1结构像掩膜文件,第2列为被试的原始b0图像,第3列为生成的b0像掩膜文件。

图2 b0像掩膜文件的生成Fig.2 Generation of b0 image mask file
图3展示了上述的4个被试利用传统方法生成的掩膜文件与利用T1结构像生成的掩膜文件之间的对比结果,也展示了利用图2中生成的b0像掩膜去除b0像非脑组织以后的结果。其中,第2列的白色掩膜是利用FSL中的BET工具所生成的,红色掩膜是利用T1像所生成的,从图中可以明显地看出,使用BET工具生成掩膜的方法并不能把所有的头骨等非脑组织去除掉,也不会对图像的畸变和伪影进行矫正。图中第3列是将利用T1结构像掩膜文件(T1_brain_mask)生成的b0像掩膜(b0_brain_mask)配准到b0像以后的结果,新生成的b0像(nodif_brain)将用于下一步的配准操作。

图3 b0像非脑组织的去除Fig.3 Non-brain removal of b0 image
在配准过程中,首先使用线性配准将没有施加梯度磁场的b0图像(nodif_brain)配准到T1图像(T1_brain)上,然后再将T1图像(T1_brain)非线性配准到标准空间的T1像上,最后将两步的配准结果合并在一起生成最终的转换矩阵,然后利用其逆矩阵将脑图谱配准到个体b0图像上,这样就可以在被试个体水平上对感兴趣区域(region of interest,ROI)进行提取。图4是使用线性和非线性配准技术将猴脑图谱配到个体b0像以后的结果图。
图4 猴脑图集配准到b0像结果图Fig.4 Register ration of the macaque brain atlas to the b0 image
1.5 连接性指纹
PASSINGHAM et al[14]首次提出了一种基于连接性指纹的跨物种同源脑区比较的方法。他们发现大脑不同区域之间的白质纤维连接存在差异,这种差异可以作为判断大脑区域唯一性的重要标识。以人脑为例,图5分别从轴向面、冠状面和矢状面三个角度对人脑不同脑区的连接指纹进行了展示。图中黄色区域代表整个大脑,蓝色代表人脑BA44或BA45与某一感兴趣脑区之间的白质纤维连接。具体来讲,第1行展示了左脑BA44区与目标脑区之间的连接性指纹三视图,第2行展示了左脑BA45区与目标脑区之间的连接性指纹三视图,第3行展示了右脑BA44区与目标脑区之间的连接指纹三视图,第4行展示了右脑BA45区与目标脑区之间的连接性指纹三视图。从图中可以看出,大脑不同脑区与某一特定脑区之间的连接指纹存在差异,即每一个大脑区域都具有自己独特的连接模式。

图5 人脑左右脑BA44和BA45区域与选定的目标脑区之间不同的连接指纹图Fig.5 Different connectivity fingerprints between the BA44 and BA45 areas of human left and right brains and a selected target brain area
1.6 构建图谱
本文使用脑图谱配准法从人脑和猕猴被试的大脑中对感兴趣脑区进行提取。在对人脑感兴趣区域进行提取时,使用了Brainnetome图谱[15]和多巴胺中脑概率图谱[16](简称Midbrain脑图谱)。其中,Brainnetome图谱共包括左右脑246个脑区域,该图谱使用了计算机大脑作图技术对人类大脑的结构、功能以及空间变化进行了研究,具有更细粒度的解剖结构。本文分别从Brainnetome图谱中提取了A8m、TH、A8dl、dlg、A9m这5个目标脑区,从Midbrain脑图谱中提取了h32、h11m、h13、h14m、h25这5个目标脑区作为人脑感兴趣脑区进行研究。
在对猴脑感兴趣区域进行提取时,本文使用了猕猴经典D99脑图集[17]和立体定位坐标中的恒河猴脑图集[18](CIVM)来对感兴趣区域进行提取。其中,D99脑图集是基于Saleem脑图集和Logothetis脑图集生成的,该图集在猕猴左右大脑区域划分出了223个子区。CIVM图谱则是基于10个验尸后的脑标本生成的,该图谱包含242个解剖结构。本文从D99脑图谱中提取了8Bm、TH、8Bd、Ig、9m这5个目标脑区,从CIVM脑图谱中提取了h32、h11m、h13、h14m、h25这5个目标脑区作为猕猴大脑感兴趣脑区进行研究。表1和表2展示了以上10个感兴趣脑区在人脑和猴脑中的对应关系。其中,同源脑区1-5曾被用于人脑和猕猴大脑腹侧纹状体连接性的种间差异研究[19],同源脑区6-10则是基于人脑和猕猴大脑中同源脑区位置一致的假设,通过参阅有关D99图谱的书籍[20]和人脑Brainnetome图谱的资料[21]进行选取的。

表1 人脑目标脑区Table 1 Human brain target brain area
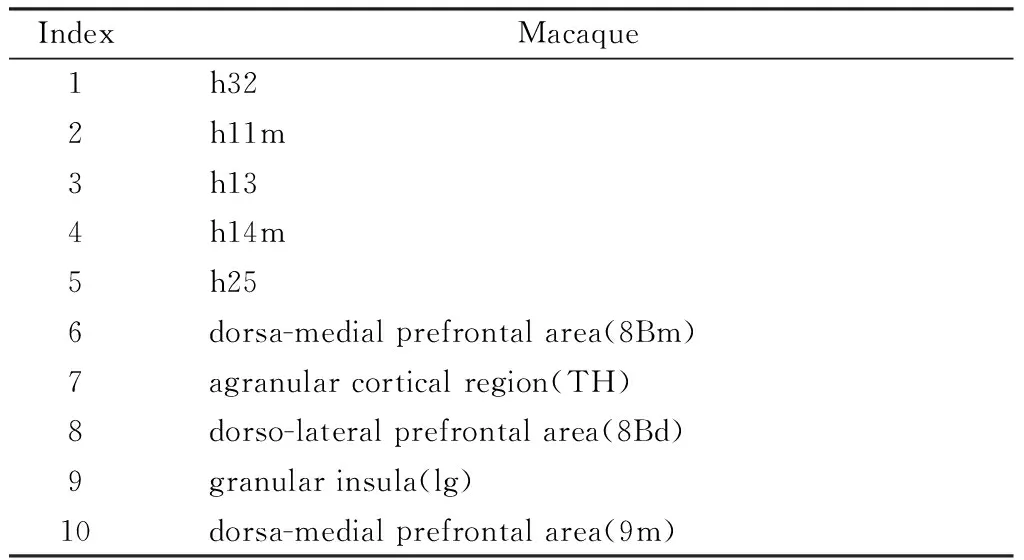
表2 猕猴目标脑区Table 2 Macaque brain target brain area
1.7 感兴趣脑区提取
本节从每一个被试大脑中对上一节所提到的10个感兴趣脑区进行提取。下面是对这10个感兴趣脑区位置的一些介绍(以人脑为例):h32位于端脑的扣带回,属于枕叶皮质区;h13、h25、h14m、h11m位于端脑的眶额叶皮层;A8m、A9m和A8dl位于大脑额上回;TH位于海马旁回;dlg位于岛回。图6展示了这10个感兴趣脑区在人脑标准脑模板和猕猴标准脑模板上的对应关系。图中左边两列的第1列为人脑1、2、3目标脑区,左边第2列为猕猴大脑1、2、3目标脑区;中间两列的第1列为人脑4、5、6目标脑区,第2列为猕猴大脑4、5、6目标脑区;右边两列的第1列为人脑7、8、9、10目标脑区,第2列为猕猴大脑7、8、9、10目标脑区。从图中可以看出,提取出来的同源脑区在人脑和猕猴大脑中具有对应的物理位置。

图6 同源目标脑区在人脑和猕猴大脑中的对应关系Fig.6 Location correspondence of homologous target brain areas in human and macaque brain
1.8 同源纤维束
在MARS et al[22]的论文中提到了一组脑白质束,即大脑中39条主要纤维束的主体。这组脑白质束可以在不同物种的大脑中被可靠地识别,因此可以用来构建整个大脑皮层的蓝图,并根据整个皮层的脑白质蓝图来描述不同物种的皮质灰质组织。
本文运用了人脑和猕猴大脑中同源的脑白质束来重新构建人脑和猕猴大脑Broca区的连接性指纹图,并将结果与同源脑区的结果进行对比,对上述实验结果进行验证。
1.9 跨物种比较
不同物种具有不同的大脑结构,即使对于同一物种,不同个体的大脑结构也不相同。为了使概率纤维追踪结果不受物种和个体大脑差异的影响,需要将得到的绝对值结果转换为相对值结果来进行跨物种比较。在以往的研究中,常用到的转换方法是使用总体的最大值、最小值或平均值来对绝对值结果进行归一化处理。本文使用总体最大连接强度(max connection strength,MaxCS)和总体最小连接强度(min connection strength,MinCS)之差来对绝对值结果进行归一化处理,将所有绝对连接强度(connection strength,CS)转换为相对连接强度(relative connection strength,RCS),公式(1)给出了具体的计算方法。其中,MeanCS代表总体平均连接强度,为了使最终的计算结果更易于比较,将乘以常数“a”来对结果进行矫正,这里将a取为1 000,即可以考虑到小数点后三位对结果的影响。
(1)
在对所有被试的连接强度做了相对值处理以后,还需要将运算结果平均化,分别计算人和猕猴所有被试在每一个感兴趣区域处的平均RCS值,将其作为物种的组平均值,最后在物种的平均水平上使用余弦相似度来进行连接性指纹的跨物种差异分析。公式(2)给出了余弦相似度FCS的计算方法,其中pi代表人脑Broca区与某一个感兴趣脑区之间的组平均RCS大小值,qi代表猕猴大脑Broca区与该感兴趣脑区之间的组平均RCS大小值。
(2)
1.10 基于小样本的统计学分析方法
本文所使用到的人和猕猴被试的数量较少,两物种被试的年龄、性别分布不均匀,而且同一物种的被试还具有一定的地域特征,样本的这些局限性会造成对最终结果刻画能力不足的问题。为了从物种整体水平对结果进行分析,也为了对结果的可信度进行分析,这里引入了统计学的分析方法。其中,独立样本t检验是通过样本均值来对总体均值进行刻画的一种分析方法,可以对Broca脑区与某一个目标脑区之间的跨物种差异性进行物种整体均值水平的刻画。排列置换测试是通过大量计算和随机排列,通过观察结果是否落在置信区间内来对结果的可信度进行分析,它解决了小样本对实验结果刻画能力不足的问题。下面对两种方法的应用进行介绍。
独立样本t检验是利用两样本均值差对总体均值差进行估计的一种统计学分析方法,主要用于样本含量较少、总体标准差未知的正态分布,本文运用该统计学方法从物种总体均值的角度对连接性指纹进行跨物种差异分析。本文假设人脑和猕猴大脑在解剖学上的Broca同源区与同一感兴趣同源脑区之间的连接性指纹在总体均值上无显著差异,在90%的置信度下,如果t统计量的P值小于显著性水平,则应拒绝原假设,即认为人脑和猕猴大脑Broca区与该感兴趣脑区之间的总体均值存在显著性差异;反之,如果t统计量的P值大于显著性水平,则应接受原假设,即认为两者的总体均值不存在显著性差异。
本文计算得到了人脑和猕猴大脑Broca区连接性指纹之间的余弦相似度值,并将其作为实验的观测值,通过对连接性指纹进行5组排列置换测试(每组100 000次随机排列置换操作),计算得出了每一次随机排列置换操作以后生成的连接性指纹之间的余弦相似度值,根据90%的置信区间大小,生成了相对应的置信区间。通过观察实验的观测值结果是否位于置信区间内来验证实验结果是否具备一定的可信度。
2 Broca区与同源脑区跨物种比较结果
本章分别对左右脑Broca区的连接性指纹结果进行了展示。其中,折线图展示了两物种连接性指纹的变化趋势,雷达图展示了两物种连接性指纹在每一个感兴趣脑区处的组平均RCS值。
2.1 Broca44区连接性指纹图
本小节对Broca44区连接性指纹结果进行了展示。图7展示了人脑和猕猴大脑左脑Broca44区的连接性指纹结果图,从折线图中可以看出,人和猕猴左脑Broca44区的连接性指纹变化趋势趋于一致。从雷达图中可以看出,两物种连接性指纹的组平均连接强度值在上下50之间进行浮动,浮动范围较小。其中,人和猕猴左脑Broca44区与感兴趣脑区2(眶额叶皮层)之间的连接性指纹差异最大,表现为人脑Broca44区的RCS要大于猕猴。

图7 左脑Broca44区连接性指纹图与排列置换测试结果Fig.7 Connectivity fingerprint and permutation test results of Broca44 area of the left brain
图8展示了人脑和猕猴大脑右脑Broca44区的连接性指纹结果图。从折线图中可以看出,人和猕猴右脑Broca44区的连接性指纹变化趋势趋于一致。从雷达图中可以看出,两物种连接性指纹的组平均连接强度值在上下60之间进行浮动,这要比左脑Broca44区的浮动范围要大。其中,人和猕猴右脑Broca44区与感兴趣脑区2(眶额叶皮层)之间的连接性指纹差异最大,表现为人脑Broca44区的RCS要大于猕猴。
2.2 Broca45区连接性指纹图
本小节对Broca45区连接性指纹结果图进行了展示。图9展示了人脑和猕猴大脑左脑Broca45区的连接性指纹结果图。从折线图中可以看出,人和猕猴左脑Broca45区的连接性指纹变化趋势趋于一致。从雷达图中可以看出,两物种连接性指纹的组平均连接强度值在上下40之间进行浮动,浮动范围较小。其中,人和猕猴左脑Broca45区与感兴趣脑区2(眶额叶皮层)和感兴趣脑区8(额上回)之间的连接性指纹差异较大,表现为人脑Broca45区的RCS要大于猕猴。

图9 左脑Broca45区连接性指纹图与排列置换测试结果Fig.9 Connectivity fingerprint and permutation test results of Broca45 area of the left brain
图10展示了人脑和猕猴大脑右脑Broca45区的连接性指纹结果图。从折线图中可以看出,人和猕猴右脑Broca45区的连接性指纹变化趋势趋于一致。从雷达图中可以看出,两物种连接性指纹的组平均连接强度值在上下50之间进行浮动,这比左脑Broca45区的浮动范围要大。其中,人和猕猴右脑Broca45区与感兴趣脑区2(眶额叶皮层)之间的连接性指纹差异最大,表现为人脑Broca45区的RCS要大于猕猴。
2.3 独立样本t检验分析
本小节用独立样本t检验方法对人脑和猕猴大脑Broca区与每一个感兴趣脑区之间的总体均值差异进行了分析。在进行独立样本t检验(置信度为90%)之前,提出了人脑和猕猴大脑Broca同源区与感兴趣脑区之间的连接性指纹总体均值无显著性差异的假设,通过对t统计量的P值进行分析来判断该假设是否成立。如果t统计量的P值大于0.1(10%)时,则认为人脑和猕猴大脑Broca区与该感兴趣脑区之间的总体均值不存在显著差异。反之,当t统计量的P值小于0.1(10%)时,则小样本事件发生,有理由拒绝原假设,即认为人脑和猕猴大脑Broca区与该感兴趣脑区之间的总体均值存在差异,这时可以根据样本均值对总体的均值差异情况进行判断。
表3展示了人和猕猴左脑Broca44区与10个目标脑区之间的独立样本t检验结果。从下表可以看出,只有h14m和A8m这两个感兴趣脑区与Broca脑区之间的t统计量所对应的P值大于0.1,即人脑和猕猴大脑Broca区与这两个感兴趣脑区之间的种间差异较小。其中,人脑Broca区与h14m区之间的连接性指纹样本均值为109,猕猴Broca区与该区的连接性指纹均值为101,相差不是很大,可以认为人脑Broca区与猕猴Broca区与该感兴趣脑区处不存在显著性种间差异。

表3 左脑Broca44区独立样本t检验结果Table 3 Independent sample t-test results in Broca44 area of the left brain
图11以折线图的形式对表3中样本的RCS均值进行了展示。从图中可以看出,人脑左脑Broca44区在大部分感兴趣脑区处的样本RCS均值都大于猕猴,这与理论是相符的。这是因为有研究表明[23],对于一只同样体重的假想猴子来讲,人脑的大小是它的4.8倍,如果将大脑的大小与大脑中白质纤维束的数量多少联系起来,人脑中同源脑区之间的白质纤维束数量要多于猕猴。PALOMERO-GALLAGHER et al[24]在其论文中也得出了随着大脑大小的增加,神经元的数量也增加的结论。除此之外,由于只有人类拥有语言能力,类人猿的语言功能与人类语言相差甚远。如果假设显微结构的差异,以及其他解剖学因素可能是造成非人灵长类动物语言能力缺乏的原因,那么这种白质纤维束的数量差异也可能是Broca同源区在不同物种中具有不同功能的基础。又因为本文是使用概率纤维束追踪技术来获取得到不同脑区之间白质纤维束数量的,所得到的结果受概率因素的影响,因而在个别脑区处出现了猴脑的RCS值大于人脑的情况,但这种情况的出现次数是非常少的。
表4展示了人和猕猴右脑Broca44区与10个目标脑区之间的独立样本t检验结果。从下表可以得出,感兴趣脑区h25的t统计量P值0.973明显大于0.1,原假设成立,即认为人脑和猕猴大脑右脑Broca44区与h25脑区之间的总体均值不存在显著差异。然后再根据样本的均值情况,人脑右脑Broca44区与h25脑区之间的平均RCS为101,猕猴大脑右脑Broca44区与h25脑区之间的平均RCS为100,从数值上确实发现两物种在h25脑区处的RCS均值十分接近。相比较h25脑区来讲,右脑Broca44区在其他感兴趣脑区处的t统计量P值都小于0.1,即认为人脑和猕猴右脑Broca44区与这些感兴趣脑区的总体连接均值存在显著差异。

表4 右脑Broca44区独立样本t检验结果Table 4 Independent sample t-test results in Broca44 area of the right brain
图12以折线图的形式对表4中样本的RCS均值进行了展示。从图中可以看出,只在感兴趣脑区4处呈现出了猴脑样本RCS均值大于人脑的情况,在其他感兴趣脑区处都呈现出人脑样本RCS均值大于猴脑的结果。
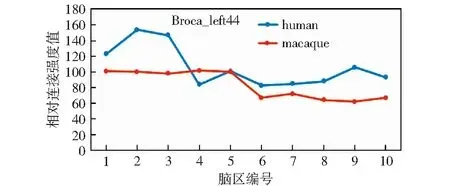
图12 右脑Broca44区样本均值比较Fig.12 Comparison of sample mean values in Broca44 area of the right brain
表5展示了人和猕猴左脑Broca45区与10个目标脑区之间的独立样本t检验结果。从下表可以得出,在感兴趣脑区h11m、A8dl/8Bd处所生成的t统计量的P值小于0.1,在其余感兴趣脑区处的t统计量的P值大于0.1,这说明了人和猕猴左脑Broca45区的连接性指纹在大部分感兴趣脑区处的总体均值不存在显著性差异。

表5 左脑Broca45区独立样本t检验结果Table 5 Independent sample t-test results in Broca45 area of the left brain
图13以折线图的形式对表5中样本的RCS均值进行了展示。从图中可以看出,只在感兴趣脑区4和9处呈现出了猴脑样本RCS均值大于人脑的情况,在其他感兴趣脑区处都呈现出人脑样本RCS均值大于猴脑的结果。

图13 左脑Broca45区样本均值比较Fig.13 Comparison of sample mean values in Broca45 area of the left brain
表6展示了人和猕猴右脑Broca45区与10个目标脑区之间的独立样本t检验结果。从下表可以得出,在感兴趣脑区h32、h14m、h25、TH处所生成的t统计量的P值大于0.1,在其余感兴趣脑区处的t统计量的P值小于0.1.

表6 右脑Broca45区独立样本t检验结果Table 6 Independent sample t-test results in Broca45 area of the right brain
图14以折线图的形式对表6中样本的RCS均值进行了展示。从图中可以看出,只在感兴趣脑区4、7、9处呈现出了猴脑样本RCS均值大于人脑的情况,在其他感兴趣脑区处都呈现出人脑样本RCS均值大于猴脑的结果。

图14 右脑Broca45区样本均值比较Fig.14 Comparison of sample mean values in Broca45 area of the right brain
2.4 排列置换测试分析
本文运用排列置换测试对实验结果的可信度进行分析。该方法基于大量计算,通过对样本进行随机排列来获取总体特征。本文分别对人脑和猕猴大脑Broca区的连接性指纹进行了5组排列置换测试,每组测试包含100 000次随机排列置换操作,最后在90%的置信度水平下,取5组测试的平均值作为置信区间。表7对Broca44区的排列置换测试结果进行了展示。从表7可以得出,人和猕猴左脑Broca44子区RCS之间的余弦相似度观测值为0.973 72,这个结果值距离左阈值0.943 44和右阈值0.996 96都比较远。人和猕猴右脑Broca44区的余弦相似度为0.978 98,尽管该测量值大于左阈值0.976 48,但距离较近,在2.1节中提到过两物种Broca44区连接性指纹左脑的浮动范围要小于右脑,这与排列置换测试的结果是一致的,即左脑Broca44区的连接性指纹距离值具有较高的可信度。总体上来讲,对于Broca44脑区,样本所反映的结果值具有较高得可信性,有理由相信人脑和猕猴Broca44脑区之间的连接性指纹差异较小,跨物种拟合程度较高。

表7 Broca44区排列置换测试结果Table 7 Broca 44 area permutation test results
表8对Broca45区的排列置换测试结果进行了展示。从表8可以得出,人和猕猴左脑Broca45区的余弦相似度为0.941 18,与左阈值0.912 35和右阈值0.995 22都距离较远。人和猕猴右脑Broca45区的余弦相似度为0.961 72,位于左阈值0.968 75的左侧,即位于置信区间外,在2.2节中提到过两物种Broca45区连接性指纹左脑的浮动范围要小于右脑,这与排列置换测试的结果是一致的,即左脑Broca45区的连接性指纹距离值具有较高的可信度。总体上来讲,对于左脑Broca45脑区,样本所反映的结果可信度较高,有理由相信人脑和猕猴大脑左脑Broca45脑区之间的连接性指纹差异较小,跨物种拟合程度较高。对于右脑Broca45脑区来讲,小概率事件发生,样本所反应的结果可信度较低。

表8 Broca45区排列置换测试结果Table 8 Broca 45 area permutation test results
上表显示右脑Broca45脑区的余弦相似度值落在阈值以外,即小概率事件发生,本文认为这与Broca45脑区的分割差异有关。这是因为,PETRIDES et al[6]在其报告中指出,猕猴大脑的Broca45脑区又可以进一步细分为头部(rostral areas,45A)和尾部(caudal area,45B)两个子区,在实际的脑区提取过程中也是按照两个子区分别从脑图谱中提取出来的。但是对于人脑Broca45脑区来讲,很少有脑模板对其进行分区,在实际的提取过程中也是将其作为一个脑区进行提取的。因此,对于人脑Broca45子区来讲,可能存在与猕猴Broca45区相似的分区情况。在接下来的实验中,我们希望在Broca45亚区水平上进行脑区连接结构的差异分析。
3 Broca区与同源纤维束跨物种比较结果
3.1 Broca44子区连接性指纹图
本小节利用人脑和猕猴大脑中的同源纤维束重新构建了Broca44区的跨物种连接性指纹图。
这里选用了39条同源纤维束中的23条来构建Broca44脑区的跨物种连接模式,这是去除了一些无用连接和极值连接以后的结果。其中,无用连接是指某些同源纤维束与所有被试的Broca44区之间的概率纤维束结果值为零,这是没有意义的,因为全零的结果并不会对最终的余弦相似度距离产生影响,因此将其去掉;极值连接是指某些同源纤维束与大部分被试的Broca44区之间的概率纤维束结果值为零,这会对最终的连接性指纹距离产生很大的影响,因此将其去掉。
图15展示了Broca44区与23条同源纤维束建立起来的连接性指纹图,其中折线图代表了两物种连接性指纹的变化趋势,从图中可以看出,两物种Broca44区在这23条同源纤维束之间的变化趋势的一致性很高,这与同源脑区的结果是一致的。雷达图则展示了人脑和猕猴Broca44区与每一个同源脑区之间的概率纤维束结果,从中也可以看出两物种Broca44脑区在每一个同源纤维处具有比较相似的轮廓。最终的排列置换测试结果也显示两物种Broca44区的连接性指纹距离落在了置信区间内,排除了小样本概率事件的发生。

图15 Broca44区与同源纤维束之间的跨物种连接模式Fig.15 Cross-species connection pattern between Broca44 region and homologous fiber bundles
通过将图15与图7,图8进行比较,不难发现,不论使用同源脑区还是同源纤维束,人脑和猕猴大脑Broca44区之间的连接性指纹拟合程度都很高,并且结果是可信的,这说明人脑和猕猴大脑在解剖学下的Broca44同源脑区也具有相似的结构连接模式,这为利用非侵入式方法来进行跨物种研究提供了依据。
3.2 Broca45亚区连接性指纹图
上一章节中利用同源脑区构建了Broca区的跨物种连接模式,结果显示右脑Broca45脑区的余弦相似度结果位于左阈值之外,即小概率事件发生。本文认为这可能是因为Broca45脑区存在亚区的关系。因此,本小节将人脑的Broca45亚区提取出来,并运用同源纤维束构建了Broca45亚区的跨物种连接模式。
图16展示了Broca45a亚区与16个同源纤维束之间的连接性指纹图,这里的16条纤维也是去除了无效连接和极值连接以后的同源纤维束。从折线图可以看出人和猕猴Broca45a亚区与16条同源纤维束之间的变化趋势具有较高的一致性,且排列置换结果也显示两物种Broca45a亚区的连接性指纹距离落在了置信区间内,排除了小样本概率事件的发生。

图16 Broca45a与同源纤维束之间的跨物种连接模式Fig.16 Cross-species connection mode between Broca45a and homologous fiber bundles
图17展示了Broca45b亚区与23个同源纤维束之间的连接性指纹图,这里的23条纤维也是去除了无效连接和极值连接以后的同源纤维束。从折线图可以看出人和猕猴Broca45b亚区与23条同源纤维束之间的变化趋势具有较高的一致性,且排列置换结果也显示两物种Broca45b亚区的连接性指纹距离落在了置信区间内,排除了小样本概率事件的发生。

图17 Broca45b与同源纤维束之间的跨物种连接模式Fig.17 Cross-species connection mode between Broca45b and homologous fiber bundles
4 结论
本文从脑区连接结构存在差异的角度,运用概率纤维束追踪的方法为人脑和猕猴大脑Broca区构建了基于同源脑区和同源纤维束的连接性指纹图。在对实验结果进行分析时,引入了统计学的分析方法,得出了人脑Broca区与猕猴大脑Broca区的连接结构拟合程度较高的结论,这为利用非侵入式手段进行跨物种比较研究提供了依据,同时也为猕猴作为人脑研究的临床对象提供了依据。虽然本文得到了一些研究成果,但仍有一些问题和工作需要进一步的研究和完善。
本文中所使用的人类数据与猕猴数据都来源于公共的数据集,对于不同的扫描参数是否会对实验结果造成影响尚不清楚。在后续的实验中,希望可以手动采集数据,在采集过程中尝试不同的扫描参数,观察最终的结果是否会有所不同。
本文仅从结构连接的角度对人脑和猕猴大脑Broca区进行了跨物种差异性分析,并没有从多模态角度[25]去对Broca区进行跨物种比较分析。后续的研究还可以从功能连接、示踪连接等角度对Broca区的跨物种一致性进行研究,并分析不同模态之间是否具有对应关系。
致谢
数据(部分)由MGH-USC联合会(主要研究者:BRUCE et al.U01MH093765)提供,由美国国立卫生研究院和人类连通症研究所的神经科学研究计划资助 基因组计划。 美国国立卫生研究院(NIH)授予编号P41EB015896; 工具赞助资金S10RR023043、1S10RR023401、1S10RR019307.
猜你喜欢
今日农业(2022年2期)2022-11-16
汉字汉语研究(2021年2期)2021-08-30
心理学报(2021年8期)2021-08-11
小猕猴智力画刊(2019年10期)2019-11-23
小猕猴智力画刊(2019年4期)2019-05-08
小猕猴智力画刊(2019年3期)2019-04-19
中学科技(2018年9期)2018-12-19
新高考·英语进阶(高二高三)(2018年8期)2018-01-15
健康管理(2017年3期)2017-04-20
读写算·高年级(2017年4期)2017-04-15
