
乡土物种补播对青藏高原高寒草甸群落稳定性的影响
2021-09-07 08:28:42段丽辉刘晓丽位晓婷才仁措邵新庆
草地学报 2021年8期
段丽辉, 刘晓丽, 韩 冰, 位晓婷, 才仁措, 邵新庆,3*
(1.中国农业大学草业科学与技术学院,北京 100193; 2.玉树州农牧业综合执法局, 青海 玉树 815000;3.青海省寒区恢复生态学重点实验室, 青海 西宁 810008)
青藏高原位于我国西北部[1]。其隆升对中国乃至亚洲的地势地貌格局、大江大河发育、气候条件形成均产生了深远影响,逐渐发育形成了独特的自然生态系统。由于该区域生态环境脆弱[2],原生植被敏感,因此植物群落受影响后自我修复和调节能力弱,极易受到干扰[3]。青藏高原植被类型丰富,以高寒草甸植被为主[4]。近年来,该区域草地出现了不同程度的退化,毒杂草丛生、植被退化、鼠害猖獗等问题频发[5],对当地牧民生产生活造成了极为恶劣的影响。
随着人们科学眼光的发展以及生活物质资料的日益丰富,草地保护、合理利用及科学管理得到了越来越多的重视。草地补播作为改良退化草地的有效途径[6],因为其成效显著、效率高而备受关注:吴宛萍[7]在宁夏荒漠草原选用乡土物种建植人工混播草地,使得草地群落地上生物量增加,群落稳定性显著提高(P<0.05);周冀琼[8]在呼伦贝尔退化羊草(Leymuschinensis(Trin.) Tzvel.)草地利用紫花苜蓿(Medicagosativa)及黄花苜蓿(MedicagofalcataL.)进行免耕补播,使草地生态系统的多样性和物种丰富度维持在较高水平,同时使草地生产力大幅提升;马和等[9]在甘肃省肃南县对因去除毒杂草后重度贫瘠的草地实施浅翻耕后补播,使得草地植被密度迅速升高,地上生物量显著增加(P<0.05)。关于补播乡土物种对于群落稳定性及种间联结性的研究较少。
为了筛选出适应青藏高原生境的生态恢复物种组合,本试验采用种间联结性及群落稳定性相结合的分析方法,选取垂穗披碱草(Elymusnutans)、异针茅(Stipaaliena)、中华羊茅(Festucasinensis)、溚草(Koeleriacristata)、星星草(Puccinelliatenuiflora)、扁蓿豆(Melissitusrutenica)、冷地早熟禾(Poacrymophila)共7种乡土物种,以5种混播组合在青海省海南藏族自治州高寒草甸天然草地进行补播。本试验为多年生草地的建植、经营与管理体系奠定了理论基础,为高寒草甸天然草地的生态保护和资源合理利用提供了理论支持。
1 材料与方法
1.1 研究区概况
本试验在青海省海南藏族自治州贵南县草原站(简称贵南站)(100°13′~101°33′E,35°09′~36°08′N)[10]的野外试验样地进行。该样地于2016年围封,长140 m,宽120 m,面积约1.9 ha。该地区属于高原大陆性气候,冬寒夏凉,年平均气温为2.3℃,雨热同期,干湿季分明而四季不分明,年降水量为403.8 mm,年平均蒸发量为1 378.5 mm[11]。植被类型为高寒草甸,土壤类型为高山草甸土[12]。该区域主要优势种包括克氏针茅(Stipakrylovii)、垂穗披碱草、矮嵩草(Kobresiahunilis)、溚草和紫花针茅(Stipacapillata)[13]。
1.2 研究方法
1.2.1试验设计与方法 本试验于2017年开展试验,选取垂穗披碱草、异针茅、中华羊茅、溚草、星星草、扁蓿豆、冷地早熟禾共7个乡土物种,搭配成5种混播组合,具体物种搭配见表1。

表1 混播组合物种组成Table 1 Species composition in combination
试验采用随机区组试验设计方案,将5种处理设置4组重复,共20个小区,单个小区面积为:5 m×4 m=20 m2,各小区间距为1 m,小区外围设有宽为1 m的保护行。在各个小区内沿长边方向将草皮划破,划沟深度为0.5~1.5 cm,行距25 cm为宜,以单个小区内分布16~17条划沟为宜。将7个草种分别依照扁蓿豆0.6 kg·km-2、异针茅0.6 kg·km-2、溚草0.325 kg·km-2、中华羊茅0.6 kg·km-2、星星草0.325 kg·km-2、垂穗披碱草1.5 kg·km-2、冷地早熟禾0.6 kg·km-2的标准,于2017年6月13日播种至相应小区内(组合E为空白对照,故仅需要沿长边划破草皮,不做补播处理。)为确保种子顺利发芽,播种后要覆土并踩实,无额外水肥投入。
1.2.2植物群落特征调查方法 对试验区的植物群落进行调查,将植物鉴定到种,根据Padisák等[14]提出的功能群分类方法划分出5个主要的功能群。此外,每个小区内,在避开单优植物斑块及边缘地块的基础上,随机选择3个面积为0.25 m2(0.5 m×0.5 m)的小样方,于2017—2018连续2年8月上旬对盖度、高度、多度和地上生物量进行测定。各指标测定方法及要求如下:盖度:利用网格法,得到群落总盖度及每个物种的分盖度[10];高度:每种植物随机选择10株并用卷尺测得高度,不足10株的物种则测定全部株的高度;多度:直接记录样方内不同种植物的株数;地上生物量:在测定得到盖度、高度和多度的基础上,对样方内不同植物种进行齐地面刈割,收集刈割得到的地上部分,装于信封袋中,置于65℃~80℃烘箱烘干,称重得到的干重即为地上生物量大小[13]。
1.3 测定指标计算方法
1.3.1群落种间联结性分析 一个植物群落中有多种植物共存,各个种群之间都产生联系、相互作用,我们将这种关系称为种间联结[15]。群落内不同种群间的关联性(或称为联结性)不仅能够客观地反映出群落内不同物种之间互作关系的有无及紧密程度[16],还能够直观地反映出群落的结构特征[17]并指示其演替方向[18]。本试验采用方差比率法(Variance ratio,VR)[19]定量测定植物群落的总体关联性。其公式如下:

(1)
(2)
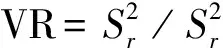
(3)
S:物种总数;Pi:包含物种i的样方数;N:总样方数;Tj:样方j的物种总数;t:样方包含物种数的均值;VR:群落内物种间的总体关联指数。
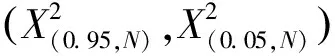
1.3.2群落稳定性分析 试验采用M.Godron贡献定率法[21](Contribution law)分析群落稳定性的变化情况。首先计算群落中所有物种的频度,并将物种按照频度从大到小的顺序排列,换算成相对频度后逐步累积,得到累积相对频度,记作y;随后将样地包含植物种类总数取倒数,依照物种排列顺序逐步对应累积,得到植物种类累积倒数百分率,记作x;最后利用成对数据拟合出一条平滑曲线,并与给定直线y=100-x相交,交点比值即可表征群落稳定性的大小[22]。一般来说,交点比值为20/80时群落稳定性最高,因此计算得到的交点坐标越接近20/80,群落就越稳定[22]。
平滑曲线模拟模型:y=ax2+bx+c
(4)
直线方程:y=100-x
(5)
将(5)代入(4)得:ax2+(b+1)x+c-100=0
(6)
交点坐标为:
(7)
y=100-x
(8)
y:物种的累积相对频度,即,将植物种类按频度按从大到小顺序排列,将频度转化为相对频度并进行累积;x为物种总数倒数的累积,即将植物种类总数取倒数并依次累积;a,b,c均为参数,算法及取值参考黄祥童[22]方法。
1.3.3生物量稳定性分析 群落生物量是群落的生长发育情况的直观表征[23],也是衡量群落稳定性大小的重要指标[24]。本试验根据Tilman[25]提出的时间稳定性计算方法计算群落的生物量稳定性。具体公式如下:
S=μ/σ
(9)
μ:群落中各个种群生物量的均值之和;σ:群落中各个种群生物量的标准差之和。
生态学界对于群落生物量的时间稳定性叫法各异,如时间序列稳定性[26],生物量稳定性[27]等。为了避免引起歧义,以下在文中一致称为生物量稳定性,记作S。但不论采取哪一种叫法,其本质都与生物统计学中的变异系数一样,能够用于不同单位、不同尺度下一组观测值变异程度的比较[27]。本试验中还把这一指标的计算方法延伸到了功能群生物量稳定性的计算中,用于探究群落生物量稳定性与功能群生物量稳定性的关系。
1.4 数据处理
采用Excel 2016进行数据处理、计算及绘图,利用SPSS 21.0软件进行统计分析,利用单因素方差分析(One-way ANOVA)方法及LSD法在显著性水平a=0.05下进行显著性检验,不同小写字母表示结果间存在显著差异(P<0.05),利用Sigmaplot 12.5进行回归分析并绘制图像。
2 结果与分析
2.1 植物群落物种组成
植物群落调查结果表明,试验区共有草本植物34种,分属于11个科(表2)。其中常见科包括禾本科(Grass)、菊科(Composite)、豆科(Legume)、莎草科(Sedge)占比超过80%。

表2 群落物种组成Table 2 Species Structure of community
2.2 种间联结性
对5种混播组合的植物群落总体关联性进行分析,结果表明(表3):2017年仅组合A的群落总体表现为显著正联结;其余4个组合植物群落总体均表现为不显著正联结。2018年,组合B、组合C和组合E群落总体表现为不显著负联结;组合A、组合D群落物种总体表现为不显著正联结。
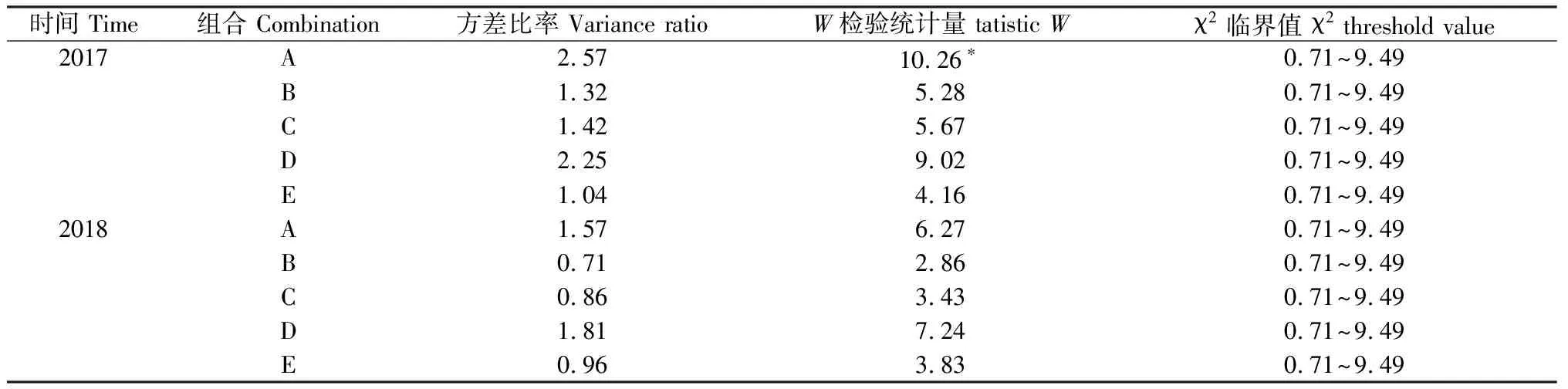
表3 群落物种联结性计算结果Table 3 Results to community species association
2.3 植物群落稳定性
由图1可知,2017年及2018年5种混播组合整体群落的种类累积倒数百分率x与累积相对频度y的比值均远离20/80。但每个组合先后两年的补播结果相比之下,2018年的图像交点更接近20/80(图1)。

图1 不同组合群落稳定性的分析Fig.1 Analysis of community stability of different combinations
2.4 群落生物量稳定性
试验结果表明,5种混播组合的群落生物量稳定性排序为D>A>C>E>B。其中组合B与组合D的差异显著(P<0.05),其余组合两两之间不存在显著差异(图2a)。
本试验根据Padisák等[14]提出的功能群分类方法,将观察到的物种划分为禾本科、豆科、菊科、莎草科、杂类草(Forb)5个功能群。采用Tilman的时间稳定性计算方法[25]对功能群的生物量稳定性和群落整体的生物量稳定性进行计算,并建立直线回归方程进行相关性分析。结果表明(图2b~2f):群落生物量稳定性与禾本科生物量稳定性之间存在显著的相关关系(R2=0.633,P<0.01),而与莎草科、菊科、豆科、杂类草的生物量稳定性之间均不存在显著的相关关系(豆科:R2=0.19,P=0.053;菊科:R2=0.044,P=0.376;杂类草:R2=0.81,P=0.223;莎草科:R2=0.002,P=0.947)。
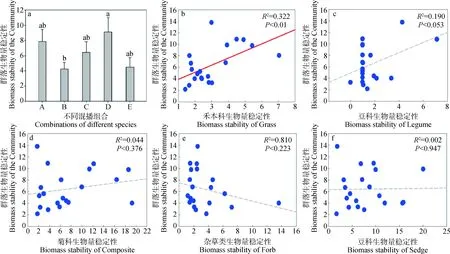
图2 群落生物量稳定性与功能群生物量稳定性的关系Fig.2 Relationship between biomass stability of the community and biomass stability of the functional group
3 讨论
3.1 种间联结性与群落稳定性
种间联结性是群落稳定性的长期背景,群落稳定性是种间联结性的阶段性表达[22]。通常情况下,种间正关联程度越大,群落越稳定,种间关系越紧密,相互促进越多;相反,种间负关联程度越大,种间竞争越激烈,群落稳定性越小[28]。有关种间联结性与群落稳定性间相互关系的研究有很多:庞晓攀等[29]对高原鼠兔有效洞口数密度,同高山嵩草(KobresiapygmaeaC. B. Clarke)草甸植物群落种间联结性之间的关系进行了一系列研究,发现植物群落主要物种总体联结性由正联结转变为负联结时对应的有效洞口数密度下,嵩草草甸植物群落稳定性最高;张鲜花等[30]对天山北坡不同海拔地段鸭茅(DactylisglomerataL.)群落主要物种进行了一系列研究,发现随海拔升高,鸭茅群落主要物种种间联结性由正关联逐渐过渡到负关联,同时群落稳定性上升到一定程度后减弱。本研究观察到,5种混播组合所得群落整体种间正关联程度降低而负关联程度升高的同时,群落稳定性升高。这与上述研究[29-30]结论一致,推测群落接下来将会向着顶级群落方向发生演替,其稳定性也将会进一步上升,并在群落总体种间联结性由正关联转变为负关联时达到最大。
对于群落种间正关联程度降低这一现象,可能是因为试验选用的混播组合较简单,且可利用资源与营养在试验初期较充沛,因此物种间竞争较弱,从而表现出较好的种间相容性,继而呈现出较好的种间联结性。而乡土物种的加入对原生植物群落产生扰动,尽管它们具有优良的植物学特性、对环境适应能力强[31-33],仍然会对原生群落的种间联结性产生负效应。而对于群落稳定性提高这一现象,推测是由于补播处理使土壤有效养分呈现出表聚性[34],使根系可利用资源量增加;以及随着植株生长,叶层高度及覆盖度出现差异[34],这两者综合作用导致的。这有利于生物成层及资源利用效率的升高,最终使得群落稳定性增大。
综上所述,在进行补播对植物群落稳定性的影响评价时,可将种间联结性分析方法与群落稳定性分析方法相结合,二者所得结论能够相互印证,并在一定程度上对群落演替方向做出指示,具有很大实践意义。
3.2 生物量稳定性
群落生物量是群落的生长发育情况的直观表征[23],生物量稳定性则是衡量群落稳定性大小的重要指标[24]。试验结果表明,5种组合的群落生物量稳定性存在差异,其中组合D(扁蓿豆+星星草+垂穗披碱草+异针茅+溚草)的生物量稳定性最高,这可能是因为该组合兼具禾草及豆草,其中豆科植物根系入土深,能够与根瘤菌共生并完成氮素固持[35];而禾本科植物根系入土浅,比表面积大,利于养分吸收[35]。这种根系分布及作用特点的差异使得组合D植物群落得以维持较高的资源分配效率及生物量稳定性,进而获得较高的群落稳定性。
此外,研究结果还表明,群落生物量稳定性与禾本科功能群生物量稳定性之间存在极显著的相关关系(R2=0.633,P<0.01),而与其它功能群的生物量稳定性不存在显著的相关关系。推测可能是以下几点原因引起的:首先,选用的7个乡土物种中有6个为禾本科植物,试验样地自身包含的34种植物中禾本科便占据了10种,故试验设计本身便体现了禾本科对于整体的重要作用;其次,基于1971—2010年的气候监测数据[36],青藏高原在近40年里整体气温升高了约0.5℃,而禾本科植物对增温十分敏感,增温有助于其纵向生长、截取更多光照资源[37],并促进根系向下生长寻求水分[38];最后,禾本科植物自身较高[37],而高度又是地上生物量的决定因素之一。综上所述,诸多因素共同作用,确定了禾本科生物量稳定性同群落总体生物量稳定性间极显著的相关关系(P<0.01)。
4 结论
研究表明,在高寒草甸天然草地选用5种乡土物种混播组合进行补播降低了群落种间联结性、提高了群落稳定性及生物量稳定性。5种组合中,组合D(扁蓿豆+星星草+垂穗披碱草+异针茅+溚草)对群落生物量稳定性的提高最多,是生态恢复的最优组合。这可能与该混播组合为豆、禾混播,对资源的利用率高、配给合理有关。本试验跨度仅为2年,仅能得到群落稳定性的初步变化趋势,要想得到天然草地选用乡土物种混播组合进行补播对草地植物群落稳定性的长期影响,还需要更长时间来探索。
猜你喜欢
草地学报(2022年11期)2022-12-01 12:15:40
现代畜牧科技(2021年11期)2021-12-21 06:11:24
现代畜牧科技(2021年7期)2021-07-28 06:40:30
草食家畜(2021年5期)2021-03-27 22:35:12
草原与草业(2020年1期)2020-04-09 10:08:36
西藏农业科技(2019年1期)2019-07-25 00:37:00
草地学报(2017年6期)2017-03-24 16:40:32
湖南城市学院学报(自然科学版)(2016年4期)2016-02-27 14:02:41
杂草学报(2015年2期)2016-01-04 14:57:54
农产品市场周刊(2015年39期)2015-10-31 19:12:27
