
赤霉素对入侵植物凤眼莲营养生长和克隆繁殖的调控
2021-09-06 06:59:06陶贵荣
生物安全学报 2021年3期
许 静,李 晶,陶贵荣
西安文理学院生物与环境工程学院,陕西 西安 710065
凤眼莲Eichhorniacrassipes(Mart.)Solms又称水葫芦、凤眼蓝、布袋莲,属于单子叶雨久花科凤眼莲属,是一种原产于南美洲亚马逊流域的多年生飘浮型水生植物,是已知生长繁殖速度最快的植物之一,目前在我国南方10多个省份都有分布,且极有可能向北方地区蔓延(郑志鑫等,2018; Jonesetal.,2018; Zhangetal.,2010)。由于凤眼莲生长繁殖快且天敌很少,已在很多地方造成严重的生态入侵问题,但凤眼莲又具备净化污染水体、可用作发酵底料等优点,因此,需要科学防控和合理利用凤眼莲(丁建清等,1995; 徐寸发等,2018; Aboul-Eneinetal.,2011; Sharmaetal.,2016; Yanetal.,2017)。成熟的凤眼莲植株有8~12片基生叶,合轴生长,叶片深绿色,呈肾圆形,须根发达,叶柄中下部中空膨大成葫芦状,植株可以漂浮于水面(Penfound & Earle,1948)。凤眼莲形态具有较高可塑性,如植株密度过大时,膨大茎会消失,叶柄呈细直状,高温或低光照强度会造成叶柄增长,叶片形状也会受一些环境因子调控有所变化(高雷和李博,2004)。凤眼莲的繁殖分为有性和无性繁殖,但因凤眼莲的种子非常细小,在自然状态下萌发率很低,造成其有性繁殖效率低下,因此,凤眼莲主要依靠产生克隆株的方式进行无性繁殖(张迎颖等,2012)。Watson(1984)曾提出凤眼莲叶基长出腋芽并发育成克隆株,克隆株与母株以匍匐茎相连,匍匐茎极易受外力影响折断,克隆株即可脱离母株成为一个独立的植株并漂浮到其他水域。
植物激素在植物整个生活史中都起到重要的调控作用。虽然凤眼莲具备生态学和经济学双重重要性,但关于其生长发育和克隆繁殖调控的研究,尤其是激素对其的调控研究甚少(Villamagna & Murphy,2010)。目前已知人工合成的生长素2,4-D 对凤眼莲有一定的毒性,可作为凤眼莲的除草剂使用,而另一种生物源的生长素吲哚乙酸和细胞分裂素在凤眼莲腋芽内的比值与其克隆株的生长有相关性,且赤霉素被发现可以协同促进2,4-D 对凤眼莲的毒杀作用(牛佳等,2012; Pieterse & Roorda,1982)。赤霉素最早被发现是因赤霉菌Gibberellafujikuroi分泌的一种双萜类化学物质造成水稻OryzasativaL.茎徒长的症状,导致水稻恶苗病。随后的研究发现,赤霉素在高等植物中普遍存在,并在植物的整个生活期都有重要调控作用(高秀华和傅向东,2018)。Pieterseetal.(1976)发现低浓度(0.1444~2.8871 μmol·L-1)的赤霉素会造成凤眼莲膨大茎消失,叶柄直立细长,且克隆株的生长受到抑制。相似的,Watsonetal.(1982)在广口瓶里用添加了不同浓度(0.2887~2.8871 μmol·L-1)赤霉素的固体培养基培养凤眼莲幼苗,发现赤霉素会造成凤眼莲叶柄成细直状,膨大茎消失,但该研究结果显示,低浓度的赤霉素会促进凤眼莲产生更多的克隆株。这些结果说明,赤霉素对凤眼莲的生长和无性繁殖均存在一定调控作用,但这些研究观察检测的生理指标并不全面,而且使用的赤霉素浓度均较低,对于调控的克隆株结论也不一致。鉴于植物激素存在浓度依赖性,同一种激素在高浓度和低浓度时对植物的调控作用并不相同(De vleesschauweretal.,2014),所以,这些结果并不能完全呈现出赤霉素对凤眼莲营养生长和克隆繁殖的调控作用。因此,本研究通过对凤眼莲外源施用较高浓度的赤霉素,并检测其形态指标,包括株高、根长和膨大茎周长,叶片的长宽和形态、叶片SPAD值以及茎上组织根重量和根重、根冠比,克隆株数量等指标,以明确高浓度赤霉素对凤眼莲营养生长和克隆繁殖的影响及调控机理。
1 材料与方法
1.1 试验材料
凤眼莲植株取自江西贵溪信江流域,将凤眼莲植株置于温度(20±5)℃、湿度70%的温室中,自然光照条件下,培养扩繁得到大量来源同一母株的克隆株系,将新长出的有5~6片叶的小克隆株从母株剥离,作为本研究的试验材料。
1.2 试验方法
1.2.1 凤眼莲的培养 将凤眼莲植株置于40 cm×32 cm×12 cm的塑料盆中,用改良霍格兰营养液[KNO3、NH4H2PO4、NH4Cl、MgSO4、Ca(NO3)2、FeSO4、EDTA、CuSO4、ZnSO4、H3BO4、MnCl2和(NH4)6Mo7O24浓度分别为1、0.25、0.1、0.5、1、0.025、0.025、0.0003、0.00033、0.0115、0.0035和0.0001 mmol·L-1]进行培养,每盆4株,每盆装5 L营养液,每周更换一次营养液。
1.2.2 激素的施用 本研究施用的激素为赤霉酸(gibberellic acid, GA3)。用天平称取0.6925 g GA3粉末,滴入几滴乙醇溶解后加蒸馏水定容至20 mL配制成100 mmol·L-1的母液,用0.22 μm过滤器灭菌后置于-20 ℃保存,使用时吸取定量的母液用无菌水稀释成50 μmol·L-1工作液,每盆用40 mL稀释后的激素均匀喷洒在植株的茎叶表面,以喷洒无菌水为对照,每周喷洒一次直至收样。
1.2.3 营养生长指标的测定 培养4周后,测量植株的株高、根长、膨大茎直径、叶片数量、叶片的长和宽、叶片SPAD值和氮含量。其中,测量膨大茎直径时每株选取3片最年幼的已经发育成熟叶片的叶柄最粗部位进行测量,并测定这3片叶片的长宽、SPAD值和氮含量。测定生物量时,先取出植株晾干至表面无明显水分,再称重。
1.2.4 无性繁殖指标的测定 培养结束后,清点每株凤眼莲上克隆株数量并用尺子测量匍匐茎长度。
1.3 数据处理与分析
本研究中试验重复2次,每次试验每个处理3盆,每盆4株植物。使用Excel进行数据分析和图表制作,采用T-test方法检验处理间差异是否显著,显著水平为P<0.05。
2 结果与分析
2.1 赤霉素对凤眼莲植株形态的影响
如图1所示,与喷洒无菌水的对照相比,外源喷洒50 μmol·L-1GA3的凤眼莲叶柄明显增长,整体株高为对照组的2倍多,根系也明显变短,膨大茎周长远小于对照组,基本不膨大。这些结果与文献中报道的低浓度赤霉素对凤眼莲植株形态的调控结果相符(Pieterseetal.,1976; Watsonetal.,1982),说明本研究的高浓度激素处理有效,且高浓度与低浓度处理对凤眼莲植株形态的影响基本一致。GA3处理对凤眼莲叶片数量不产生明显作用(表1)。
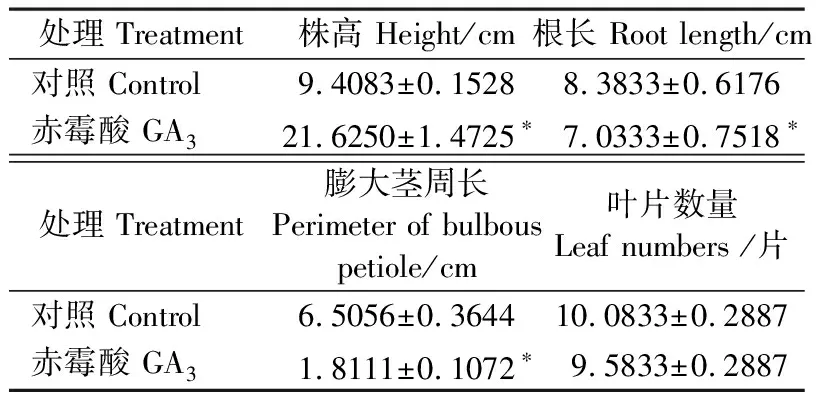
表1 赤霉素对凤眼莲形态相关指标的影响

图1 不同处理下凤眼莲植株的形态
2.2 赤霉素对凤眼莲叶片发育的影响
如图2所示,正常情况下,凤眼莲的叶片呈肾圆形,颜色为深绿色,而50 μmol·L-1GA3处理后的叶片呈心形,虽然叶片长度基本不变,但宽度变窄,叶片整体面积变小,且颜色由深绿变为浅绿。叶片SPAD值和氮含量的测定结果(表2)表明,50 μmol·L-1GA3处理后的叶片SPAD值和氮含量均显著低于对照组。这些结果说明赤霉素抑制了凤眼莲叶片的发育。

图2 不同处理下凤眼莲叶片的形态

表2 赤霉素对凤眼莲叶片生长的影响
2.3 赤霉素对凤眼莲生物量累积的影响
从表3可知,50 μmol·L-1GA3处理下凤眼莲的根系和茎上组织都显著低于对照组,但根冠比无明显差异,说明赤霉素抑制了凤眼莲生物量的累积,但对生物量在茎上组织和根系之间的分配无明显作用。

表3 赤霉素对凤眼莲生物量累积的影响
2.4 赤霉素对凤眼莲克隆繁殖的影响
由表4可知,对照组在第3周开始出现克隆株,平均每株克隆株数为0.5个,到第4周基本每株都有1个克隆株,克隆株与母株相连的匍匐茎平均长度为5.36 cm;而50 μmol·L-1GA3处理下的凤眼莲直到第4周仍未出现发育成熟的克隆株,说明赤霉素对凤眼莲克隆株产生有显著抑制作用。

表4 赤霉素对凤眼莲产生克隆株的影响
3 讨论
内源赤霉素含量的增加或外源施用赤霉素都会造成植物茎或叶柄增长,从而使株高增高,而赤霉素合成突变体植物一般出现矮小表型(高秀华和傅向东,2018)。Pieterseetal.(1976)和Watsonetal.(1982)的研究均表明,低浓度赤霉素促进凤眼莲叶柄的纵向生长,抑制膨大茎的发育。本研究使用高浓度的赤霉素出现了同样的表型:叶柄增长,膨大茎消失。这一结果说明,无论赤霉素浓度高低,都能造成凤眼莲叶柄细直且纵向增长,最终出现株高增高的表型特征。除了对株高的调控外,赤霉素对叶片发育也有重要调控作用,但作用因植物种类而异。首先,赤霉素可改变植物叶片形态,如番茄LycopersiconesculentumMill.叶片经赤霉素处理后叶缘由锯齿状变光滑,而胡椒PipernigrumL.叶片的叶面则由光滑变粗糙(Gray,1957)。本研究表明,叶面喷洒高浓度赤霉素使凤眼莲叶片形状从肾圆形变为心形,说明赤霉素同样对凤眼莲叶片的形状有重要影响。其次,赤霉素还能影响叶片的长宽和叶面积,如大豆Glycinemax(Linn.)Merr.和豌豆PisumsativumL.等植物叶片的长度都会因赤霉素处理变长,而水稻等单子叶植物的叶片除了长度变长外,其宽度还有可能变窄,但叶片的叶面积整体变大(Brain,1959; Gray,1957; Wheeler & Humphries,1964)。对同为单子叶植物的凤眼莲而言,赤霉素确实使叶片宽度变窄,但长度基本没有变化,所以整体叶面积变小。由于植物叶片基部有分生组织,赤霉素可通过促进细胞分裂或增加细胞长度而造成叶片伸长(Brain,1959; Gonzaleetal.,2012; Nelissenetal.,2012),但是凤眼莲叶片基部连接的是膨大茎,膨大茎下方才有分生组织,这也是赤霉素可以促进凤眼莲茎伸长而不是叶片伸长的原因。此外,赤霉素对叶片颜色也有影响。赤霉素下游信号重要调控因子DELLA的突变体植株颜色比野生型更加深绿(Pengetal.,1997),而外源施用赤霉素的植物叶片绿色更浅(Kuraishi & Hashimoto,1957)。同样的,本研究表明,施用赤霉素使凤眼莲叶片颜色更浅,SPAD值和氮含量更低。目前,关于赤霉素信号通路对叶绿素生物合成的调控机制已基本明确,DELLA通过与 SCL(scarecrow-like)蛋白的结合解除了SCL对抑制原叶绿素酸脂氧化还原酶编码基因PORC表达的抑制作用,从而促进了叶绿素的合成。因此,外源赤霉素通过抑制DELLA的累积实现了对叶绿素合成的抑制(Maetal.,2014)。
赤霉素对植物生物量的影响因植物种类而异,目前并没有一致的结论。本研究结果表明,经赤霉素处理的凤眼莲虽然根冠比与对照无差别,但茎上组织鲜重和根鲜重均低于对照组,说明赤霉素对凤眼莲生物量的累积起抑制作用,这可能是以下几个原因造成的:一是赤霉素抑制了凤眼莲叶片的发育,造成其叶面积减小和叶绿素含量减少,从而降低了光合作用效率并最终使得生物量累积速度减慢;二是赤霉素抑制了凤眼莲克隆株的产生,由于本研究计算的重量包括了母株和克隆株的重量,因此赤霉素处理的植株的重量远低于对照植株的重量;三是由于赤霉素处理过的凤眼莲植株没有膨大茎,造成植株不能正常浮于水面,而是部分茎叶沉入水中并最终溃烂,因此损失了部分重量。
本研究结果表明,赤霉素处理显著抑制了凤眼莲的克隆株产生,这与Pieterseetal.(1976)的研究结果一致,但和Watsonetal.(1982)的结论刚好相反。Watsonetal.(1982)研究中提到赤霉素对凤眼莲克隆株产生的调控存在浓度依赖性,浓度在1.4435 μmol·L-1时对凤眼莲克隆株产生的促进作用最强,在其试验的最高浓度2.8871 μmol·L-1时促进作用开始减弱,但Pieterseetal.(1976)的研究中赤霉素浓度同样为低浓度(0.2887~2.8871 μmol·L-1),而本研究参考了同为单子叶的模式植物水稻中赤霉素常用的外源喷洒浓度50 μmol·L-1(De vleesschauweretal.,2016),约为另外2个研究中最高浓度的17.3倍。可见,本研究与另外2个研究中使用的赤霉素浓度的差异不太可能是造成不同研究中赤霉素对克隆株产生的调控作用不一致的原因,真正的原因可能是植株培养方式和激素施用方法的不同。在Pieterseetal.(1976)和本研究中,凤眼莲的培养都是模拟自然条件,将植株置于宽阔的塑料盆中用液体营养液培养,由于赤霉素处理使得凤眼莲膨大茎消失,植株无法正常浮于水面,新发的腋芽一直泡在营养液中,也极有可能导致其无法正常发育成成熟的克隆株,而Watsonetal.(1982)是将凤眼莲幼苗置于1 L的玻璃瓶中用培养基培养,不存在新发腋芽无法暴露于空气中的问题。另外,二者施用激素的方式也不同,Pieterseetal.(1976)和本研究是将赤霉素喷洒在叶茎表面,而Watsonetal.(1982)是将赤霉素添加在培养基中经由根系吸收,施用方式的不同可能造成吸收部位和效率的不同,进而影响最终的功效。值得注意的是,Watson(1984)曾提出凤眼莲每个叶片的叶腋处都能产生新的腋芽进而发育成克隆株,可见,腋芽生长对凤眼莲克隆繁殖至关重要。而赤霉素对腋芽生长的影响却有完全相反的报道,如Nietal.(2016)发现赤霉素无论浓度高低都能促进桐油树腋芽的生长,其机制牵涉与细胞分裂素协同调控腋芽的细胞分裂(Nietal.,2017),而Zhangetal.(2019)却报道赤霉素依赖DELLA-SPL9蛋白复合体抑制拟南芥腋芽的产生。由此可见,在植物体内赤霉素通过不同的机制分别促进或抑制腋芽的生长,而这些机制之间如何相互作用并形成最终的生理功效却不得而知。因此,今后应继续深入研究赤霉素对凤眼莲腋芽生长的调控机制,进而明确其对凤眼莲克隆繁殖的作用。
凤眼莲是世界上已知生长繁殖速度最快的植物之一,其快速生长繁殖的调控因子和有针对性的防控策略仍有待更多的研究。本研究发现,对模拟自然条件下生长的凤眼莲喷洒赤霉素可改变其植株形态,抑制叶片发育,最终造成生长和无性克隆繁殖速度减缓,这为其未来的防控提供了一定参考。
猜你喜欢
亚热带植物科学(2023年5期)2024-01-27 10:38:00
吉林蔬菜(2022年4期)2022-11-04 07:22:20
山东农业科学(2022年11期)2022-03-07 05:57:10
陕西农业科学(2020年10期)2020-11-13 12:47:20
中国园林(2020年4期)2020-05-23 08:47:38
华声文萃(2019年12期)2019-09-10 07:22:44
文萃报·周二版(2019年43期)2019-09-10 07:22:44
中国蔬菜(2018年1期)2018-01-11 03:39:14
浙江大学学报(农业与生命科学版)(2015年4期)2015-12-15 12:47:44
新疆农垦科技(2014年7期)2014-02-28 19:20:29
