
Modulating poststroke inflammatory mechanisms:Novel aspects of mesenchymal stem cells,extracellular vesicles and microglia
2021-09-03 03:10WenQiangXinWeiWeiYongLiPanBaoLongCuiXinYuYangMathiashrThorstenDoeppner
World Journal of Stem Cells 2021年8期
Wen-Qiang Xin,Wei Wei,Yong-Li Pan,Bao-Long Cui,Xin-Yu Yang,Mathias Bähr,Thorsten R Doeppner
Wen-Qiang Xin,Wei Wei,Yong-Li Pan,Bao-Long Cui,Mathias Bähr,Thorsten R Doeppner,Department of Neurology,University Medical Center Göttingen,Göttingen 37075,Germany
Xin-Yu Yang,Department of Neurosurgery,Tianjin Medical University General Hospital,Tianjin 300052,China
Abstract Inflammation plays an important role in the pathological process of ischemic stroke,and systemic inflammation affects patient prognosis.As resident immune cells in the brain,microglia are significantly involved in immune defense and tissue repair under various pathological conditions,including cerebral ischemia.Although the differentiation of M1 and M2 microglia is certainly oversimplified,changing the activation state of microglia appears to be an intriguing therapeutic strategy for cerebral ischemia.Recent evidence indicates that both mesenchymal stem cells(MSCs)and MSC-derived extracellular vesicles(EVs)regulate inflammation and modify tissue repair under preclinical stroke conditions.However,the precise mechanisms of these signaling pathways,especially in the context of the mutual interaction between MSCs or MSC-derived EVs and resident microglia,have not been sufficiently unveiled.Hence,this review summarizes the state-ofthe-art knowledge on MSC- and MSC-EV-mediated regulation of microglial activity under ischemic stroke conditions with respect to various signaling pathways,including cytokines,neurotrophic factors,transcription factors,and microRNAs.
Key Words:Extracellular vesicles;Mesenchymal stem cells;Microglial activation;M2 polarization;Ischemic stroke
INTRODUCTION
Inflammation plays an important role in the pathophysiology of ischemic stroke,and systemic inflammation worsens the outcome of stroke patients[1-3].Preclinical models of focal cerebral ischemia are associated with the activation of inflammatory cells,including neutrophils,T cells,and resident microglia.The latter cells contribute to both immune defense and brain tissue repair[4].Under physiological conditions,microglia are resting and maintain a balance with regard to the inflammatory status of the brain[5].Ischemic stroke immediately activates microglia,prompting an inflammatory response in brain tissue[5].Consequently,microglia migrate toward the location of the lesion and exacerbate brain damage by secreting inflammatory cytokines.On the other hand,microglia can remove debris and secrete anti-inflammatory cytokines,supporting endogenous brain repair[4,6].These opposing roles of microglia under ischemic conditions correlate with distinct phenotypes,as indicated by the proinflammatory M1 type and the anti-inflammatory M2 type[7,8].M1-type microglia exacerbate brain damage by secreting interleukin(IL)-6,IL-1β,nitric oxide(NO),tumor necrosis factor(TNF)-α,and other factors,whereas M2-type microglia promote brain repair by secreting IL-4,IL-10,and transforming growth factor(TGF)-β[9,10].Changing the activation state of microglia might therefore be an intriguing approach for stroke therapy.
Regulating poststroke immune responses offers novel therapeutic strategies even for patients who qualify for neither systemic thrombolysis nor endovascular thrombectomy[11,12].Hence,modulating poststroke immune responses and stimulating endogenous repair mechanisms through the transplantation of adult stem cells,such as mesenchymal stem cells(MSCs),has gained increasing interest in recent years.In fact,preclinical work,as well as clinical trials,revealed the efficacy and tolerance of grafted MSCs in stroke settings[13-15].However,MSCs do not typically integrate into residing neural networks within the ischemic hemisphere but rather act indirectly.Such paracrine effects are mediated,at least in part,by extracellular vesicles(EVs).EVs are small vesicles in the range of 30 nm to 1000 nm that are secreted by virtually all eukaryotic cells and contain a plethora of proteins,noncoding RNAs,and DNA[16,17].MSCs and MSC-EVs affect a great number of intracellular and extracellular signaling pathways,among which are immunoregulatory cascades.
Despite the aforementioned data on the effects of MSCs and EVs on poststroke inflammation,the precise mechanisms underlying such a therapeutic approach are unknown.Although some evidence suggests that bone marrow-derived MSCs and MSC-EVs may inhibit microglial activation and improve neurological function under pathological conditions,including stroke[18-21],data on the interaction between MSCs or MSC-EVs and microglia are limited.In this review,we summarize the therapeutic effects of MSCs and MSC-derived EVs both in preclinical studies and in clinical stroke trials.We also summarize the mutual interactions between MSCs or MSC-EVs and activated microglia under such stroke conditions.
THE ROLE OF MICROGLIA IN ISCHEMIC STROKE
Microglial activation in ischemic stroke
Under physiological conditions,microglia display a ramified structure characterized by a small soma and fine processes to maintain homeostasis within the extracellular milieu[22,23].Stroke results in microglial activation,which represents the first step in an inflammatory response that is followed by the activation of other immune cells,such as T cells,neutrophils,and natural killer cells[24,25].When activated after stroke,microglial cells undergo four distinct phenotypes:ramified,intermediate,amoeboid,and round[26].Ramified microglia indicate the resting state,whereas intermediate microglia have larger cell bodies and shorter bumps.The amoeboid microglial cell body,on the contrary,is larger and displays shorter bumps or even no bumps at all,similar to round microglia,which are found in the lesion center[26].Based on these morphological characteristics and their secretion patterns,microglia are characterized as M1 or M2.
Correlation between morphological phenotypes and microglial function in cerebral ischemia
Modifying microglial morphology changes cellular functions such as the production of cytokines[27].The immediate activation of microglia due to cerebral ischemia results in a change in cell size,as well as in different migration and secretory properties[28].Classic(M1 type)and alternatively(M2 type)activated microglia are most commonly reported after ischemic insult[28].M1 microglia participate in the proinflammatory tissue response and are able to present antigens,whereas M2 microglia remove necrotic tissue and stimulate tissue repair,thus maintaining homeostasis by producing anti-inflammatory substances.Phenotypic shifts between the M1 and M2 types,therefore,has practical implications,and promoting an M2 phenotype would assist tissue repair by decreasing inflammatory factors.
Phenotypic shifts between M1 and M2 microglia involve a plethora of characteristics for which not only the aforementioned phenotype but also the secretion patterns of these cells are important.Hence,proinflammatory M1 microglia typically secrete factors such as IL-1β,interferon(IFN)-α,IL-6,cyclooxygenase-2(COX-2),motif chemokine ligand(CXCL10),and inducible NO synthase(iNOS)[29-32].In contrast,anti-inflammatory M2 microglia have the capacity to produce and secrete IL-10,IL-13,IL-4,insulin-like growth factor(IGF)-1,and IFN-β[33,34].Of note,based on the current knowledge on M2-polarized microglia in the central nervous system,the M2 phenotype is further categorized as M2a,M2b,or M2c based on cellular function[35,36].M2a microglia are strongly associated with IL-13 and IL-4 and exert strong antiinflammatory effects.These cells also produce significant amounts of arginase-1,Ym-1,CD206,and Fizz1[36],while the M2b phenotype does not have the capacity to produce the latter[35,37].The M2c phenotype,also known as deactivated microglia,is associated with promoting tissue regeneration at a later stage of disease when inflammation is declining[38,39].
Mechanisms and role of M2 microglial activation in ischemic stroke
A large number of studies have been performed to identify the main path ways and mediators that modulate M2 microglial activation.Two primary transcription factors,c-AMP response element-binding protein(CREB)and nuclear factor-κB(NF-κB),are strongly associated with signaling pathways for M2 microglial polarization.Activated microglia,for instance,display reduced expression of COX-2 by suppressing the activation of NF-κB[40,41].In addition to the aforementioned signaling cascades,other pathways,such as Toll-like receptor 4(TLR4)[30],CD8[42],and mitogen-activated protein kinase(MAPK),are also involved in promoting the polarization of the M2 phenotype,as indicated by preclinical stroke studies[43].Typically,the pathways and mediators responsible for M2 microglial activation interact with each other to some extent,and they tend to work synergistically rather than independently to achieve the maximum anti-inflammatory effect.Some of the signaling pathways associated with the polarization of the M2 phenotype are summarized in Figure 1.

Figure 1 Signaling pathways describing the process of polarization of the M2 phenotype.Damaged neurons induced by ischemic stroke activate resting microglia by producing damage-associated molecular pattern molecules.M1 phenotype microglia participate in the proinflammatory tissue response,whereas M2 microglia produce antiinflammatory substances.DAMPs:Damage-associated molecular pattern molecules;PPAR:Peroxisome proliferator-activated receptor;CREB:c-AMP response element binding protein;NF-κB:Nuclear factor-κB;YM1:Chitinase-3 Like protein;CD:Cluster of differentiation;STAT:Signal transducer and activator of transcription;ROS:Reactive oxygen species;Arg-1:Arginase-1;TNF:Tumor necrosis factor;iNOS:Inducible nitric oxide synthase;TLR:Toll-like receptor;IGF-1:Insulin-like growth factor-1;IL:Interleukin;IFN:Interferon;FIZZI:Found in inflammatory zone 1;TGF:Transforming growth factors;MAPK:Mitogenactivated protein kinase.
Blood-brain barrier(BBB)disruption is significantly involved in the pathology of ischemic stroke[44,45].BBB disruption allows extracerebral substances to reach the brain parenchyma in an unregulated way,worsening brain tissue damage and inducing brain edema.The early stage of BBB disrupture occurs at 12-24 h after stroke exposure,followed by a second delayed phase at approximately 48-72 h poststroke,during which microglia are activated,which,in turn,also affects BBB integrity.Microglia initially protect BBB integrity by promoting the levels of the tight junction protein claudin-5.During sustained inflammation,however,activated M1 microglia are able to phagocytose astrocytic end-feet and destroy BBB integrity by secreting various vascular proteins[30,46].Of note,M1 microglia induce endothelial necroptosis and impair BBB integrity by expressing TNF-α as a primary mediator of these effects[47].
Based on current studies,M2 microglia may attenuate BBB disruption by producing progranulin,which is helpful in preventing brain edema[48].To date,studies revealing a direct interaction between microglial activation and BBB disruption have been scarce.In this context,the stroke-associated release of reactive oxygen species(ROS)also activates microglia,which further worsens the integrity of the BBB[49].Likewise,the postischemic upregulation of NF-κB in microglia stimulates the these cells to secrete matrix metalloproteinases,which play key roles in the disruption of the BBB[50].
MSCS AND EVS FOR THE TREATMENT OF ISCHEMIC STROKE
The biological concept of MSCs and EVs
In the 1960s and 1970s,Friedensteinet al[51]first revealed that ectopic transplantation of rodent bone marrow cells into the kidney capsule has osteogenic effects.It was not until 1991,however,when Caplanet alrecommended the term “mesenchymal stem cells” due to the capacity of these cells to differentiate into various cell lineages[52].Interestingly,researchers were able to establish other tissue sources of MSCs,including adipose tissue[53,54].In 2006,the International Society for Cellular Therapy proposed specific MSC criteria due to some controversy about the characteristics of MSCs[55].These criteria include adhesion to the substrate in question,the ability to express the surface antigens CD73,CD90,and CD105,the absence of proteins such as CD14,CD34,CD79a,CD11b,CD45,and human leukocyte antigen-11 and the potential to differentiate into adipocytes,osteoblasts,and chondroblasts[55,56].To date,MSCs have been widely studied in various preclinical models and clinical settings alike,and these studies have focused on cell migration patterns and immunosuppressive functions[57-62].
MSCs,however,act in an indirect way,which is supported by the fact that transplanted MSCs rarely reach the ischemic lesion site after intracerebral injection and even less so after systemic transplantation[63,64].Rather,the majority of MSCs are trapped in peripheral organs such as the lung after systemic transplantation.In addition,neural differentiation,synaptogenesis,and reconstruction of neural network systems are time-consuming processes,but MSC transplantation is known to also yield quick therapeutic results within days[65].Current evidence suggests that grafted MSCs are not integrated into residing neural networks but act in an indirect paracrine way,including the secretion of trophic factors such as brain-derived neurotrophic factor(BDNF),IGF,and vascular endothelial growth factor(VEGF)[66].In addition to these soluble factors,however,recent evidence suggests that EVs secreted from MSCs might significantly contribute to exerting their biological effectsin vitroandin vivo.
EVs are secreted by virtually all cells into the extracellular matrix,which is regarded as a novel mechanism of intercellular communication.EVs are a heterogeneous group of vesicular structures ranging in size from approximately 40 nm to 1000 nm,among which are endosome-derived exosomes[16,67].EVs carry highly active biological cargo such as transmembrane proteins,RNA,and various lipids[68].This mix of cargo is thought to mediate the biological properties of EVs and indirectly of MSCs under both physiological and pathological conditions.
Application of MSCs or MSC-EVs results in tissue regeneration upon the induction of ischemic stroke
Although the focus of the present review is on poststroke inflammation and the mutual interaction between MSCs/MSC-EVs and microglia,it stands to reason that both MSCs and MSC-EVs increase neurological recovery though mechanisms that are partly independent of MSCs.In this context,the modulation of poststroke neurogenesis,angiogenesis,and axonal plasticity is of importance.
MSCs are able to improve neurological function by promoting astrocyte-derived IGF-1,epidermal growth factor(EGF),VEGF,and basic fibroblast growth factor(bFGF)[69].Indeed,many preclinical stroke studies have demonstrated the positive effects of MSCs on both neurogenesis and angiogenesis[19,70].Increased tissue regeneration after MSC transplantation is at least partially a consequence of secreted factors such as VEGF and Ang-1[69,71].Under conditions of hypoxia,neurons that synthesize γ-secretase increase the production of Hes-1 and activate the Notch-1 signaling pathway after MSC administration,which in turn may promote the production of hypoxia inducible factor(HIF)-1α and VEGF[69].Hes-1 can further stimulate this effect by inducing a positive feedback loop with VEGF by reducing phosphatase and tension homolog levels[72].Consistent with this,Ang-1 helps stabilize new vessels induced by VEGF expression[71].
Not only neurons but also oligodendrocytes are responsive to MSC treatment.Oligodendrocytes are also sensitive to ischemic cell injury and play an important role in the neural network.However,these cells have long been a neglected target in current stroke research[73].Some data,however,indicate that stimulating the production of myelin sheaths by mature oligodendrocytes through MSC transplantation may result in increased axonal plasticity after stroke[74].These neurorestorative effects are mediated by inhibiting the production of both reticulin and neurocan.The aforementioned mechanisms and signaling pathways associated with poststroke tissue regeneration have to be regarded as prominent examples only.Other factors,such as hepatocyte growth factor,platelet-derived growth factor,BDNF,IGF-1,fibroblast growth factor-2,and neutrophil-activating protein 2(NAP-2),are also involved[75,76].An overview of how MSCs promote neurogenesis and neurological recovery is shown in Figure 2.

Figure 2 An overview as to how mesenchymal stem cells promote neurogenesis and neurological recovery.Mesenchymal stem cells(MSCs)are isolated and identified from various tissue sources.These MSCs produce different angiogenic and neurotrophic factors by paracrine mechanisms in order to act on astrocytes,microglia,oligodendrocytes,and angiogenesis,promoting neurological recovery and neurogenesis.IGF-1:Insulin-like growth factor 1;BDNF:Brainderived neurotrophic factors;VEGF:Vascular endothelial growth factors;HGF:Hepatocyte growth factor;NAP-2:Neutrophil activating protein 2;PDGF:Plateletderived growth factor;Ang-1:Angiopoietin-1;MSC:Mesenchymal stem cell.
As previously described,stem cell-derived EVs(MSC-EVs)and other factors induce tissue regeneration after ischemia in various organs,including the heart,kidney,and brain.Under such conditions,EVs not only reduce cell injury but also promote angiogenesis and neurogenesis.Some of these observations are related to regulating inflammation[77-79].In fact,previous work from our group suggests that EVs from different stem cell sources,such as MSCs and neural progenitor cells,are not inferior to their host cells with regard to their therapeutic potential in a mouse stroke model[80,81].Various preclinical stroke studies report positive effects of MSC-derived EVs on infarct volume and tissue recovery.To date,a series of preclinical studies[20,21,80,82-103]have assessed the effect of MSC-derived EVs on treating cerebral ischemia.The characteristics and primary outcomes of some of these studies are summarized in Table 1 and demonstrate that MSC-EVs are immunologically active and promotetissue repair and neurogenesis[104-107].

Table 1 Therapeutic application of mesenchymal stem cell-derived extracellular vesicles in preclinical disease models associated with cerebral ischemia
MSCs and MSC-EVs suppress microglial activation and promote M2 microglial polarization in cerebral ischemia
PubMed,the Cochrane Library(last searched in November 2020),and relevant websites,such as Web of Science and EMBASE(1990 to November 2020),were searched to identify all studies on the effect of MSCs and MSC-derived EVs,including MSC-derived exosomes,on microglial activation in the treatment of brain ischemia.The following keywords in combination with Boolean logic were used:“mesenchymal stem cell”,“extracellular vehicles” or “exosomes” together with “ischemia”,“stroke”,“microglia”,“middle cerebral artery occlusion”,or “MCAO”.Beyond this,the reference list was manually checked to determine other potentially qualified trials.The process was iterated until no more publications were obtained.A total of 25 publications were found in this section from the United Kingdom,Japan,China,United States,France,Germany,Poland,and South Korea,which were performed between 2013 and 2020[19-21,84-87,91,99,102,108-122].The most common species and animal model used were rats and the middle cerebral artery occlusion model,respectively.The most common source of cells and administration route were bone marrow and intravenous delivery,respectively.Cunninghamet al[109]was the only group that used subcutaneous injection.Concerning the dose and delivery time,the studies used heterogeneous experimental paradigms to meet their own applied study purposes.Additional details are shown in Table 2.
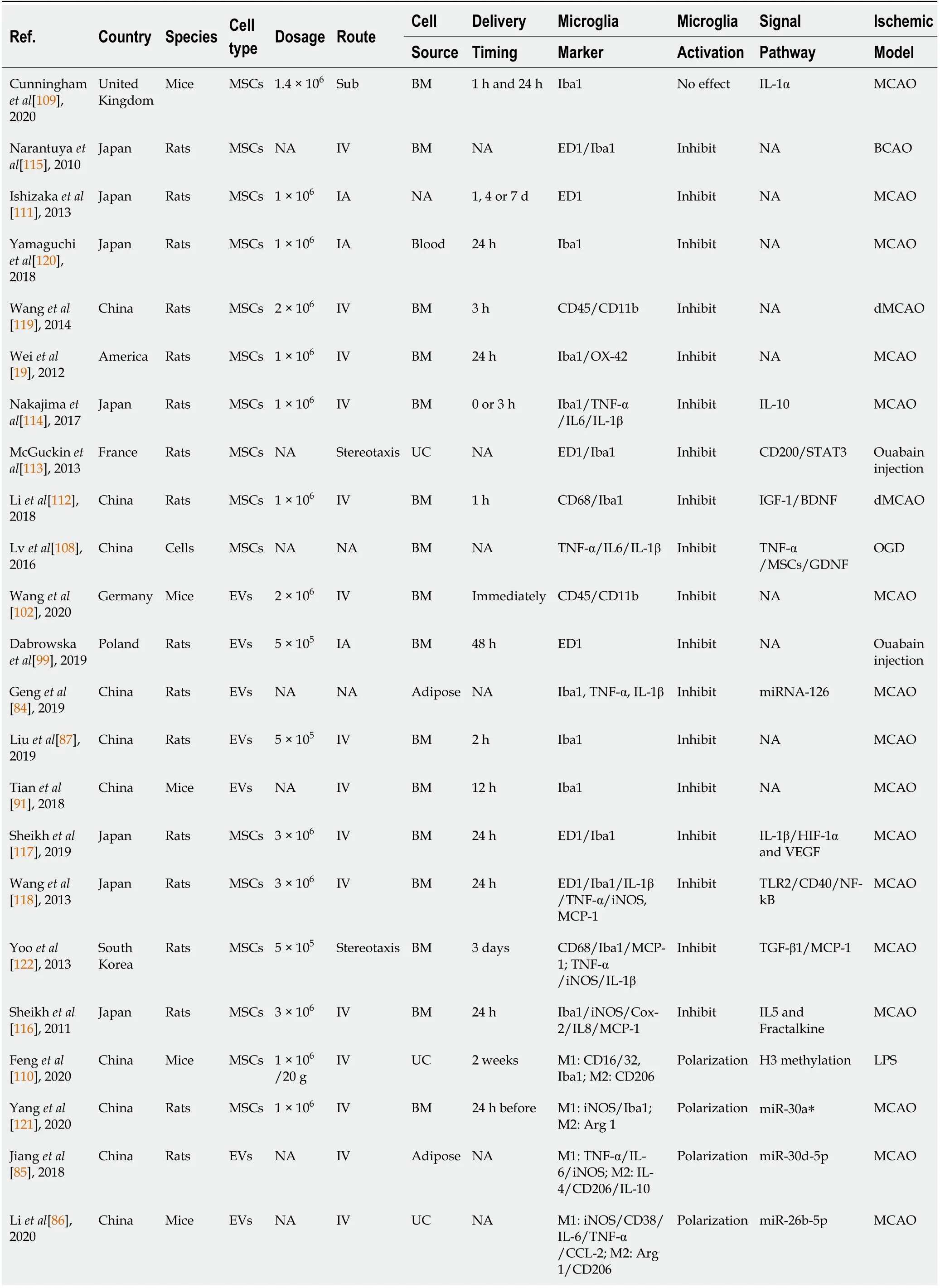
Table 2 Preclinical stroke studies assessing the effect of mesenchymal stem cells and mesenchymal stem cell-derived extracellular vesicles on the activation of microglia

NA:Not available;IA:Intraarterial;IV:Intravenous;Sub:Subcutaneous;BM:Bone marrow;UC:Umbilical cord;OGD:Oxygen-glucose deprivation;HIF-1α:Hypoxia inducible factor-1α;dMCAO:Distal middle cerebral artery occlusion;MSCs:Mesenchymal stem cell;EVs:Extracellular vehicles;LPS:Lipopolysaccharide;BCAO:Bilateral carotid artery occlusion;c(RGDyK):Cyclo(Arg-Gly-Asp-D-Tyr-Lys);MCP-1:Monocyte chemoattractant protein-1;iNOS:Inducible nitric oxide;IL:Interleukin;NF-κB:Nuclear factor-κB;TLR:Toll-like receptor;VEGF:Vascular endothelial growth factor;TGF:Transforming growth factor;IGF:Insulin-like growth factor;BDNF:Brain-derived neurotrophic factor;TNF:Tumor necrosis factor;GDNF:Glial-derived neurotrophic factor;COX-2:Cyclooxygenase-2;CysLT2R:Cysteinyl leukotrienes receptor;ERK:Extracellular regulated protein kinase.
The immediate activation of brain-resident immune cells,mainly microglia,is the primary characteristic of inflammatory reactions after ischemic stroke[4,23].Activated microglia produce neurotoxic substances that accelerate acute brain damage,and some of these neurotoxic substances reciprocally promote further microglial activation.Attenuating microglial activation with MSCs or MSC-EVs offers great therapeutic potential.The transplantation of MSCs(2 × 106)into rats 3 h after focal cerebral ischemia,for instance,yielded a significant reduction in macrophages at day 3 after treatment[119].Likewise,the expression of OX-42+or Iba-1+microglia was significantly reduced after the administration of MSCs during the acute and subacute stages of the disease[19].Increased neurological recovery after stroke induction due to MSC transplantation,which had repeatedly been observed before,— is partly a consequence of an increase in the ratio of M2/M1 microglia[121].This finding is consistent with similar reports on MSC-EVs,in which tissue restoration was enhanced due to the suppression of M1 microglial polarization after MSC-EV treatment[20,21,85,86].Of note,the aforementioned effects of MSCs or MSC-EVs are not always sustained;the positive effects of treatment are sometimes only observed during the acute or subacute phase of the disease[109,115]Thus,the ability of MSCs to suppress microglialactivation appears to be limited over time,and additional and reliable data are urgently needed.
The role of neurotrophic factors and cytokines in regulating microglial activity after MSC or MSC-EV treatment in ischemic stroke
Systemic or intrathecal injection of growth factors such as BDNF and IGF-1 promotes angiogenesis and reduces infarct volume in ischemic stroke[123].However,these proteins do not cross the BBB and are prone to rapid degradation,preventing longterm effects in the ischemic milieu.MSC transplantation might therefore overcome some of these limitations.Indeed,rats that received MSC transplantation displayed increased levels of IGF-1 and BDNF in the ischemic cortex[112].Increased secretion of these factors is likely to be a consequence of microglial activity,which was found to be increased in the ischemic core site.Likewise,ischemic stroke itself increases the expression of monocyte chemoattractant protein-1(MCP-1)and activates CD68-positive microglia to cross the damaged BBB.Consistent with this finding,Yooet al[122]observed that TGF-β is key to the ability of MSCs to effectively reduce the infiltration of CD68-positive microglia into the ischemic zone by downregulating MCP-1.
The amount of cytokines and growth factors that are secreted directly by MSCs or that are modulated in microglia due to stem cell transplantation is vast and cannot be discussed in full detail here.Thus,other evidence provides insights into MSC production of neurotrophic cytokines such as IL-5 and fractalkine,which in turn suppress the production of proinflammatory factors,including iNOS and TNF-α[116].In a mutual interaction between MSCs and activated microglia,the latter secrete TNFα to stimulate the production of glial cell-derived neurotrophic factor in MSCs,which in turn prevents neuronal damage and contributes to tissue repair[108].Changes in the extracellular milieu due to MSC transplantation are well known and suggest increases in HIF-1α,VEGF,IL-1β,and TGFβ protein levels,all of which are highly expressed in microglia[108,114].Thus,MSC transplantation also promotes poststroke angiogenesis by regulating HIF-1α and VEGF secretion by microglia,and IL-1β might play an important role[117].
IL-1,IL-6,and TGF-β are proinflammatory factors associated with the immune response after ischemic stroke.It has been reported that the levels of IL-1 are increased within a few hours after ischemic damage.Increased IL-1 Levels,in turn,stimulate the secretion of other cytokines,chemokines,and cell adhesion molecules that contribute to the disruption of the BBB[124].Recent literature indicates that the transplantation of MSC-EVs reduces IL-1 secretion by microglia due to direct inhibition and modification of T helper cells,which drive microglia into an anti-inflammatory state[99,125].Dabrowskaet al[99]revealed that the transplantation of MSC-EVs significantly reduced the levels of IL-6 and TGF-β in the focally injured rat brain through the inhibitory effect of EVs on local immunologically effective cells,such as microglia and macrophages.Other work describes that CH25H,a hydroxylating enzyme that alters cholesterol into its 25-hydroxycholesterol form,is activated by proinflammatory cytokines such as IL-1β,TNF-α,and IL-6[126].CH25H,in turn,exacerbates cerebral inflammation and significantly activates Iba-1-positive microglia[127].MSC-EVs might therefore contribute to suppressing microglial activation by inhibiting IL-1β,TNF-α,and IL-6,further reducing CH25H activation.Information regarding this phenomenon,however,is scarce.
The role of transcription factors,ligands and surface receptors in regulating microglial activity after MSC or MSC-EV treatment in ischemic stroke
The effects of TLR4 on worsening ischemic injury have been discussed for over a decade[128].Reduced TLR4 inhibit the NF-κB signaling pathway and decrease the expression of iNOS and COX-2[129].NF-κB is known to promote various proinflammatory mediators,and the inhibition of NF-κB signaling has beneficial effects on cerebral ischemia[130].Wanget al[118]showed that MSC transplantation suppressed the NF-κB signaling pathway in microglia during stroke due to soluble factors such as TNF-α and prostaglandin E2 produced by MSCs.In addition,TLR2 and CD40 have important roles in modulating NF-kB pathways activation,and the expression levels of these factors are lower after MSC treatment,as described in the same scientific report.Previously,it was reported that spontaneous microglial activation occurs in mice with CD200 knockout[113].Neurons have been shown to produce CD200,which in turn suppresses the activity of microglia that produce CD200R.McGuckinet al[113]found that IL-4 induced MSCs to produce CD200 when cocultured with activated glia,which suppressed the expression of IL-6 and IL-1β in glial cells.Furthermore,the ability to modulate CD200 expression was successfully reversed by anti-IL-4 and anti-CD200 antibodies.Inflammation in ischemic stroke is also related to STAT3 signaling pathway activation in microglia,which is affected by MSCs in cerebral ischemia[113].
With regard to the administration of MSC-EVs significantly inhibits stroke-induced inflammation and M1 microglial polarization,increases the expression of anti-inflammatory factors and enhances the polarization of M2 microglia.Finally,MSC-EVs regulate the expression of phosphorylated ERK 1/2 and CysLT2R,which were downregulatedin vitroandin vivo[20].
MicroRNAs are key players in regulating microglial activity
MicroRNAs(miRNAs),a family of noncoding RNAs containing 20-25 nucleotides,play key roles in the remodeling process under stroke conditions[131].Current evidence has revealed that miRNAs are effective treatment candidates due to their capacity to promote angiogenesis and neuronal recovery in ischemic diseases.Studies have demonstrated the anti-inflammatory effects of specific miRNAs that are highly expressed in MSCs and MSC-EVs.
MSCs carrying miR-30a and EVs carrying miRNA-126,miR-30d-5p,miR-26b-5p,or miR-223-3p modulate microglial activation and anti-inflammatory abilities[21,84-86,121].EVs exert a regulatory effect by delivering prewrapped miRNAs to recipient cells.As discussed previously,CysLT2R is involved in the regulation of microglial activation.EV-derived miR-223-3p increases functional recovery after cerebral ischemia by promoting microglial M2 polarization because of its inhibitory effect on CysLT2R[21].Similarly,miR-26b-5p promotes microglial M2 polarization by regulating CH25H to repress the TLR pathway and reduce tissue injury[86].Both stroke patients and rats exhibited significantly reduced levels of miRNA-126,and EVs containing miRNA-126 could suppress microglial activation and increase neurogenesis and angiogenesis[84].In addition,some EVs that carry miR-30d-5p have increased potential to suppress neuronal damage by inhibiting autophagy-mediated M1 microglial polarization[85].Similarly,miR-30a*(known as miR-30a-3p)also participates in several pathways to drive M2 polarization[121].Hence,a plethora of miRNAs are found in MSCs and their corresponding EVs,but the precise signaling cascades that are regulated under stroke conditions are not yet fully known.
MSCs and MSC-EVs in clinical stroke trials
Taking into account the numerous preclinical reports on MSCs and stroke,as well as the easy access to these cellsviathe bone marrow or adipose tissue[132-134],MSCs are important in the novel adjuvant treatment paradigm against stroke.In fact,MSC transplantation is considered to be safe.Apart from transient febrile reactions,no research has reported signs of intoxication,thrombogenesis,central nervous system deterioration,or increased mortality after MSC transplantation in humans[135,136].
Rigorous evaluation of the literature available on the effects of MSCs on stroke patients published in electronic databases(PubMed,Cochrane Library,and EMBASE)until January 31,2021 yielded a total of 18 studies including 631 participants[14,137-153].Of these 631 patients with ischemic stroke,323 patients were treated with MSCs,and 308 were assigned to the control group.Prasadet al[144]organized a clinical stroke trial on MSCs with the primary endpoint as the modified Rankin scale(mRS)and Barthel Index(BI)score that includes the highest number of patients enrolled to date:a total of 120 patients with ischemic stroke were equally assigned to either the MSC group(n= 60)or the control group(n= 60)[144].MSCs were intravenouslyadministered at a dose of 280.5×106cells at a median of 18.5 d after stroke onset.Patients who received MSC treatment displayed better outcomes according to the BI and the National Institute of Health Stroke Scale(NIHSS)score at a one-year followup.Similarly,Savitzet al[137]found that intravenous autologous transplantation of bone marrow mononuclear cells(8.5 × 107)significantly improved the BI,mRS,and NIHSS scores without any side effects.Although long-term observations are still rare,some data indicate that MSC treatment is associated with a lower rate of mortality compared to that of the control group during a 5-year observation period[144].The data showed that MSC treatment can increase neurological recovery in stroke patients,even though additional data are urgently needed.Table 3 summarizes recent clinical trials on stroke and MSC transplantation.Although MSC-EVs have been shown to be as effective as MSCs in improving functional outcomes in preclinical stroke studies[154],to date,no article has reported the use of EVs under clinical stroke conditions.Therefore,more evidence-based information is needed in this respect.

Table 3 The key parameter of clinical trials using transplantation of mesenchymal stem cells in treating ischemic stroke
CONCLUSION
The application of MSCs and MSC-EVs offers a great opportunity for adjuvant stroke treatment,and inflammation is an excellent target.The differentiation of M1 and M2 microglia has practical implications,and promoting the M2 phenotype enhances tissue repair by decreasing inflammatory factors.Currently,the application of MSCs and MSC-EVs appears to inhibit microglial activation and promote M2 polarization,which results in the modification of various signaling pathways,such as cytokines,neurotrophic factors,transcription factors,and miRNAs.
 World Journal of Stem Cells2021年8期
World Journal of Stem Cells2021年8期
- World Journal of Stem Cells的其它文章
- Differences and similarities between mesenchymal stem cell and endothelial progenitor cell immunoregulatory properties against T cells
- Inter-regulatory role of microRNAs in interaction between viruses and stem cells
- Mesenchymal stem cells for enhancing biological healing after meniscal injuries
- Antler stem cells and their potential in wound healing and bone regeneration
- Therapeutic prospects of mesenchymal stem/stromal cells in COVID-19 associated pulmonary diseases:From bench to bedside
- Mesenchymal stem cells as a potential therapeutic tool to cure cognitive impairment caused by neuroinflammation