
真蛸染色体核型分析*
2021-08-25 07:04李凤辉史会来周丽青陈四清葛建龙刘长琳
渔业科学进展 2021年5期
李凤辉 边 力 史会来 周丽青 陈四清① 葛建龙 刘长琳 卢 斌 常 青
(1. 上海海洋大学水产科学国家级实验教学示范中心 水产动物遗传育种中心上海市协同创新中心 上海水产养殖工程技术研究中心 上海 201306;2. 中国水产科学研究院黄海水产研究所 农业农村部海洋渔业可持续发展重点实验室 山东 青岛 266071;3. 青岛海洋科学与技术试点国家实验室海洋渔业科学与食物产出过程功能实验室 山东 青岛 266071;4. 浙江省海洋水产研究所 浙江省海水增养殖重点实验室 浙江 舟山 316021)
真蛸(Octopus sinensis, d’Orbigny, 1841)隶属于软体动物门(Mollusca)、头足纲(Cephalopoda)、二鳃亚纲(Dibranchiata) 、 八 腕 目(Octopoda) 、 蛸 科(Octopodidae)、蛸属(Octopus),主要分布于北太平洋西部的温带浅海海域,特别是中国、韩国和日本沿海,成体营底栖生活(Amoret al, 2017; Gleadall, 2016)。O. sinensis过去常与分布于大西洋和地中海等海域的真蛸(Octopus vulgaris)视为同种异名(Iglesiaset al,2007; Okumuraet al, 2005; Warnkeet al, 2004)。近年来,基于分子生物学和外部形态特征,在物种水平上将O. sinensis和真蛸(O. vulgaris)分离开来,英文命名为East Asian common octopus (O. sinensis)(Amoret al,2019、2017; Gleadall, 2016)。鉴于国内没有重新定名,本文暂沿用真蛸。蛸类是海洋大型经济鱼类的重要饵料,作为捕食者和被捕食者,在海洋生态系统食物链中扮演着重要角色;其味道鲜美,营养丰富,备受消费者青睐,具有较高的经济价值(廖永岩, 2006)。随着头足类海洋捕捞量的逐渐减少和消费量的逐渐增大,蛸类人工养殖将是必然,市场前景广阔(叶守建等, 2014)。O. sinensis具有生命周期短、产卵量大、食物转化率高和生长速度快等特点,是极具潜力的海水增养殖品种(Danet al, 2019)。目前,中国、日本等国家已经开始进行O. sinensis人工养殖技术的研究(Danet al, 2019; 徐大凤等, 2019)。
染色体是生物遗传信息的载体,是两性生物进化、遗传变异的物质基础,研究染色体数目和核型,对了解生物遗传变异规律、亲缘关系、进化地位、性别决定机制等均具有重要意义(史宝等, 2017; Amoreset al, 2014)。同时,可为生物的种质鉴定、雌(雄)核发育、多倍体诱导、杂交育种等技术的研究以及染色体水平的基因组组装提供参考依据(Molina-Luzónet al,2015)。现存的头足类大约有700 余种,但对其染色体核型的研究相对较少。目前,国内外仅对10 种头足类的染色体进行了研究:金乌贼(Sepia esculenta)、虎斑乌贼(Sepia pharaonis)、拟目乌贼(Sepia lycidas)、莱氏拟乌贼(Sepioteuthis lessoniana)、长枪乌贼(Heterololigo bleekeri)、剑尖枪乌贼(Photololigo edulis)、商乌贼(Sepia officenalis)、短蛸(Octopus ocellatus)、长蛸(Octopus minor)和真蛸(王晓华等, 2011; Papanet al, 2010; Gaoet al, 1990; Vitturiet al, 1982; Inaba,1959)。目前,国内外对O. sinensis的研究主要集中在分类、发育生物学以及人工养殖技术等方面,对其染色体核型的研究未见报道(Amoret al, 2017、2019;Gleadall, 2016; Danet al, 2019)。本研究对O. sinensis的染色体核型进行分析,旨在丰富其细胞遗传学数据,以期为其种质鉴定、资源保护和人工养殖等研究提供理论基础。
1 材料与方法
1.1 实验材料
实验用真蛸为2019 年10 月在浙江省舟山市附近海域捕获的野生群体幼体,根据右侧第3 腕足确定其雌雄(图1B、C),雌雄各5 只,雌性体重为(169.25±4.03) g,全长为(34.05±3.46) cm,生殖腺指数为1.31±0.62;雄性体重为(345.60±5.32) g,全长为(41.20±4.35) cm,性腺指数为2.31±0.89。按照Amor 等(2017)的研究,基于线粒体COI 基因部分序列对随机选取的5 只章鱼进行分子生物学鉴定,鉴定结果为O. sinensis(图1D)。实验在浙江省舟山市浙江省海洋水产研究所试验基地进行。实验开始前,车间暂养30 d,暂养水温为22℃~24℃,盐度为30~33,pH 为7.8~8.4,DO>6 mg/L。饲以菲律宾蛤仔(Ruditapes philippinarum)和日本蟳(Charybdis japonica),日投饵1 次,投饵量为体重的 5%~10%,采用流水养殖,日换水量为100%~200%。
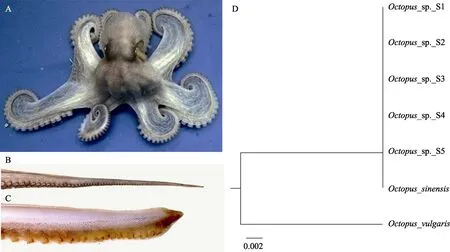
图1 O. sinensis 外部形态及分子生物学鉴定Fig.1 External morphology and molecular identification of the O. sinensis
1.2 染色体标本制备
染色体标本制备参照金乌贼染色体核型分析方法(王晓华等, 2011),略作修改。预处理:将O. sinensis活体置于含秋水仙素终浓度为0.005%的过滤海水中暂养2 h,暂养水温为22℃~24℃,暂养过程中持续充气。处理:解剖并取鳃和肾脏组织于过滤海水配制的秋水仙素溶液(终浓度为0.04%)中浸泡30 min。低渗:将处理后的鳃和肾脏组织切成小块,并置于50 ml 浓度为0.075 mol/l 的KCl 溶液中(蒸馏水配制)处理50 min。固定:将低渗后的组织转移至预冷的卡诺氏液(甲醇∶冰醋酸=3∶1)中,每隔15 min 更换固定液,重复3 次。固定后的组织4℃保存过夜。解离:固定后的组织于50%的冰醋酸(蒸馏水配制)中解离,解离时间视组织解离状态而定,待解离组织近乎透明,终止解离,剔除未解离的组织,并用吸管轻轻吹打解离液2 min。滴片:采用热滴片法制备染色体标本。染色:载玻片在空气中自燃干燥后,用10%的吉姆萨染液染色30 min,蒸馏水冲洗,晾干后镜检。整个实验过程雌雄各重复5 次。
1.3 染色体核型分析
使用Nikon ECLIPSE 80i 显微镜对O. sinensis染色体标本进行观察并拍照。选取200 个染色体分散较好的中期分裂相进行数目统计。再从中选取20 个着色较好、轮廓清晰、收缩适中的染色体中期分裂相经放大打印后进行测量和分析,并计算相对长度和臂比。相对长度=(实测染色体长度/全部染色体长度总和)×100;臂比=长臂长度/短臂长度;按照Levan 等(1964)提出的标准进行核型分析,按臂比将染色体分为4 种类型:中部着丝点染色体(m,臂比1.00~1.70)、亚中部着丝点染色体(sm,臂比1.71~3.00)、亚端部着丝点染色体(st,臂比3.01~7.00)、端部着丝点染色体(t,臂比≥7.0)。中部着丝粒染色体(m)和亚中部着丝粒染色体(sm)臂数为2,亚端部着丝粒染色体(st)和端部着丝粒染色体(t)臂数为1。
2 结果与分析
2.1 染色体数目的确定
利用O. sinensis的鳃和肾脏组织均能获得中期分裂相(图2A),雌雄各选取100 个分散较好的分裂相进行数目统计(表1)。统计结果显示,200 个分裂相中,染色体条数为60 的分裂相有156 个,占比为78%,由此确定O. sinensis的二倍体染色体条数为2n=60。
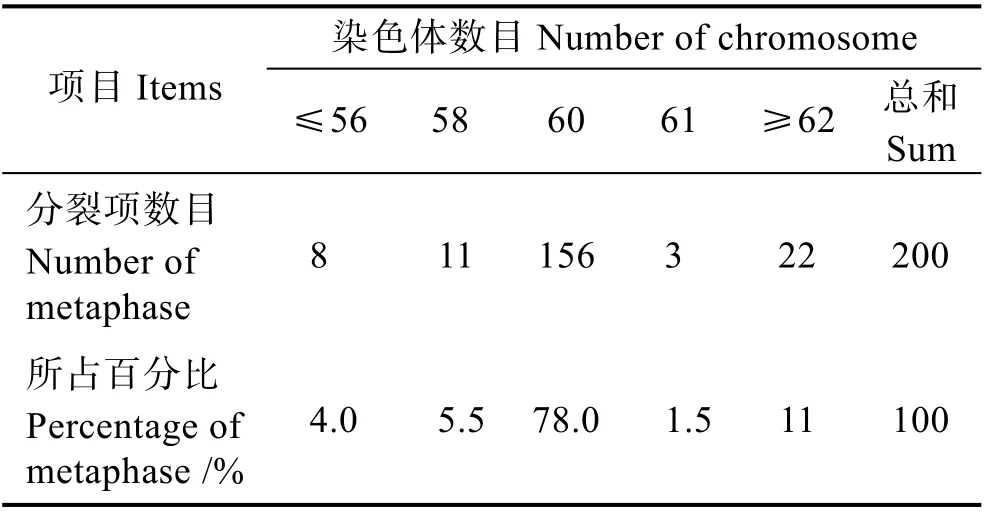
表1 O. sinensis 二倍体染色体数目频率分布Tab.1 Frequency distribution of O. sinensis diploid chromosome number
2.2 染色体相对长度及核型分析
染色体相对长度和臂比统计结果见表2。根据相对长度和臂比,O. sinensis的60 条染色体可配对为30 对(图2B),其中,7 对为中部着丝粒染色体(m),13 对为亚中部着丝粒染色体(sm),6 对为亚端部着丝粒染色体(st),4 对为端部着丝粒染色体(t)。最长和最短的染色体相对长度分别为3.76±0.05 和0.70±0.13,均为亚中部着丝粒染色体。雌雄染色体类型一致,未发现性别相关的异型染色体,也没发现次溢痕和随体。确定本研究O. sinensis的染色体核型为2n=14 m+26 sm+12 st+8 t,臂数(NF)为100。
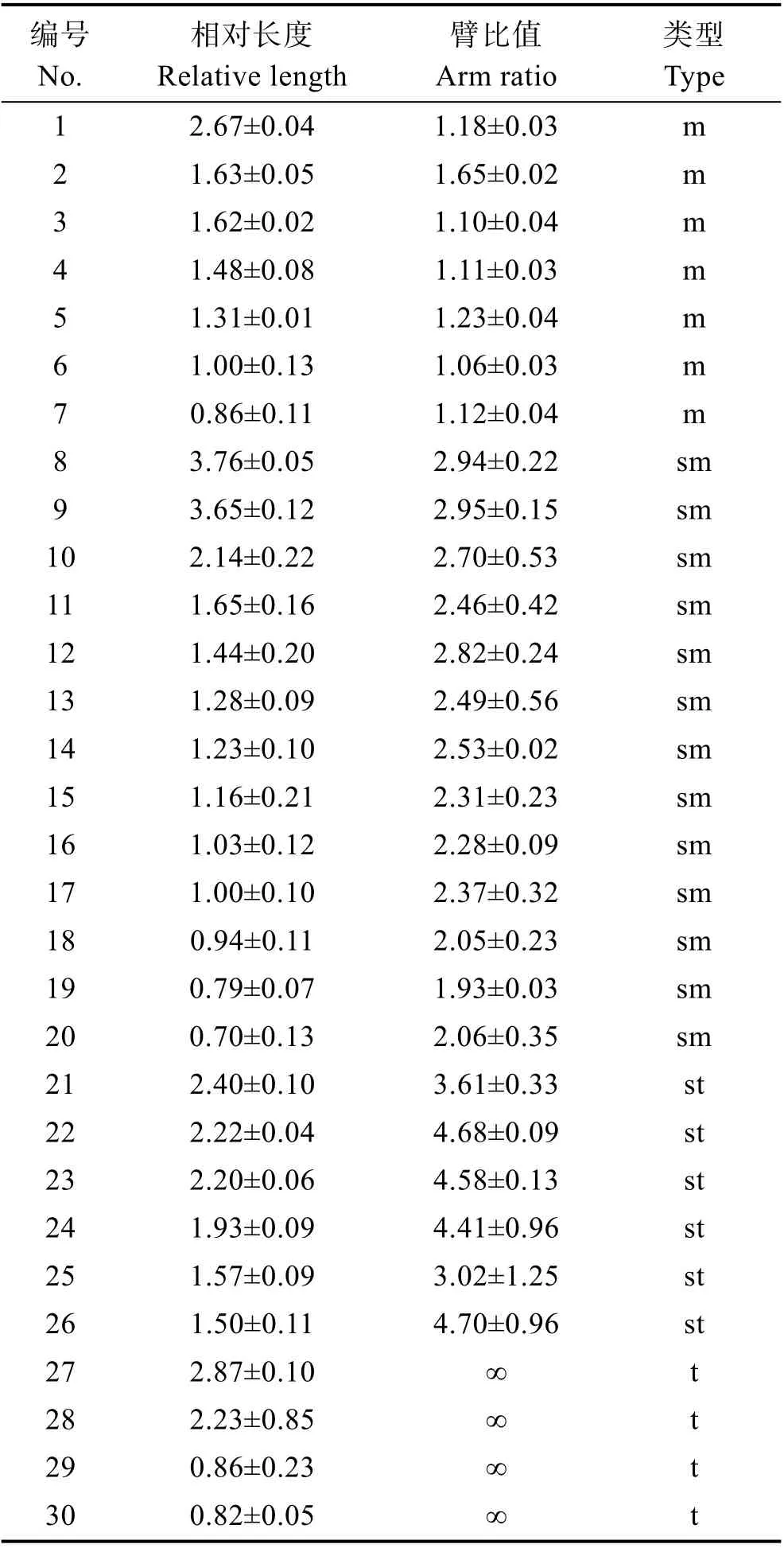
表2 O. sinensis 中期染色体类型、相对长度和臂比值(平均值±标准差)Tab.2 Relative length and arm ratio of metaphase chromosome in O. sinensis (Mean±SD)
3 讨论
现存头足类有700 余种,但仅有7 种报道了染色体核型(表3)。头足类染色体的研究最早见于Inaba(1959)的报道,采用精原细胞和初级精母细胞对长蛸和真蛸的染色体进行制备并分析染色体数目,并未报道核型。染色体标本多采用生物细胞分裂能力较强时期或组织进行制备,像鱼类的头肾、鳃等,细胞分裂指数高,更易获得较多的中期分裂相。已知头足类染色体核型的研究多采用性细胞、受精卵、正在发育的胚胎等(王晓华等, 2011; Papanet al, 2010; Gaoet al,1990; Vitturiet al, 1982; Inaba, 1959)。性腺和受精卵能够获得中期分裂相,但制备过程中细胞极易破裂,容易造成染色体丢失,同时,受精卵因含有较多的卵黄组织,会影响染色体分散及染色(王晓华等, 2011;Vitturiet al, 1982; Inaba, 1959)。正在发育的胚胎细胞分裂旺盛,所制备的染色体标本中期分裂相较多,染色体分散好,轮廓清晰,极适用于头足类染色体标本的制备(王晓华等, 2011)。在胚胎时期,头足类性别尚未分化,难以确定头足类的染色体性别决定机制。本研究利用O. sinensis幼体鳃和肾脏组织,采用活体浸泡秋水仙素暂养的方法,成功获得雌雄个体分散较好、轮廓清晰、收缩适中的染色体中期分裂相(图2A)。首先,O. sinensis幼体处于生长期,细胞分裂相对旺盛,且已出现性别分化,有利于结合染色体核型分析其染色体性别决定机制。其次,采用含秋水仙素海水暂养的方法对活体进行预处理,保证在组织细胞活性下获得实验样本,提高有丝分裂指数。但与胚胎相比,幼体鳃和肾脏有丝分裂指数相对较低,可通过多次重复实验来弥补不足。
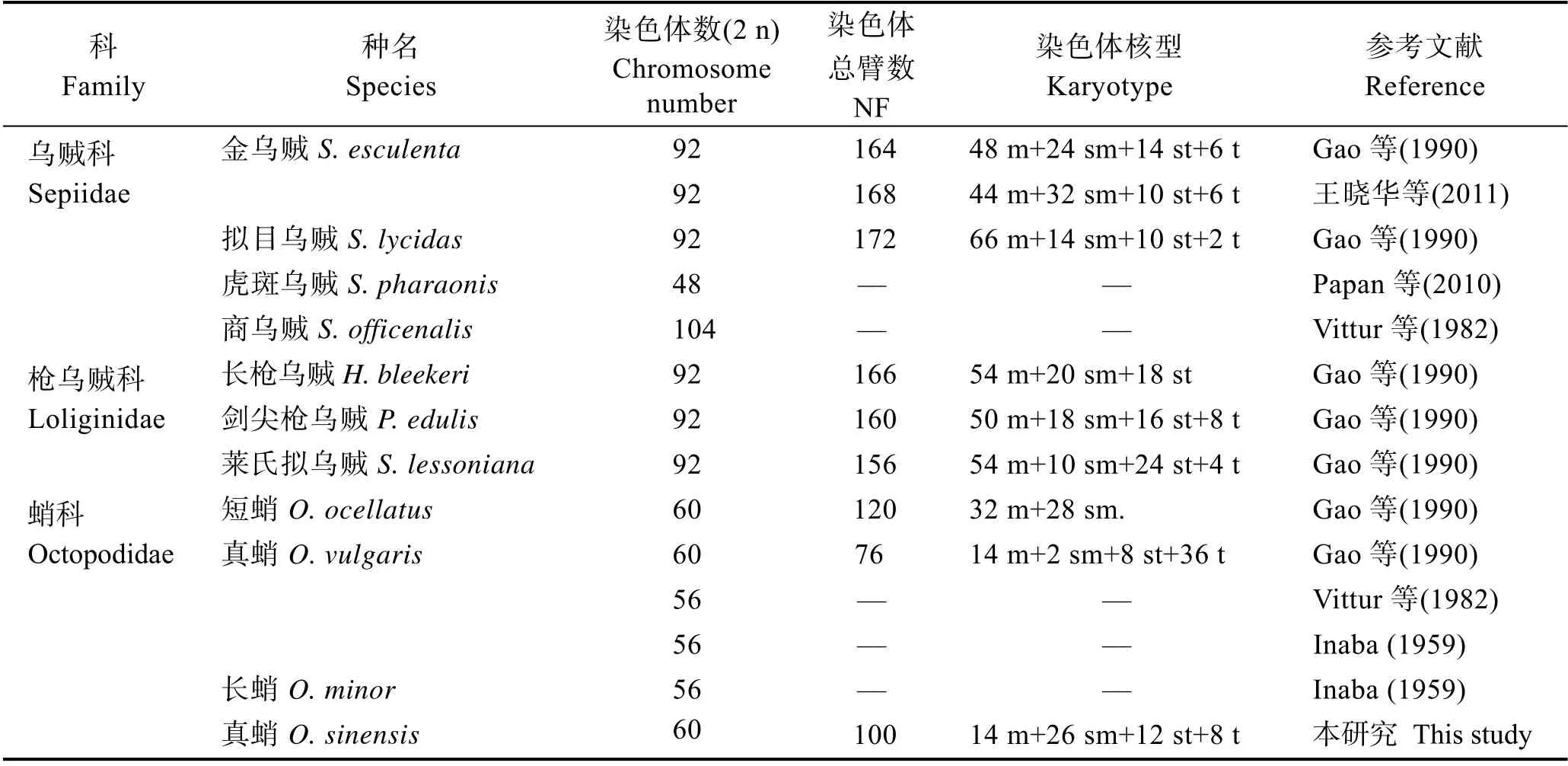
表3 国内外头足类染色体核型研究Tab.3 Study on karyotype of chromosome of cephalopod
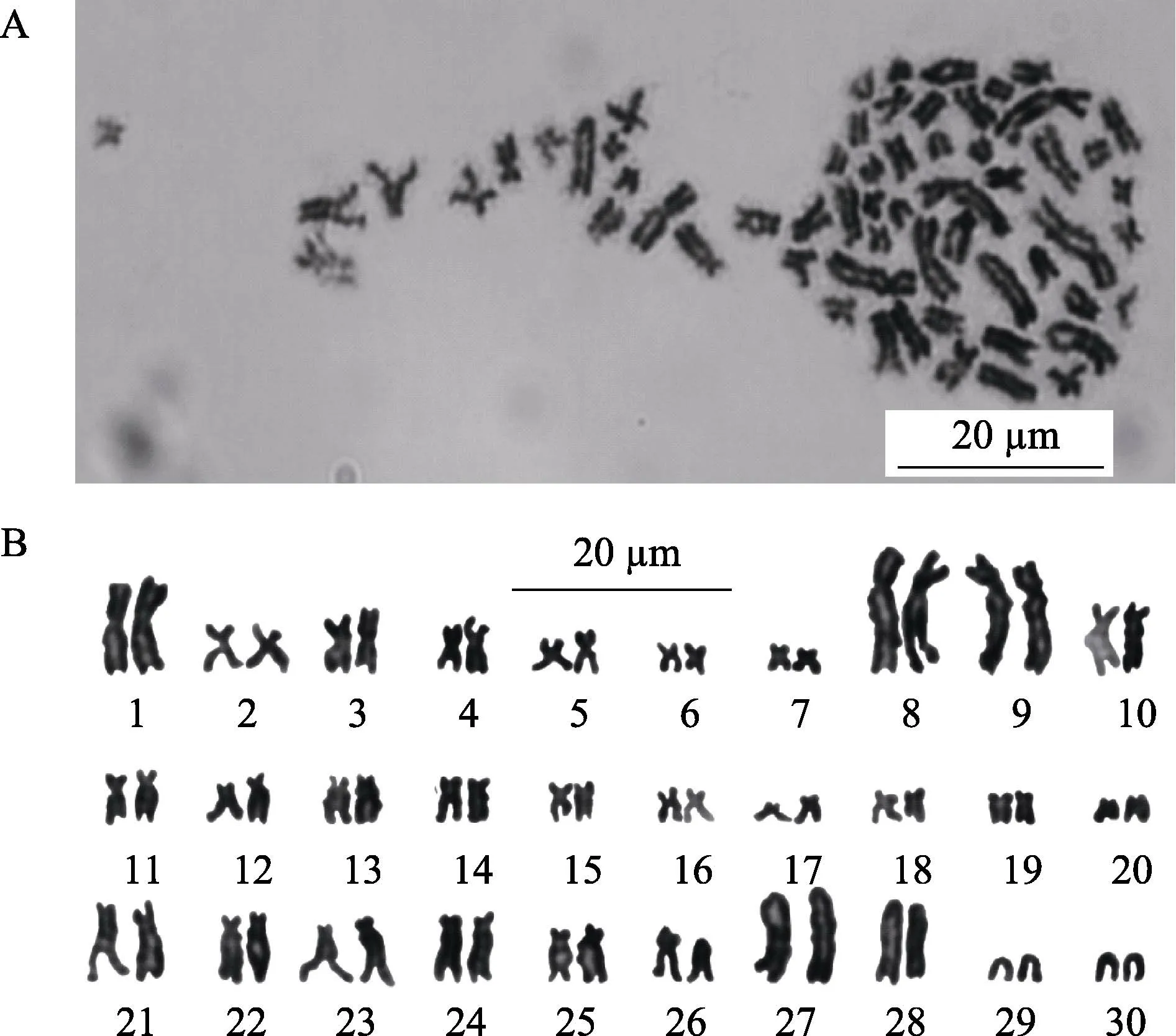
图2 O. sinensis 中期分裂相染色体及核型Fig.2 Metaphase chromosome and karyotype of O. sinensis
两性分化与有性生殖推动了生物的遗传和变异过程。异形染色体(性染色体)对在生物性别决定过程中发挥重要作用。根据异形染色体对在两性中的存在形式,可将目前已知的生物染色体决定模式分为XX/XY 型、ZW/ZZ 型、ZO/ZZ 和XX/XO 型以及复性染色体型(尚晓莉等, 2010)。部分生物的性别决定模式除受到性染色体调控外,常染色体也发挥一定的调控作用,即常染色体性别决定类型(童金苟等,2003)。常染色体性别决定类型是指在现有的技术条件下未能发现异形染色体(尚晓莉等, 2010)。目前,国内外对软体动物染色体性别决定机制的研究较少,国内仅见栉江珧(Atrina pectinata)。周丽青等(2018)利用愈伤组织制备栉江珧染色体标本,分析发现栉江珧的染色体性别决定机制为XX/XY 型。国外研究发现,在腹足类中可能存在XO 或XY 型性染色体(陈宇,2007)。国内外未见头足类染色体存在异形染色体的报道。本研究通过分析O. sinensis雌雄个体染色体核型亦未发现异形染色体,推断头足类染色体性别决定模式可能为常染色体性别决定类型。Twyman(2002)认为,大部分雌雄异体型动物原始性别及第二性征是由性染色体决定的。蛸类外部形态已出现雌雄性别的差异,即第二性征,具体表现为雄性右侧第3 腕足特化为交接腕(图1B、C)。因而,O. sinensis是否存在性染色体有待通过优化实验条件,染色体带型分析等方法进一步探究。
表3 显示,在蛸科中,Gao 等(1990)报道的真蛸(O. vulgaris)二倍体染色体条数为60 条,与Vitturi 等(1982)和Inaba (1959)的研究结果(56 条)存在差异。Gao 等(1990)认为产生该种差异的原因为不同地理群体的生物存在染色体的种内变异。鉴于本研究O.sinensis的染色体条数也为60 条,作者认为产生上述差异的原因更可能为染色体制备方法的差异。研究表明,取样材料对头足类染色体制备的影响较大(王晓华等, 2011)。本研究发现,O. sinensis性腺细胞在染色体制备过程中极易破裂,容易造成染色体的丢失。在乌贼科中,Gao 等(1990)与王晓华等(2011)对金乌贼染色体核型分析所得的染色体条数一致(均为92条),但染色体核型有差异,主要体现在中部着丝粒染色体、亚中部着丝粒染色体和亚端部着丝粒染色体数目上的差异。该差异的产生可能有2 个原因:1) 实验方法不同导致染色体收缩程度不同,进而影响到测量结果;研究显示,秋水仙素浓度过大或处理时间过长均会导致染色体收缩严重,进而影响到测量结果和后期染色体分型(梁述章等, 2019);2) 鱼类中不同地理群体的个体存在染色体多态性现象(Renet al,2014),头足类也可能存在类似现象。
染色体是生物遗传信息的载体,染色体的核型能够反映物种的进化历史和种间亲缘关系的远近(Vitturiet al, 1982)。乌贼科的金乌贼和拟目乌贼与枪乌贼科的长枪乌贼、剑尖枪乌贼和莱氏拟乌贼的二倍体染色体条数均为92 条,而蛸科的O. sinensis、短蛸和真蛸为60 条,说明乌贼科与枪乌贼科的亲缘关系可能更近,而与蛸科的亲缘关系较远。本研究O. sinensis的染色体核型为2n=14 m+26 sm+12 st+8 t,与真蛸(O. vulgaris)(2n=14 m+2 sm+8 st+36 t)相比,O. sinensis的染色体核型主要由中部着丝粒染色体和亚中部着丝粒染色体构成,说明O. sinensis的染色体核型更为稳定(Ahmed, 1976),染色体核型的差异也为二者的区分提供了另一个判定依据。短蛸(O. ocellatus)的染色体核型为2n=32 m+28 sm,不存在亚端部和端部着丝粒染色体,说明在蛸类进化过程中,染色体可能存在易位或倒置现象(Gaoet al, 1990)。从染色体核型特征上可以看出,O. sinensis与真蛸(O. vulgaris)的亲缘关系更近,而与短蛸(O. ocellatus)的较远。头足纲是软体动物门中最高等的类群,其染色体条数(48~104)比其他软体动物(18~70)多(Vitturiet al, 1982;王梅林等, 2000; 孙振兴, 2004),结果支持Patterson(1978)的推断:越原始的软体动物,染色体条数越少。在蛸科、乌贼科和枪乌贼科中,蛸科的染色体条数最少,推断蛸科的进化地位更为原始。
猜你喜欢
云南医药(2021年6期)2022-01-08
川北医学院学报(2021年6期)2021-07-13
军事文摘(2020年14期)2020-12-17
科学之谜(2019年3期)2019-03-28
科学之谜(2018年8期)2018-09-29
中国实用医药(2016年36期)2017-06-20
家教世界·创新阅读(2016年12期)2017-01-09
中学生理科应试(2016年4期)2016-11-19
恋爱婚姻家庭·养生版(2016年9期)2016-09-07
阅读与作文(小学高年级版)(2016年1期)2016-03-04
