
草鱼生长相关优势基因型的聚合效果分析*
2021-08-25 07:03李胜杰杜金星周家辉白俊杰
渔业科学进展 2021年5期
孙 雪 李胜杰 杜金星 姜 鹏 周家辉 白俊杰
(1. 中国水产科学研究院珠江水产研究所 农业农村部热带亚热带水产资源利用与养殖重点实验室广东 广州 510380;2. 上海海洋大学水产与生命学院 上海 201306)
多基因聚合育种是通过不同基因型的个体杂交将有利基因聚合到同一个基因组中,从而获得优良品种的育种方法(Servinet al, 2004)。多基因聚合育种技术在农作物和畜禽中已广泛被应用。利用分子标记辅助选择与传统选育相结合的方法,将小麦的高蛋白基因GPC-B1和抗白粉病基因Pm21、1Dx5和1Dy10等进行聚合,获得兼具高蛋白质含量和高抗病力性状的小麦品系(胡云等, 2016)。在优质肉鸡多基因聚合研究中,将4 个与繁殖性能相关的基因(NPY、NCOA-1、IGF-1和GDF-9)进行聚合,获得在开产日龄、开产体重及300 日龄合格种蛋率最优的优势基因聚合个体,其繁殖性状显著优于上一世代亲本(王钱保等,2018)。中国美利奴羊(Ovis arios)(曾献存等, 2011)和苏姜猪(Sus scrofa)(王宵燕等, 2007)多基因聚合研究结果显示,与生长性状关联的优势基因型聚合数量越多,生长性状表型越优秀。目前,在水产动物中有关基因聚合研究的报道较少,孙效文等(2009)对德国镜鲤(Cyprinus carpio)群体中最大个体及最小个体所富集的优势基因型情况进行分析,发现最大个体中优势基因型数量为1.7 个,最小个体中优势基因型数量为0.7 个,反映了与生长性状相关优势基因型的聚合数量多少与镜鲤生长速度紧密相关。李胜杰等(2019)分析了大口黑鲈(Micropterus salmoides)家系中生长性状相关优势基因聚合效果,发现优势基因聚合数量多少与生长性状显著呈正相关。
草鱼(Ctenopharyngodon idella)是我国淡水养殖鱼类中年产量最高的养殖品种。目前,在草鱼养殖产业中尚未有经过人工选育、通过国家审定的优良养殖品种。近年来,多数草鱼繁殖场对繁殖使用亲本的操作不规范,通常选择亲缘关系近、体型小、性成熟早的个体进行苗种生产,导致养殖草鱼出现一定程度的种质退化现象,制约草鱼养殖产业的健康发展(Jiang,2009; Yuet al, 2014; 田园园等, 2019)。由于草鱼的性成熟年龄普遍为4~5 龄,选育周期长,在草鱼良种选育研究中,有必要探索利用分子标记辅助育种技术来加快草鱼良种选育进度。本实验室前期利用候选基因关联分析方法在草鱼载脂蛋白A-I-1 基因(apoA-I-1)(刘小献等, 2012)、丙酮酸激酶1 型(PKL)(唐小红,2015)、羧肽酶A1 (CPA1)(曹婷婷等, 2012b)、柠檬酸合酶(CS)(樊佳佳等, 2014)、醛缩酶B (Aldo-B)(曹婷婷等, 2012a)、生长催乳素α (SLα)和肌球蛋白重链(MYH)基因上筛选到10 个与生长性状关联SNP 标记,本研究在此基础上通过对亲本的基因型进行分析并构建优势基因型聚合家系,进一步分析生长相关优势基因型的聚合数量与生长性状的相关性,以期为草鱼分子标记辅助育种提供基础资料和实践依据。
1 材料与方法
1.1 草鱼亲本的挑选与养殖
实验鱼来自江苏省苏州市未来水产养殖场,从中挑选48 尾亲鱼,其中,雌鱼和雄鱼各24 尾,放于同一池塘养殖。将每尾亲鱼注射电子芯片标记,同时,剪取腹鳍样本并放入无水乙醇中保存,用于提取DNA 和基因型分析。
1.2 生长性状相关分子标记的选择
本实验室前期所获得的10 个与生长性状相关的SNP 标记(G851A、T+744C、A+762G、C+36A、C-499G、A117C、C+1042A、U8014、U2903 和U11628)分别位于apoA-I-1、PKL、CPA1、CS、Aldo-B、SLα、MYH基因上,经关联分析表明,其与生长性状显著(P<0.05)或极显著(P<0.01)相关。各个标记在进行PCR 反应中所用的引物序列见表1。所有引物由上海捷瑞生物工程有限公司合成。

表1 所用草鱼生长性状相关分子标记的相关信息Tab.1 Primers of growth-associated markers of grass crap
1.3 基因组DNA 的提取
使用TIANGEN 海洋动物组织基因组提取试剂盒(北京天根生化科技有限公司)提取草鱼亲本和子代群体鳍条样品的基因组DNA,使用1.2%的琼脂糖凝胶电泳和酶标仪检测DNA 质量和浓度,DNA 保存于-20℃备用。
1.4 生长相关标记的基因型检测
SNP 标记采用SNaPshot 分型方法,分析其在每尾草鱼中的基因型,委托上海捷瑞生物工程有限公司完成。实验流程:首先根据SNP 标记上下游的序列设计引物,长度为200~500 bp,用于含有SNP 目的片段的扩增,采用多重PCR 方法对模板进行扩增,反应程序采用Touch-down 方法,95℃预变性3 min;94℃变性15 s,60℃退火15 s,72℃延伸30 s,11 个循环,每个循环的退火温度降0.5℃;然后,进入下一个循环程序,94℃变性15 s,54℃退火15 s,72℃延伸30 s,24 个循环;最后,72℃延伸3 min。将扩增到的目的片段用ExoⅠ和FastAP 进行纯化,去除反应产物中的剩余引物和dNTP。使用SNaPshot 试剂盒(ABI 公司, 美国)中的SNaPshot Mix 试剂与纯化后的PCR 产物混合,对PCR 产物进行延伸反应,延伸引物的设计是在SNP 标记的上游或是反向的下游位置,延伸产物温度变性后在ABI3730 全自动测序仪上进行测序。
1.5 家系的构建及子代培育
分析各个与生长性状相关的标记在24 尾雌鱼和24 尾雄鱼亲本中的基因型分布情况,依据家系两亲本中至少有一个杂合子且子代中存在优势基因型的原则,选择可聚合出优势基因型最多的1 对亲本进行繁殖并构建全同胞家系。在繁殖过程中,采用人工注射催产剂的方法促进产卵,收取的受精卵置于环道孵化池内孵化。将孵出的家系鱼苗放入面积为3333 m2的池塘养殖。7 月龄时,从家系子代中采集382 尾实验鱼进行体质量、全长、体高、头长、尾柄高和尾柄长的测量,同时,剪取腹鳍样本并放入无水乙醇中保存,用于提取DNA 和基因型分析。
1.6 数据统计分析
先对家系子代个体中的10 个生长标记进行基因分型,再进行优势基因型的统计分析。根据所含优势基因型数量不同进行分组,采用SPSS 19.0 软件的一般线性模型(general linear model, GLM),对不同优势基因数量的组别与生长性状之间相关性进行最小二乘分析。统计分析模型采用如下公式:
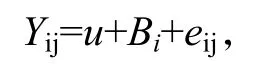
式中,Yij为某个性状第i个标记第j个个体观测值;u为实验观测所有个体的平均值(总体平均值);Bi为第i个标记的效应值;eij为对应于观察值的随机残差效应(徐磊等, 2014)。
2 结果与分析
2.1 草鱼亲本基因型分析
分析48 尾草鱼亲本的基因型,从中筛选出能够在子代中聚合出优势基因型数量最多(7 个)的1 对父母本个体配对繁殖及构建家系,亲本基因型结果见表2。在家系子代中,G851A 标记的基因型均为纯合子,C+36A 和U11628 标记都不存在优势基因型,在下面的优势基因型数量统计中不分析这个3 个标记,仅统计其余7 个标记。部分标记的基因型检测结果见图1。
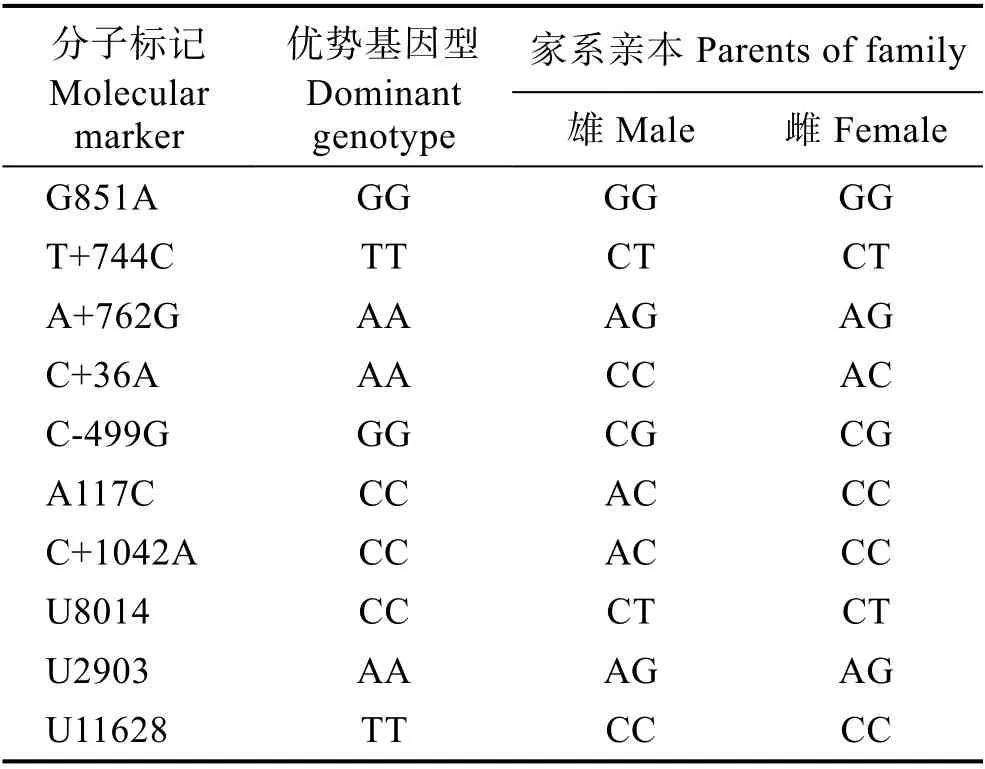
表2 10 个生长标记在草鱼家系亲本中的基因型Tab.2 Genotype of 14 growth related markers in grass crap parents
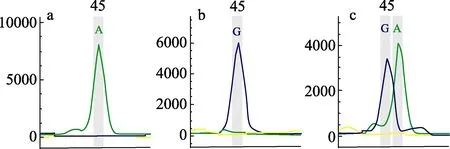
图1 A+762G 标记基因型检测图Fig.1 A+762G marker genotype detection map
2.2 草鱼子代基因型分析
对T+744C、A+762G、C-499G、A117C、C+1042A、U8014 和U2903 标记在草鱼家系子代中的优势基因型进行统计分析,每个标记的优势基因型分布频率见表3。结果显示,C+1042A 标记的优势基因型分布频率最高,在子代中所占比例为51.31%;U8014 标记在子代中的优势基因型频率最低,所占比例为24.35%。不同子代个体中含有的优势基因型数量范围为0~7 个,其中,具有聚合优势基因型数量7 个的个体数量是6 尾,而不含优势基因型数量的个体数量为44 尾。所占比例最多的为具有3 个优势基因型的个体,占子代总数的21.99%。全部子代个体中平均优势基因型的数量为2.58,相比亲本群体的优势基因型平均数量(1.00)得到显著提高(P<0.05)。

表3 不同标记中优势基因型的分布频率Tab.3 The frequency of dominant genotypes of different markers
2.3 草鱼生长性状相关的优势基因聚合效果分析
对草鱼子代中含有的优势基因数量及其生长性状进行分析。结果显示,具有7 个优势基因型组的生长性状最佳,体质量平均值为176.67 g,比不含优势基因型组的平均体质量高36.26%。不含优势基因型的个体的体质量、体长、体高、头长、尾柄长和尾柄高等5 个生长性状平均值均低于其他具有多个优势基因型的个体,且7 个、6 个、5 个、4 个、3 个和2 个优势基因型组均分别与1 个和0 个优势基因型组的平均体质量差异显著(P<0.05)。7 个优势基因型组与5 个、4 个、3 个和2 个优势基因型组的平均体质量差异显著(P<0.05),6 个优势基因型组与3 个和2 个优势基因型组的平均体质量差异显著(P<0.05)。
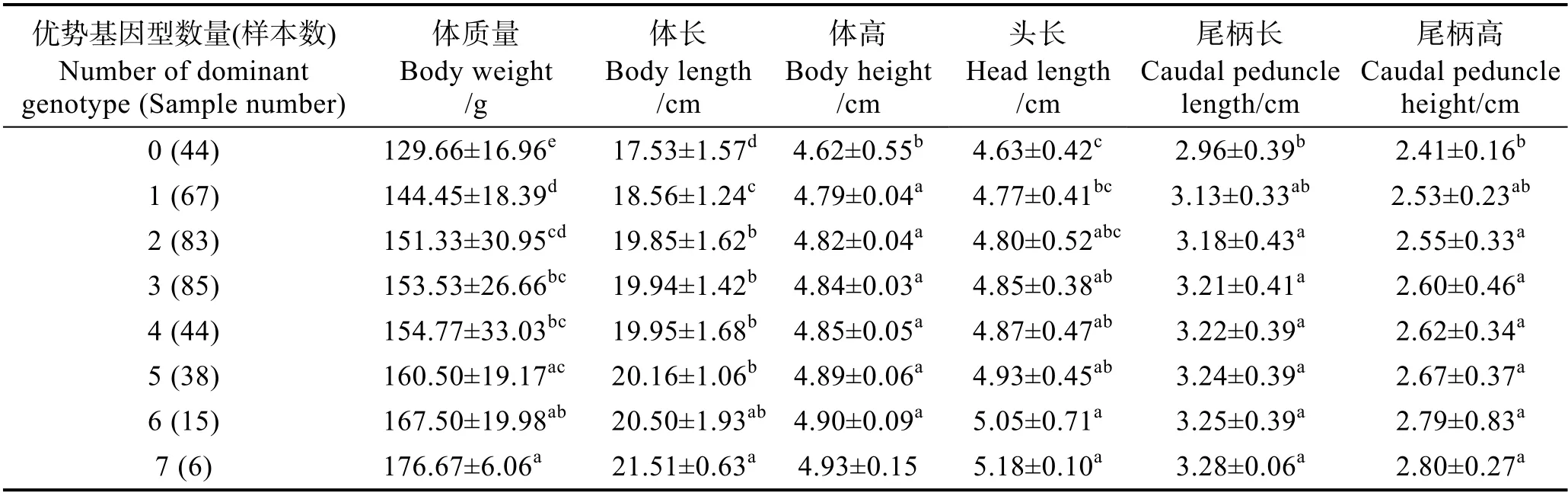
表4 家系子代中所含不同优势基因型数量的个体的生长数据比较(平均值±标准差)Tab.4 Analysis of growth traits of grass carp with different number of pyramiding dominant genotypes (Mean±SD)
3 讨论
传统选育方法主要通过对表型性状选择来进行选育改良,选择效果易受环境等因素影响。分子标记选择可克服传统育种技术的一些不足,提高育种效率和选择强度(刘福平等, 2008)。分子标记分为连锁标记和功能标记,应用连锁标记经常达不到育种目的,且连锁标记可能受遗传背景的限制,无法应用于非多态群体(孙立亭等, 2019; Fujiiet al, 2000)。功能标记是指从影响性状变异基因的功能域开发出来的多态性标记,利用功能性标记开展分子标记辅助选择,对基因本身进行选择,从而保证了选择的准确性和高效性(李扬等, 2016; 陈华增等, 2010; 顾冰宁等, 2018)。本研究基于实验室前期利用候选功能基因关联分析研究获得的与草鱼生长性状相关10 个SNP 标记作为研究对象,选择可聚合出优势基因型最多的1 对亲本构建家系。结果显示,子代个体的优势基因型的平均数量为2.58,显著高于亲本群体的平均优势基因型数量(1.00),说明通过目的性挑选亲鱼进行基因聚合育种研究,能够提高选择群体中优势基因型的数量,为后续利用分子标记辅助育种技术开展草鱼良种选育提供了科学依据。
生长是决定鱼类经济价值最重要的性状之一,受众多功能基因调控(李胜杰等, 2018)。如果仅仅从单基因入手进行分子辅助育种,往往不能达到全面提高生长性状的目的。多基因聚合技术可实现多个优势基因的有利整合,达到培育出生长性状优良的草鱼品系(傅建军等, 2016)。宋易等(2016)在翘嘴鳜(Siniperca chuatsi)“华康1 号”5 代选育中发现,F1~F5所含优势基因型平均数量依次增加,且生长速率也随之增加。徐磊等(2014)分析大口黑鲈优势基因型聚合效果发现,子代个体中含优势基因型数量与生长性状显著相关,且含6 个优势基因型的个体平均体质量比含1 个优势基因型的个体提高34.14%。李红霞等(2014)在建鲤(Cyprinus carpiovar.Jian)优势基因聚合中发现,子代含4 个优势基因型的个体平均增重比不含优势基因型的个体快14%。本研究子代个体所含优势基因型数量与生长性状的相关性分析结果显示,优势基因型聚合数量与草鱼生长性状呈正相关,与鳜鱼(宋易等,2016)、大口黑鲈(李胜杰等, 2019)、鲤(孙效文等, 2009)的研究结果一致。含7 个优势基因型个体的平均体质量比不含优势基因型的个体提高36.26%。可见,利用多基因聚合技术将草鱼生长相关的基因聚合,可实现生长表型性状的大幅度改良。这也表明,在今后的草鱼良种选育过程中,利用分子标记技术直接选择可聚合草鱼生长优势基因数量多的个体作为亲本进行繁育,可取得良好的遗传改良效果。
多基因聚合过程中,每增加一个基因的筛选,将会有一部分材料遭到淘汰,可供挑选的目标个体数也将大大降低(徐磊等, 2014)。本研究也反映出这种现象,从子代个体所含的优势基因型的数量分布结果来看,含2 个和3 个优势基因型个体数最多,所占比例分别为21.73%和22.25%;含7 个优势基因型的个体数最少,所占比例为1.57%,可供选择的数量相对较少。孙立亭等(2019)对水稻稻瘟病抗性进行多基因聚合效果分析时发现,随着含抗病基因数的增加,对应的材料数量明显降低。这可能是由于个体在含2~3 个优势基因型时,其基因型的组成方式存在多样化,随着子代含有优势基因型数目的增加,基因型之间的组合方式随之减少。后续可通过扩大研究群体或者增加聚合基因的数量来解决这一问题,从而获得较多数量的目标群体。由于受客观条件因素的影响,本研究未能探讨多基因聚合育种技术应用于选育中的效果,尚需进一步研究。
多基因聚合效果分析所用的7 个与体质量显著相关的标记均为本实验前期通过候选基因关联分析方法筛选而获得的,前期研究结果显示,这7 个标记的优势基因型个体的平均体质量比其他基因型个体的平均体质量高6.86%~11.00% (曹婷婷等, 2012a、b;樊佳佳等, 2014; 唐小红, 2015),其中,A+762G 标记对体质量影响作用最大,其优势基因型个体的平均体质量比非优势基因型个体高11.00% (唐小红, 2015)。在本研究的家系子代中,这7 个标记的优势基因型个体的平均体质量比非优势基因型个体提高了4.27%~12.11%,其中,A+762G 标记的优势基因型个体与非优势基因型个体的平均体质量差异最大,优势基因型个体的平均体质量为162.41 g,比其余个体的平均体质量提高了12.11%,本研究结果与前期的研究结果基本一致,均反映出A+762G 标记对体质量的影响作用最大。A+762G 标记位于丙酮酸激酶1 型(PKL)基因上,PKL是糖酵解过程中最关键的催化酶,可催化PEP 转化为丙酮酸,将高能磷酸键转移给ADP生成ATP,从而参与机体生命活动及生长发育过程(戴超等, 2014)。在草鱼基因聚合育种实践中可考虑将PKL基因上A+762G 标记作为首选目标标记。
本研究不同基因的组合效应分析发现,生长相关的优势基因型聚合数量越多,其生长性状表型值越高,但具有4 个和3 个优势基因型的个体平均体质量相差很小(1.24 g),具有3 个和2 个优势基因型的个体平均体质量相差也很小(2.2 g)。优质肉鸡繁殖性能相关标记的聚合基因型的效应分析研究中也存在类似结果(张增荣等, 2013)。推测可能多基因聚合并不是单基因的基因型效应的简单相加,而是不同基因之间存在一定的相互作用,从而影响整个性状的形成(李国辉等, 2010)。李红霞等(2014)分析建鲤ODC1基因与增重相关性发现,其主效SNP 标记之间也存在拮抗或协同作用。李胜杰等(2019)研究大口黑鲈生长相关标记的聚合效果也发现,基因之间存在互作效应,进而影响其生长性状。赵秀华(2012)对京海黄鸡GFBP-1、IGFBP-2和STAT5基因进行三基因互作效应分析,结果显示,三基因聚合效应>二基因聚合效应>单个基因效应,但组合基因效应并非是单个基因型效应的简单累加。上述研究提示,今后的多基因聚合育种要将单基因效应分析及不同基因的组合效应分析相结合,选择最优秀和合理的多基因聚合式应用于生产实践中。
猜你喜欢
中国生育健康杂志(2022年4期)2022-11-25
传染病信息(2022年4期)2022-11-23
当代水产(2022年7期)2022-09-20
当代水产(2022年5期)2022-06-05
当代水产(2022年3期)2022-04-26
承德医学院学报(2021年1期)2021-02-25
广东第二课堂·小学(2018年9期)2018-10-24
生物学教学(2018年8期)2018-09-03
安徽农学通报(2017年9期)2017-05-19
中国医药导报(2017年6期)2017-04-06
