
不同昆虫取食对烟草信号分子和防御酶的影响
2021-08-13 06:06:02余源婵杨茂发商胜华刘健锋于晓飞
环境昆虫学报 2021年4期
余源婵,杨茂发, *,商胜华,刘健锋,于晓飞
(1.贵州大学昆虫研究所/贵州山地农业病虫害重点实验室,贵阳 550025;2. 贵州大学烟草学院,贵阳 550025;3. 贵州省烟草科学研究院,贵阳 550081)
烟草NicotianatabacumL.是我国重要的农业经济作物,随着烟草的规模化种植,烟草病虫害呈逐年上升趋势(曾维爱等, 2017),其中烟蚜MyzuspersicaeSulzer、斜纹夜蛾SpodopteralituraFabricius和棉铃虫HelicoverpaarmigeraHübner是烟草种植中常见的3种主要害虫,分属刺吸性和食叶性害虫,具有分布范围广、为害周期长等特点,严重危害烟草生产(郭线茹等, 2006)。目前关于烟草虫害防治方法较多,然因化学防控具有经济、快速和使用方便等优势仍在多数地区得到广泛应用(彭孟祥等, 2019),严重威胁到烟草产品质量安全(李江舟等, 2018)。目前已知烟蚜、斜纹夜蛾及棉铃虫已对多种常用杀虫剂产生了不同水平的抗性(刘佳等, 2016; 胡红岩等, 2018; 孟建玉等, 2018)。因此,通过探究虫害诱导的植物防御机制以寻求合适的方法使植物产生抗虫防御警备,可大大减少杀虫剂的使用,已成为当前害虫综合防治的研究热点(王杰等, 2018)。
植食性昆虫取食植物时,其口腔分泌物内的激发子可被植物识别从而激活植物体内一系列信号转导途径(Schuman and Baldwin, 2016)。常见植食性昆虫口器有咀嚼式和刺吸式两类,前者取食主要诱导激活植物茉莉酸(jasmonic acid, JA)途径,使得植物体内JA及其化合物含量发生变化(顾小辉等, 2017);后者则通过刺吸植物韧皮部汁液获取植物营养,同时传播植物病毒,主要激活植物体内水杨酸(salicylic acid, SA)途径(Leitneretal., 2005)。各种防御信号途径之间可相互影响,交叉反应,使植物产生多种抗性机制(彭金英和黄勇平, 2005)。茉莉酸、一氧化氮(nitric oxide, NO)和硫化氢(hydrogen sulfide, H2S)是植物体内普遍存在的信号分子,参与植物体内多种生理活动,其中JA除参与调节植物的生长发育外,还在昆虫取食和胁迫反应中起到信号传递作用(蒲恒浒等, 2019),可诱导增加植物多酚氧化酶(polyphenol oxidase, PPO)、脂氧合酶(lipoxygenase, LOX)、过氧化物酶(peroxidase, POD)及SA信号转导途径的关键限速酶苯丙氨酸解氨酶(phenylalanine ammonia lyase, PAL)等多种防御酶的合成直接抵御虫害(Koramutlaetal., 2014; Kanchiswamyetal., 2015)。NO作为一种重要的信号分子,不仅参与植物的种子休眠和萌发以及根的形态建成等生长发育过程,还参与调节植物细胞的气孔运动以及增加植物抗性(Hasanuzzamanetal., 2018),其中NO依赖SA的产生激活或强化植物防御反应,SA通过改变顺乌头酸酶(aconitase,ACO)、过氧化氢酶(catalase,CAT)和POD活性进而参与NO合成途径(彭金英和黄勇平, 2005; 张艳敏, 2017)。有报道发现烟蚜和斜纹夜蛾取食均可诱导烟叶NO含量升高(张艳敏等, 2017)。H2S信号分子具有与NO类似的功能,根据两者各自浓度在植物中作为信号或损伤促进剂起协同或拮抗作用(Corpasetal., 2019)。研究证实NO和H2S水平的增加及其相互作用,可增加烟草对极端盐度条件胁迫的耐受性(Da Silvaetal., 2017)。此外,H2S作为JA信号的下游,调控拟南芥Arabidopsisthaliana子叶的气孔发育(Dengetal., 2020)。以上研究均证实JA、NO和H2S等防御信号分子参与了植物抗性的诱导。
植物体通过多种信号途径间通讯,激活各种防御基因的表达并合成积累相关防御物质,如PAL、PPO、POD和LOX等多种防御酶蛋白直接抵御虫害(林丹, 2018)。植物体内的防御蛋白酶经昆虫摄入后,可影响害虫肠道内胰蛋白酶、胰凝乳蛋白酶等消化酶活性,扰乱昆虫原有的肠道环境极其消化吸收功能,造成虫体发育减缓或死亡(桂连友等, 2005)。本文以棉铃虫、斜纹夜蛾及烟蚜等3种昆虫取食后检测烟草叶片内的防御生理指标变化来探讨不同昆虫取食诱导的烟草抗虫生理机制,旨在为形成烟草虫害绿色防控综合技术体系,替代或减少化学农药的使用次数与用量提供理论依据和技术参考。
1 材料与方法
1.1 试验材料与试验设计
试验于2019年6-9月在贵州大学昆虫研究所的实验用地及实验室内完成,实验选用贵州烟区生产上常用的烟草品种“雄性不育K326(male sterility K326, MS K326)”为材料,于实验棚育苗池内进行漂浮育苗,待盘内幼苗长至4片真叶时,用腐殖土进行单株盆栽培育。将盆栽烟苗移入孔径为160目的防虫网笼中,自然光照,定期浇灌肥料水,待烟苗长至6片真叶后,选取长势一致的健康烟苗供试(Zong and Wang, 2007)。
试验用烟蚜M.persicae无翅成蚜为实验棚内烟株上笼罩繁殖的自然种群,采自贵州省遵义市进化镇烟田,采用李林森法传接蚜虫(李林森, 1983),烟蚜饥饿3 h后供试。斜纹夜蛾种群为实验室长期建立的稳定实验种群,用人工饲料连续饲养多代,饲料配方参照陈其津等(2000),略作修改,饲养条件为温度27±1℃,相对湿度70%±5%,光周期14 L∶10 D;棉铃虫种群购于科云生物,饲养条件为温度27±1℃,相对湿度70%±5%,光周期16 L∶8 D。用新鲜烟叶喂食斜纹夜蛾和棉铃虫初孵幼虫,待虫体长至3~4龄时选取大小一致的虫体单头进行饥饿处理12 h后供试。
在6片真叶期,选取长势一致的烟苗用于试验。试验设置6种处理:(1)烟蚜取食,接15头饥饿3 h后的无翅成年蚜虫至烟苗除心叶外的从上至下数第1片完全展开叶背面(张艳敏等, 2017);(2)斜纹夜蛾和棉铃虫取食,分别接1头蜕皮12 h内的斜纹夜蛾/棉铃虫3龄幼虫至烟苗从上至下数第1片完全展开叶,用纱袋笼罩处理叶,防止虫体转移取食(Zong and Wang, 2007);(3)针刺,用PUL-1000程控水平控制仪拉取的毛细血管细针即显微注射针5根一束,对供试烟苗从上至下数第1片完全展开叶进行针刺处理,针刺3次为一组,分别在T5 min、T10 min时进行针刺,之后开始正式处理计时(Gossetetal., 2009);(4)打孔,用自制打孔器(d=0.3 cm)在烟苗除心叶外的从上至下数第1片完全展开叶上打孔,1次/h,打孔3 h模拟虫体持续取食(打孔总面积约为斜纹夜蛾、棉铃虫组取食面积);(5)以同期未受为害的健康烟苗相同位置叶片作为对照(CK)。将相同处理烟株放入无虫网笼中,保证不同处理间直线距离不少于10 m,所有处理烟苗置于相同实验用地环境。处理6、12、24、48和72 h后摘取各处理烟苗从上至下数第2片完全展开叶组织放入提前准备好的自封袋中封口后立即放入冰盒,转移至-80℃冰箱中保存待测。以上处理重复3次,每次重复各处理烟苗5株。
1.2 样品处理、生化指标测定与计算方法
1.2.1样品处理
JA含量测定:将叶片叶肉组织加液氮研磨后分别称取0.1 g加入1 mL预冷PBS缓冲液(购于生工;0.01 M, pH 7.4),快速震荡摇匀后对匀浆液进行超声破碎,按照Plant JA ELISA KIT进口试剂盒(购于上海晶抗生物公司)说明书要求条件离心(常温条件下5 000 r/min离心5~10 min),取上清检测。NO、H2S、PAL、PPO、LOX和POD等生理指标测定:将叶片叶肉组织加液氮研磨后分别称取 0.1 g加入1 mL各测定指标提取液,快速震荡摇匀后按照一氧化氮(NO)、硫化氢(H2S)和苯丙氨酸解氨酶(PAL)、脂氧合酶(LOX)、多酚氧化酶(PPO)、过氧化物酶(POD)各微量法测试盒(购于苏州科铭生物技术有限公司)说明书要求进行低温离心(NO和H2S粗样品分别于4℃、10 000 r/min下离心15 min和10 min;PAL粗样品于4℃、10 000 r/min下离心10 min,LOX粗样品于4℃、16 000 r/min下离心20 min,PPO和POD粗样品于4℃、8 000 r/min下离心10 min),然后取上清置于冰上待测。
1.2.2生化指标测定与计算方法
JA测定采用酶联免疫分析法(张兴丽, 2013),按照Plant JA ELISA KIT进口试剂盒说明书进行操作,测定产物特定波长的吸光值(JA: 450 nm),代入测定已知浓度(y)标准品吸光度值(x)绘制的直线回归方程(y=1 427.4x-190.09,R2=0.9964)中,计算出样品浓度(pmol/L FW)。NO、H2S两种信号分子含量和PAL、LOX、PPO、POD等酶活性测定按照对应的各试剂盒说明书进行操作,测定产物特定波长的吸光值(NO: 550 nm; H2S: 665 nm; PAL: 290 nm; LOX: 280 nm; PPO: 525 nm; POD: 470 nm),最后根据说明书提供的公式:NO含量(μmol/g FW)=0.25×(ΔA+0.0103)÷W、H2S含量(nmol/g FW)=681.6×ΔA÷W;PAL(U/g FW)=26.6×ΔA÷W、LOX(U/g FW)=33.33×ΔA÷W、PPO(U/g FW)=120×ΔA÷W(注前面公式中ΔA=A测定-A对照,W为样品质量);POD(U/g FW)=4 000×ΔA÷W(注ΔA=A2 min-A1 min,W为样品质量),计算样品中对应的各信号分子含量和防御酶活性(温娟等, 2017)。每样品重复检测2次。
1.3 数据处理
采用Microsoft Excel 2010软件对实验数据整理,利用SPSS 18.0软件对数据进行单因素方差分析(One-way ANOVA)检验不同处理间的差异显著性,利用GraphPad Prism 7.0对数据进行绘图。
2 结果与分析
2.1 不同昆虫取食对烟草‘MS K326’叶片中信号分子含量的影响
2.1.1不同昆虫取食对烟草‘MS K326’叶片中JA含量的影响
棉铃虫取食条件下,烟叶内JA含量呈升高降低趋势,48 h时含量显著升高达到峰值,较对照含量上升了10.55%,之后有所降低;斜纹夜蛾取食下,JA在12 h时含量显著低于对照和打孔含量,之后回升接近对照。烟蚜取食下,烟叶内JA含量升高总体受到抑制,24 h时抑制显著,含量较对照含量降低了10.38%,针刺JA含量则逐渐升高,直到48 h时含量回升与对照含量接近。说明不同昆虫取食诱导烟叶内JA含量变化存在差异,棉铃虫取食明显诱导JA含量升高,斜纹夜蛾取食抑制JA含量升高,之后含量回升接近对照,烟蚜取食则抑制了烟叶内JA含量升高(表1)。
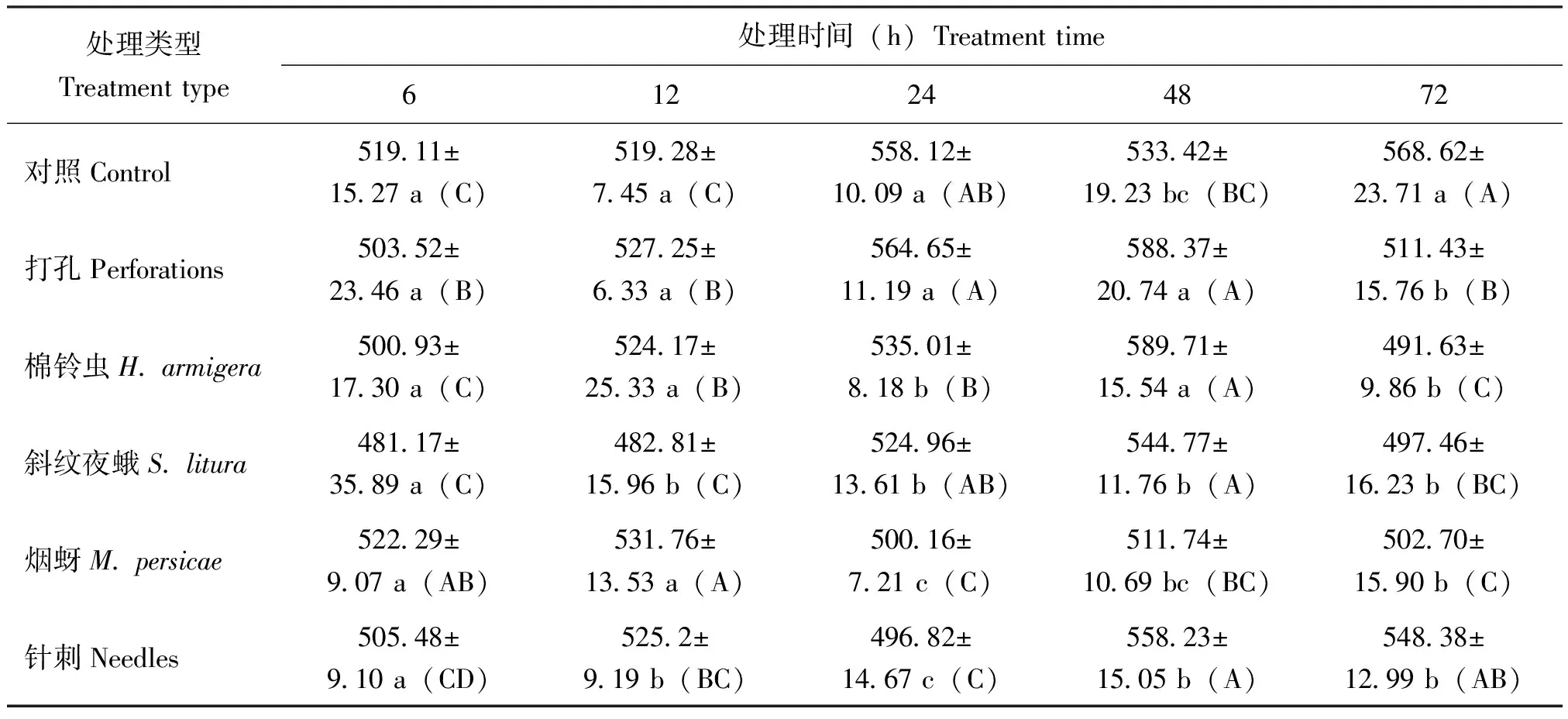
表1 不同昆虫取食对烟草‘MS K326’叶片中信号分子JA含量的影响(单位:pmol/L)
2.1.2不同昆虫取食对烟草‘MS K326’叶片中NO含量的影响
棉铃虫取食后烟叶内NO含量呈逐渐升高后持平趋势,取食24 h时,NO含量升至峰值,较对照升高1.48倍;斜纹夜蛾取食烟草后烟叶内NO含量升高整体受到一定抑制,处理12 h时含量显著低于对照,较同时段对照含量降低了33.33%,处理72 h时NO含量回升至与对照接近,打孔NO含量在处理48 h时达到含量峰值,以上3种处理中的NO峰值出现前后顺序为:棉铃虫>打孔>斜纹夜蛾。烟蚜取食后NO含量在6 h时明显受到抑制,12 h时含量升高与对照含量接近,之后呈逐渐下降趋势,而针刺NO含量在前两个时段中出现一定下降趋势,之后逐渐升高。实验表明,棉铃虫取食诱导NO升高明显,斜纹夜蛾取食下NO含量受到一定抑制后升高不明显,烟蚜则是抑制烟叶内NO含量升高(表2)。
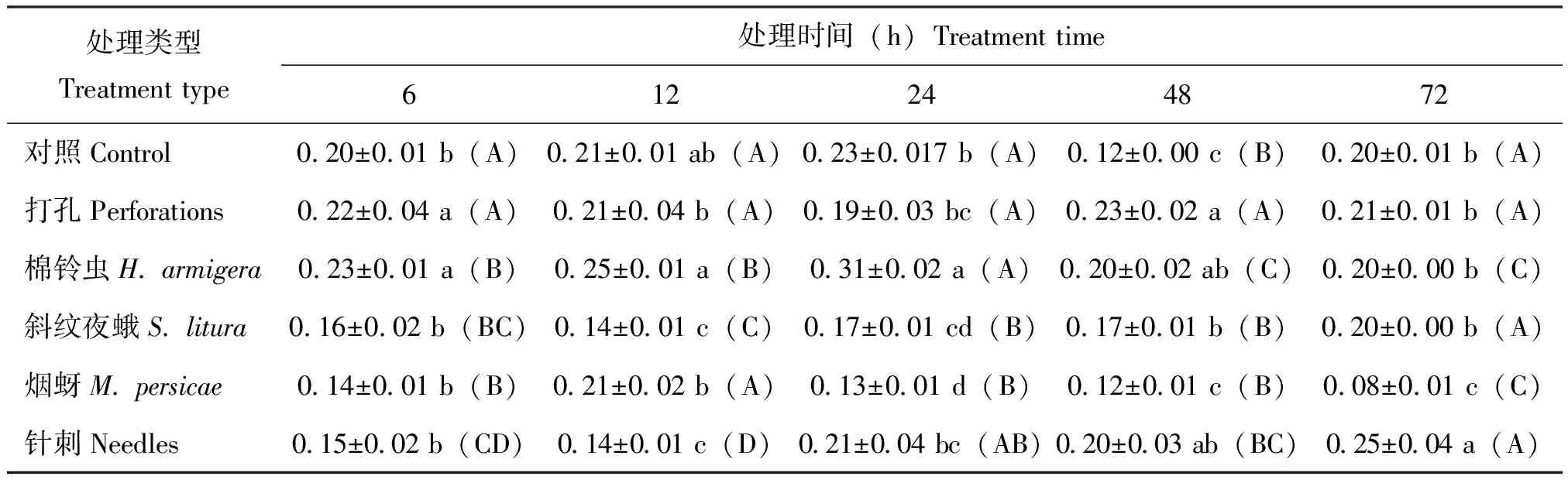
表2 不同昆虫取食对烟草‘MS K326’叶片中信号分子NO含量的影响(单位:μmol/g)
2.1.3不同昆虫取食对烟草‘MS K326’叶片中H2S含量的影响
不同昆虫取食烟草‘MS K326’均可诱导烟草内H2S含量发生显著变化(表3)。其中,斜纹夜蛾幼虫取食6 h时烟草内H2S含量与对照差异显著,含量高达对照含量的1.74倍,之后烟叶内H2S含量均呈一定下降趋势,而棉铃虫取食则是呈持续性下降趋势,6 h时达到含量峰值较对照显著升高。打孔处理下H2S含量变化呈缓慢升高后降低趋势,其峰值低于斜纹夜蛾和棉铃虫取食处理组。烟蚜取食下,H2S含量呈逐渐升高后降低趋势,在处理48 h时含量最高,为对照含量的1.77倍,针刺含量仅在处理6 h时显著高于对照。上述结果表明棉铃虫和斜纹夜蛾撕咬取食较烟蚜刺吸取食能较快诱导烟叶内H2S含量升高。
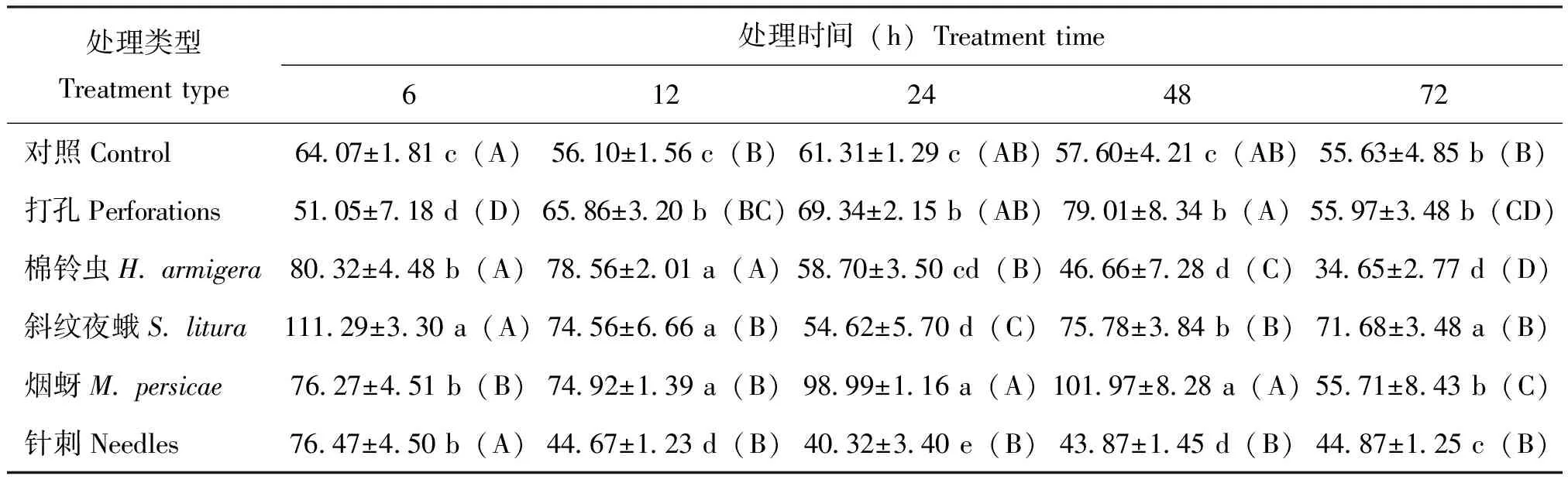
表3 不同昆虫取食对烟草‘MS K326’叶片中信号分子H2S含量的影响(单位:nmol/g)
2.2 不同昆虫取食对烟草‘MS K326’叶片中防御酶的影响
2.2.1不同昆虫取食对烟草‘MS K326’叶片中PAL活性的影响
棉铃虫和斜纹夜蛾取食后叶片内PAL活性均在72 h时达到峰值,棉铃虫取食下PAL活性开始呈持续上升趋势,48 h时活性骤减,之后迅速升高,除48 h外其余各时段下PAL活性均显著高于对照;斜纹夜蛾取食6 h时烟叶内PAL活性与较对照明显受到抑制,12 h时活性有所升高,24 h时活性受到抑制,之后处理时段下逐渐升高;无论棉铃虫还是斜纹夜蛾取食后,叶片内PAL活性升幅均高于对照和打孔处理组,斜纹夜蛾诱导的PAL活性峰值高于棉铃虫。烟蚜取食下烟叶内PAL活性在12 h时明显受到抑制,之后缓慢升高,48 h明显升高后活性与棉铃虫取食株接近,之后降低,其活性峰值高于对照和针刺。表明棉铃虫和斜纹夜蛾取食诱导的PAL活性较烟蚜高,打孔的PAL活性高于针刺组,烟蚜处理较针刺组明显诱导PAL活性升高,由此可推测昆虫唾液分子和较大的机械损伤创口更易诱导烟叶内PAL活性升高(图1)。
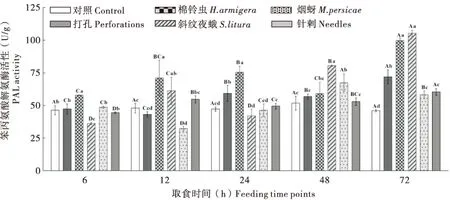
图1 不同昆虫取食对烟草‘MS K326’叶片中防御酶PAL活性的影响Fig.1 Dynamics of the PAL activity of tobacco ‘MS K326’ leaves infested by different insects at different feeding time points注:图中各数据误差线来源于SD。不同柱状图上的英文小写字母不同表示在同一时间不同处理下在0.05水平上差异显著,相同柱状图上的英文大写字母不同表示相同处理不同时间段下在0.05水平上差异显著。下同。Note: Error lines in the figure were from SD. Different lowercase letters on different bar charts indicated significant differences at the 0.05 level under different treatments at the same time, different uppercase letters on the same histogram indicated significant differences at the level of 0.05 at different time periods of the same treatment. The same below.
2.2.2不同昆虫取食对烟草‘MS K326’叶片中LOX活性的影响
棉铃虫处理24 h后,烟叶内LOX活性升至峰值(60.64 U/g)与对照接近,之后活性升高受到抑制;斜纹夜蛾取食后烟叶内LOX活性呈现波动性降低升高,12 h时LOX活性明显受到抑制,72 h时活性升至峰值(63.28 U/g)与对照接近,打孔的LOX活性在12 h明显下降,之后缓慢升高,72 h后活性与对照接近。烟蚜取食6 h时快速诱导烟叶内LOX活性显著升高达到峰值(72.40 U/g),较对照上升了54.01%,其余时段内活性接近或略低于对照;针刺前期烟叶内LOX活性与其他处理组相同,均受到一定抑制,之后活性回升。表明棉铃虫和斜纹夜蛾取食下烟叶内LOX酶活性在一定时间内较对照明显受到抑制,而烟蚜取食可快速诱导烟叶内LOX活性快速显著升高(图2)。
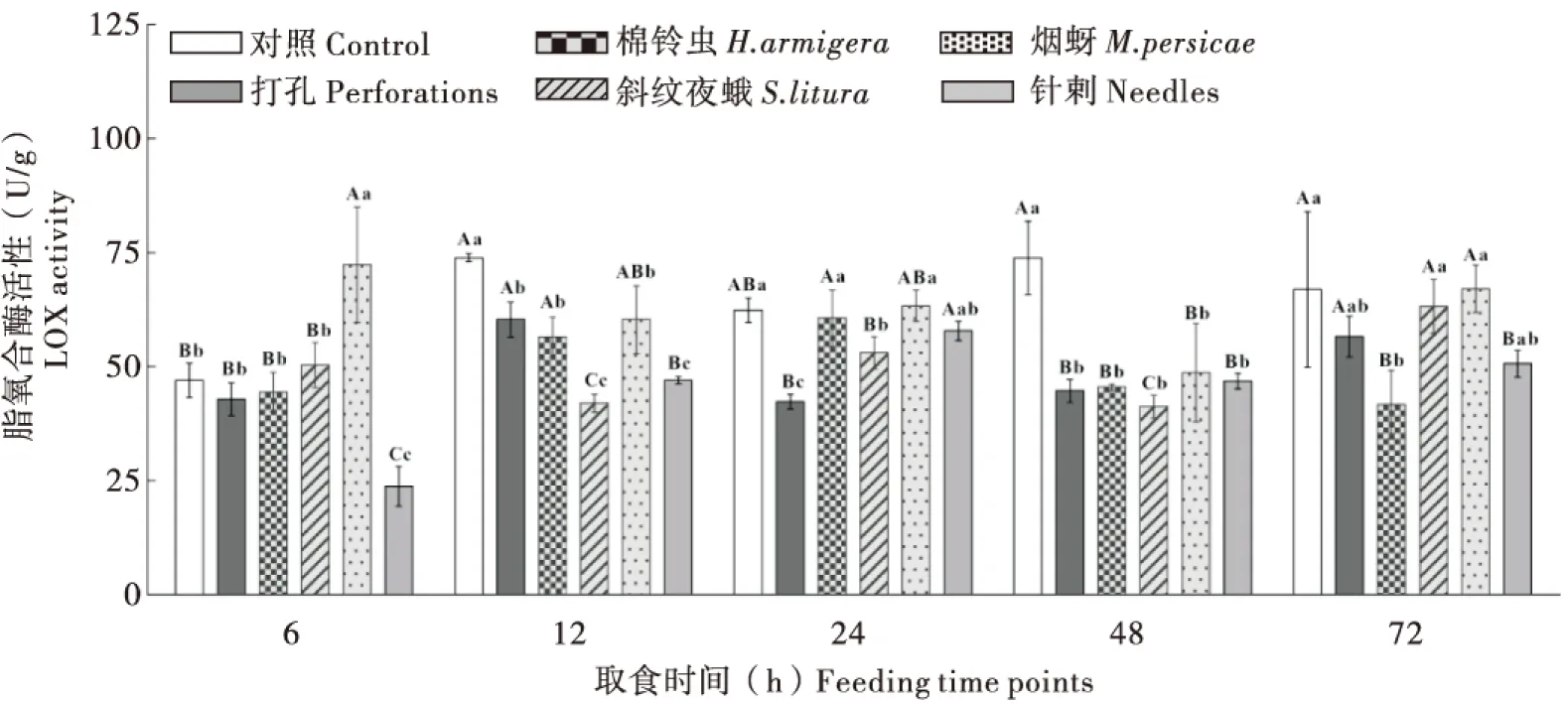
图2 不同昆虫取食对烟草‘MS K326’叶片中防御酶LOX活性的影响Fig.2 Dynamics of the LOX activity of tobacco ‘MS K326’ leaves infested by different insects at different feeding time points
2.2.3不同昆虫取食对烟草‘MS K326’叶片中PPO活性的影响
棉铃虫和斜纹夜蛾取食后烟叶内PPO活性均升高,棉铃虫取食后烟叶PPO活性在6 h时较对照受到明显抑制,48 h时出现活性峰值(190.66 U/g);斜纹夜蛾取食下PPO活性自6 h时开始升高,12 h时达到峰值(195.32 U/g),之后保持稳定;打孔的烟叶内PPO活性在24 h升至最高,其峰值低于咀嚼口器昆虫取食诱导的PPO活性。烟蚜取食烟株后烟叶内PPO活性呈现升高-降低-升高的趋势,取食6 h时,烟叶内PPO活性升至最高(156.61 U/g),较同时段对照明显升高;针刺诱导烟叶内PPO活性变化与烟蚜取食的情况存在一定差异,但同在处理6 h时活性升高且接近烟蚜组活性(活性略低于烟蚜取食),之后变化情况与烟蚜组相似。以上结果说明,不同昆虫均能引起烟叶内防御酶PPO活性升高,其中斜纹夜蛾诱导的烟株防御酶活性升高最高(图3)。
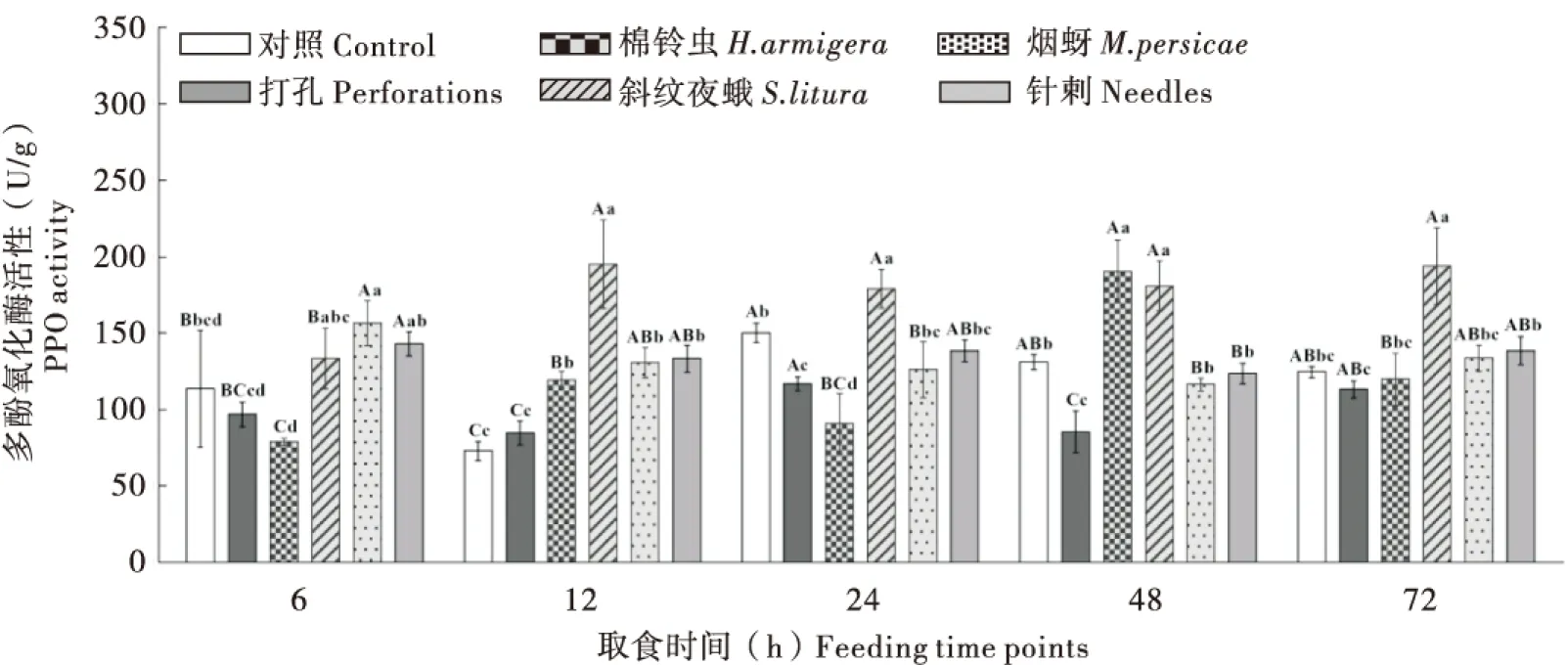
图3 不同昆虫取食对烟草‘MS K326’叶片中防御酶PPO活性的影响Fig.3 Dynamics of the PPO activity of tobacco ‘MS K326’ leaves infested by different insects at different feeding time points
2.2.4不同昆虫取食对烟草‘MS K326’叶片中POD活性的影响
棉铃虫取食初期烟叶内POD活性受到明显抑制,之后逐渐升高,48 h时升至峰值(99.65 U/g)接近同时段对照,斜纹夜蛾取食烟株后烟叶内POD活性呈现降低后升高趋势,24 h时升至最高(169.33 U/g),较对照显著升高1.11倍;打孔烟叶内POD酶活性升高,呈现波动性变化,活性峰值介于棉铃虫与斜纹夜蛾之间。烟蚜取食烟株后烟叶内POD活性较对照被诱导显著升高,12 h时为活性峰值点(161.48 U/g),接着活性降低,之后逐渐升高;针刺烟叶POD活性呈现升高-降低-升高取食,其活性峰值较烟蚜处理组滞后。由此可见,不同昆虫取食均可诱导烟叶内POD酶活性升高,其中斜纹夜蛾取食烟草诱导POD酶活性峰值最高(图4)。
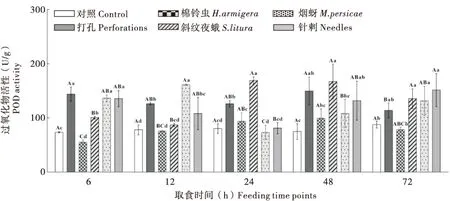
图4 不同昆虫取食对烟草‘MS K326’叶片中防御酶POD活性的影响Fig.4 Dynamics of the POD activity of tobacco ‘MS K326’ leaves infested by different insects at different feeding time points
3 结论与讨论
虫害诱导的寄主植物体内各信号物质变化直接影响到植物防御信号的通路转导及相关防御基因的表达,从而增加植物防御抗性(Lietal., 2016)。研究不同虫害对烟草信号物质及防御酶活性变化的影响为筛选与昆虫相关的生防化学激发子提供参考依据。本实验采用生化分析法研究了棉铃虫、斜纹夜蛾和烟蚜3种虫害对烟草防御信号物质JA、NO、H2S和防御酶PAL、PPO、LOX、POD的影响。结果表明,棉铃虫取食可诱导烟草JA、NO和H2S含量均升高;斜纹夜蛾取食下,烟叶JA降低,NO含量升高不明显,H2S含量显著升高;烟蚜为害后JA、NO含量降低,H2S含量在48 h时升至最高,而棉铃虫和斜纹夜蛾取食下H2S含量在6 h时迅速升高。说明棉铃虫比斜纹夜蛾和烟蚜更易诱导烟草内各信号分子含量增加,烟草对棉铃虫取食非常敏感,能够对其为害迅速做出反应,而斜纹夜蛾和烟蚜则在一定程度上抑制了烟草防御信号,此外棉铃虫和斜纹夜蛾咀嚼较烟蚜刺吸可快速诱导烟叶内H2S迅速响应。这些结论与前人报道的棉铃虫为害烟草后JA含量明显升高(Zong and Wang, 2007),烟粉虱Bemisiatabaci以60头/叶的为害密度刺吸危害辣椒后,叶片内JA含量较对照降低,与张海波等(2018)等研究结果相似。但张艳敏等(2017)在温室大棚条件下研究发现烟蚜和斜纹夜蛾取食下烟叶内信号分子JA和NO含量升高后降低,这可能是由于实验环境不同及各类处理烟苗株距导致的诱导效果存在差异。此外本实验发现不同虫害均可诱导H2S信号物质发生强烈反应,且反应时间与取食方式密切相关。
植物在受到害虫取食为害时,通过改变其体内PAL、LOX、POD和PPO等多种防御酶活性从而提高植物抗性(温娟等, 2017; 郭祖国等, 2018)。前人研究报道了棉铃虫取食诱导棉花叶片内POD、LOX、PAL等酶活性升高,烟粉虱为害辣椒和黄瓜后POD活性下降(沙品洁等, 2012; 刘明杨等, 2016)。本实验中,棉铃虫和斜纹夜蛾两者取食烟草诱导的烟草防御存在差异。两者分别取食烟草后,导致烟叶内JA途径关键酶LOX活性降低,PPO活性升高(斜纹夜蛾取食升高明显),SA途径关键酶PAL活性升高,棉铃虫诱导烟叶内POD活性降低至与CK接近,斜纹夜蛾则诱导POD活性增加,其中多数防御酶对斜纹夜蛾的取食防御应答较强。宗娜等(2004)研究发现棉铃虫、烟青虫及斜纹夜蛾3种咀嚼口器昆虫取食对烟草烟碱的诱导情况存在差异原因在于棉铃虫和烟青虫下唇腺中存在葡萄糖氧化酶可能抑制烟碱的诱导,而斜纹夜蛾取食则促进烟碱的诱导,下唇腺提取液中未检测到该酶活性。因此推测导致该实验结果有可能是因为存在于不同植食性昆虫口腔分泌物中的特异激发子有一定关系(Singhetal., 2008; 李静等, 2011)。此外,本实验中烟蚜取食后,各防御酶活性均被诱导升高,其中PAL活性在处理12 h时受到抑制,48 h时活性有所升高,JA途径关键酶LOX和PPO、POD活性在取食6 h时显著升高,之后受到不同程度的抑制,POD活性在24 h抑制最为明显后又逐渐升高,该诱导结果与本实验室之前研究已发表的结果相似。以上结果均说明不同昆虫取食诱导的烟叶内防御信号分子及防御酶活性变化存在显著差异,烟草对3种虫害产生的防御具有专一性。
本研究从虫害对烟草诱导生理防御方面揭示了不同虫害诱导的烟株抗性差异,为棉铃虫、斜纹夜蛾和烟蚜等对种烟草害虫的暴发、植株本身诱导的防御反应监测提供了有效依据。由于不同昆虫与烟草的相互关系存在差异,在以烟草诱导防御机制为重点的绿色防控中,建议应针对性地提出防治措施。同时,外源H2S供体(NaHS)可诱导拟南芥A.thaliana内源H2S含量升高增强对假单胞菌Pseudomonassyringae的抗性(Shietal., 2015)。合理喷施外源JA、NO和H2S等信号物质可提高植株抗性(李顺欣等, 2017; 彭仁义, 2018),该实验研究证实虫害可诱导烟叶内源H2S含量升高,因此在烟草虫害防治中,可考虑合理喷施H2S等相关外源信号物质来提高植物抗虫性,开展相关绿色防控以此降低虫害所造成的损失。
猜你喜欢
ELLE世界时装之苑(2022年11期)2022-05-30 20:23:43
ELLE世界时装之苑(2022年10期)2022-05-30 08:36:21
中国生物防治学报(2021年5期)2021-12-17 11:18:48
河南农业科学(2017年6期)2017-07-05 13:05:17
西南农业学报(2017年4期)2017-05-17 05:43:27
新疆农垦科技(2016年2期)2016-08-21 13:50:22
现代农业(2016年5期)2016-02-28 18:42:35
现代农业(2016年4期)2016-02-28 18:42:09
现代农业(2015年5期)2015-02-28 18:40:42
华东师范大学学报(自然科学版)(2014年1期)2014-04-16 02:54:58
