
不同油橄榄品种苗期快速光曲线模型选择及光合特性分析*
2021-08-11 03:30马成艳刘世增姜成英吴文俊赵梦炯陈炜青金高明戚建莉
西部林业科学 2021年4期
马成艳,刘世增,姜成英,吴文俊,赵梦炯,陈炜青,金高明,戚建莉
(1.甘肃省林业科学研究院,甘肃 兰州 730020;2.甘肃农业大学林学院,甘肃 兰州 730070;3.国家林业局油橄榄工程技术研究中心,甘肃 兰州 730020)
油橄榄(OleaeuropeaL.)为木犀科(Oleeacea)木犀榄属(Olea)亚热带常绿小乔木,是地中海气候的典型植物,也是世界著名的四大木本油料树种之一。橄榄油具有较高的不饱和脂肪酸和多种维生素,长期食用可增强人体消化系统功能,减少心血管疾病,促进骨骼发育。橄榄油还可用于开发医药、保健、化妆品等系列产品,具有重要的药用和保健价值。油橄榄也具有速生、耐旱、耐高温和根系发达等生物学特性,具有保持水土、涵养水源的生态功能,是荒山绿化、生态恢复的可选树种[1-2]。
叶绿素荧光技术可以从内在揭示植物光合生理的表现,在一定程度上能够反映植物对环境的适应情况,被称为是研究植物光合作用的有效探针[3-4]。有关植物叶绿素荧光特性的研究报道较多,但研究各植物种的叶绿素荧光参数所用光合模型也各有不同,主要有直角双曲线模型(RH)、非直角双曲线模型(NRH)、单指数模型(SEM)、双指数模型(DEM)和直角双曲线修正模型(MRH)等,这些模型都存在一些缺点,且不同模型拟合的结果偏差较大[5-8]。因此,应用多种模型对同一研究对象的光响应曲线进行拟合,对拟合优度进行比较,采用最佳拟合模型对相关光合参数进行估算,可提高对这些参数估计的准确性,从而更好地认识供试植物的光合特性。
甘肃陇南白龙江干热河谷区,油橄榄种植面积占全国总面积的80%,建有国家级油橄榄种质资源圃,引进品种130余个。有关油橄榄生理生态特性,已有大量研究报道[9-13]。但对从土耳其引进的在陇南地区表现优良的Domat等油橄榄在生理生态特性方面的研究鲜有报道。本研究采用叶绿素荧光技术测定7个油橄榄品种的叶绿素荧光参数,通过RH、NRH、SEM、DEM和MRH拟合分析各油橄榄品种的叶绿素荧光光响应曲线(RLCs),以期选择最适合的叶绿素荧光光响应模型,根据叶绿素荧光最佳光响应模型,评价各品种的苗期光合特性,为陇南地区引进种植油橄榄的种质高效优质栽培和良种选择提供参考依据。
1 材料与方法
1.1 试验地概况
试验地位于甘肃省陇南市武都区两水林场(104°75′E, 33°37′N),属于北亚热带半湿润气候,年均气温 14.9 ℃,年日照时数1 911.7 h,年无霜期210~240 d,年降水量474~900 mm,年平均湿度61%~71%,是油橄榄的最佳适生区。
1.2 试验材料
采用从土耳其引进的Domat、Odemis、Yamalak sarisi、Memecik、Ayvalik、Cekiste、Gemlik共7个油橄榄品种的嫁接苗,见表1。

表1 7个土耳其引进油橄榄品种的基本情况
引进自砧嫁接苗定植,砧木为原品种半同胞种子实生苗,3 a生;接穗为同一品种优树1 a生枝条,插皮芽接,每砧嫁接4~5芽。2018年春季嫁接,2019年秋季引进,定植到高60 cm,直径80 cm的无纺布营养钵,基质为泥炭土、椰糠和熟土以1︰1︰1的比例混合,每营养钵1株,苗木的接穗生长1 a后作为试验材料。
1.3 测定方法
于2020年8月上旬,选择连续晴朗的3天,每天8:00—11:00进行。每个品种选取3株长势健壮、光照一致的植株。测定前,将油橄榄幼苗叶片先经过 20 min 的充分暗适应,用Li-6400-40便携式荧光叶室测定系统,每株选取3片叶子,每片叶子3个重复,测得初始荧光(Fo),随后施加饱和脉冲光,测得最大荧光(Fm)。并由Fo,Fm计算,暗适应叶片 PSII 最大光化学效率(Fv/Fm),PSII 潜在光化学效率(Fv/Fo)。之后经0.5 h自然光适应,测定在光合有效辐射强度(PAR)分别为2 000、1 500、1 000、500、200、100、50、20、10和0 μmol/(m2·s)时的叶绿素荧光相关参数。使用开放气路,流速为300 μmol/s,外接CO2钢瓶,浓度控制为 400 μmol/mol,最小等待时间设定为120 s,最大等待时间设定为200 s,通过外接缓冲瓶稳定大气中 CO2浓度。测定过程中,每个梯度光照持续时间为 10 s,依次测定。
1.4 数据统计
利用 Excel 2010进行数据整理和作图。用叶子飘光合计算软件4.1.1中5 种光合模型对RLCS及其参数进行拟合,各模型的数学表达式[14-16]见表2。利用origin 8.0得出信息量准则(AIC)、均方根误差(RMSE)。图中的部分数据用平均值±标准误差表示,显著性水平设定为α=0.05。

表2 5种光响应曲线(RLCS)模型的数学表达式
2 结果与分析
2.1 叶绿素荧光—光响应曲线(RLCs)模型拟合结果
R2是衡量所建立模型效果好坏的指标之一,其值越大表明模型拟合得越好,却不能说明所得到的光合参数是否与实测值相符[15]。为了更好地对模型拟合效果进行比较,本研究同时引入了RMSE和AIC评价指标。RMSE表示相对于真实值的离散程度,用来描述误差大小;AIC值越小,说明拟合的模型精度高且简洁[14-16]。因此,叶绿素荧光—光响应曲线模型拟合中,R2值越接近1,RMSE值越小,AIC值越小,则说明模型的拟合效果越好。结果表明,5个模型对7个油橄榄品种的RLCs拟合效果优劣顺序为:MRH>SEM>RH>NRH>DEM。MRH中R2、RMSE和AIC的平均值分别为0.999 4(最大)、10.591 5(最小)和1.419 3(最小),拟合优度最好,是7个油橄榄品种的最佳拟合模型(表3)。

表3 5个模型拟合7个油橄榄品种的RLCs回归参数
通过比较单个油橄榄品种RLCs拟合优度,拟合效果的优劣顺序如下:Domat为MRH>SEM> RH >DEM>NRH,Odemis为RH>DEM>MRH>SEM>NRH,Yamalak sarisi为RH>SEM>DEM>MRH>NRH,Memecik为MRH>RH>SEM>NRH>DEM,Ayvalik为MRH>DEM>RH>SEM>NRH,Cekiste为MRH>SEM>RH>NRH>DEM,Gemlik为MRH>SEM>RH>NRH>DEM。这与整体拟合效果和拟合精度稍有不同。
结果表明,7个油橄榄品种MRH模型拟合RLCs总体趋势:随PAR在0~2 000 μmol/(m2·s)内递增,各油橄榄品种叶片的ETR逐渐增大,且在低光强部分,ETR随PAR的增强上升迅速,达到相对较高光强部分则上升缓慢(图1)。Domat、Odemis、Yamalak sarisi、Memecik、Ayvalik和Gemlik叶片的ETR随PAR的增强而增加。Cekiste叶片的ETR随PAR的增强逐渐增加,达到饱和光强后,随PAR的增强趋于稳定。
2.2 叶绿素荧光—光响应曲线(RLCs)参数比较
7个油橄榄品种叶片的ETR随PAR变化的实测值与MRH模型所得拟合值非常接近。Domat叶片的最大电子传递速率实测值为217.562 7 μmol/(m2·s),MRH模型拟合值为215.737 2 μmol/(m2·s),占实测值99.12%,且R2=0.999 9(图1A)。Odemis、Yamalak sarisi、Memecik、Ayvalik、Cekiste、Gemlik叶片ETR拟合值占实测值的百分比分别为96.49%、99.97%、99.76%、99.98%、99.73%、99.46%。且最大电子传递速率(Rmax)大小顺序为:Memecik>Odemis>Yamalak sarisi>Gemlik>Domat>Ayvalik>Cekiste。
RLCs的初始斜率(α),称为初始量子效率,又称低光强下的光合量子效率,表示植物将光能转化为电子流的能力,是植物在光合作用中光能转化效率的一种度量。其值越大,表明植物叶片在低光强阶段对光能的利用效率越高[17]。MRH拟合各油橄榄的RLCs中,初始斜率(α)大小顺序为:Odemis> Ayvalik> Memecik>Domat>Gemlik>Cekiste>Yamalak sarisi(图1),则说明Odemis对弱光的利用能力最强,对光能的转化效率最高,Ayvalik、Memecik、Domat和Gemlik次之,Yamalak sarisi和Cekiste最弱。

图1 7个油橄榄品种的快速光响应MRH曲线
PARsat表示一定条件下植物的潜在光合作用能力。其值越大,表明植物在强光下的生长发育不易受到抑制[18]。MRH拟合各油橄榄的RLCs中,饱和光合有效辐射(PARsat)大小顺序为:Memecik>Ayvalik>Odemis>Yamalak sarisi>Cekiste>Gemlik>Domat。表明Memecik潜在光合能力最强,对光强的适应范围更广;Ayvalik、Odemis、Yamalak sarisi和Cekiste次之,Gemlik和Domat最弱。
初始荧光Fo是植物PSII所有反应中心均处于开放状态时的最小荧光产量。最大荧光Fm是PSII反应中心完全关闭时的荧光产量,反映了PSII的电子传递情况。Fv/Fm是PSII最大光化学效率,为反应PSII反应中心原初光能转化效率[19-20]。结果表明,Odemis、Memecik和Ayvalik的Fo、Fm、Fv/Fm和Fv/Fo值均高于其它品种,Yamalak sarisi、Cekiste均低于其它品种(表4)。Odemis和Memecik与Yamalak sarisi的Fo值之间存在显著差异。Odemis、Memecik和Ayvalik的Fm值之间差异显著;Yamalak sarisi、Ayvalik的Fv/Fm的值与其他品种间都存在显著差异,Cekiste与Yamalak sarisi、Memecik和Ayvalik的Fv/Fm的值之间存在显著差异;Ayvalik的Fv/Fo的值与其他品种间都存在显著差异,Yamalak sarisi与Memecik、Ayvalik和Gemlik的Fv/Fo的值存在显著差异。
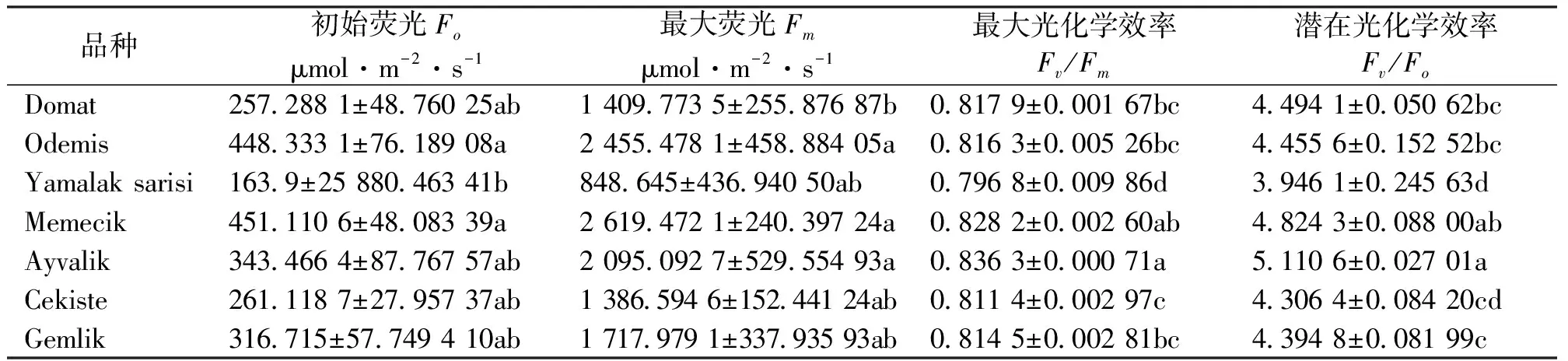
表4 7个油橄榄品种叶片叶绿素荧光参数的比较
2.3 PSII的实际光化学效率(PhiPS2)和电子传递速率(ETR)的光响应曲线
PhiPS2 为PSII反应中心的实际光化学效率,表示植物在光合作用过程中PSII系统反应中心部分关闭时的原初光能捕获效率,表征植物在光照条件下PSII吸收光能用于原初反应阶段的实际运行效率,可快速地确定光强变化时或其他环境条件下PSII反应中心的作用效率[21-25]。随着光强在0~2 000 μmol/(m2·s)内递增,7个油橄榄品种叶片PhiPS2的总体趋势都逐渐减小,范围在0.18~0.78,各品种间PhiPS2无显著性差异(P<0.01)(图2)。且在低光强部分〔0~200 μmol/(m2·s)〕,PhiPS2随光强的增加而缓慢减小,到达相对较高光强部分迅速减小。在光强为1 500~2 000 μmol/(m2·s)内,Memecik叶片PhiPS2整体水平高于其它品种,Cekiste叶片PhiPS2低于其它品种;Ayvalik、Domat、Gemlik、Yamalak sarisi、Odemis间PhiPS2无明显差异。

图2 7个油橄榄品种的PSII实际光化学效率 (PhiPS2)的光响应曲线Fig.2 Response of actual photochemical efficiency of PSII(PhiPS2)to light intensity in seven olive cultivars
植物RLCs中ETR随光强的变化规律,可以反映植物叶片对光强变化的适应机理。在光强相同的情况下,植物叶片的ETR越高,碳同化过程中形成的电子传递载体(ATP 和 NADPH)就越高[10,14]。随着光强在0~2 000 μmol/(m2·s)内递增,各油橄榄品种的ETR逐渐增大,各品种间ETR无显著性差异(P<0.01)(图3)。在低光强部分[0~200 μmol/(m2·s)],ETR随光强的增加而缓慢上升,到达相对较高光强部分则上升迅速。在光强低于1 000 μmol/(m2·s)时,各品种间ETR整体水平无明显差异。在光强1 500~2 000 μmol/(m2·s)内,Odemis和Memecik叶片的ETR随光强增加而增大的速度最快,且高于其它5个品种(α=0.05);Domat、Yamalak sarisi、Ayvalik、Gemlik上升速度较慢,但高于Cekiste;Cekiste在光强1 500~2 000 μmol/(m2·s)范围内ETR无明显差异,但低于其它6个品种(图3)。
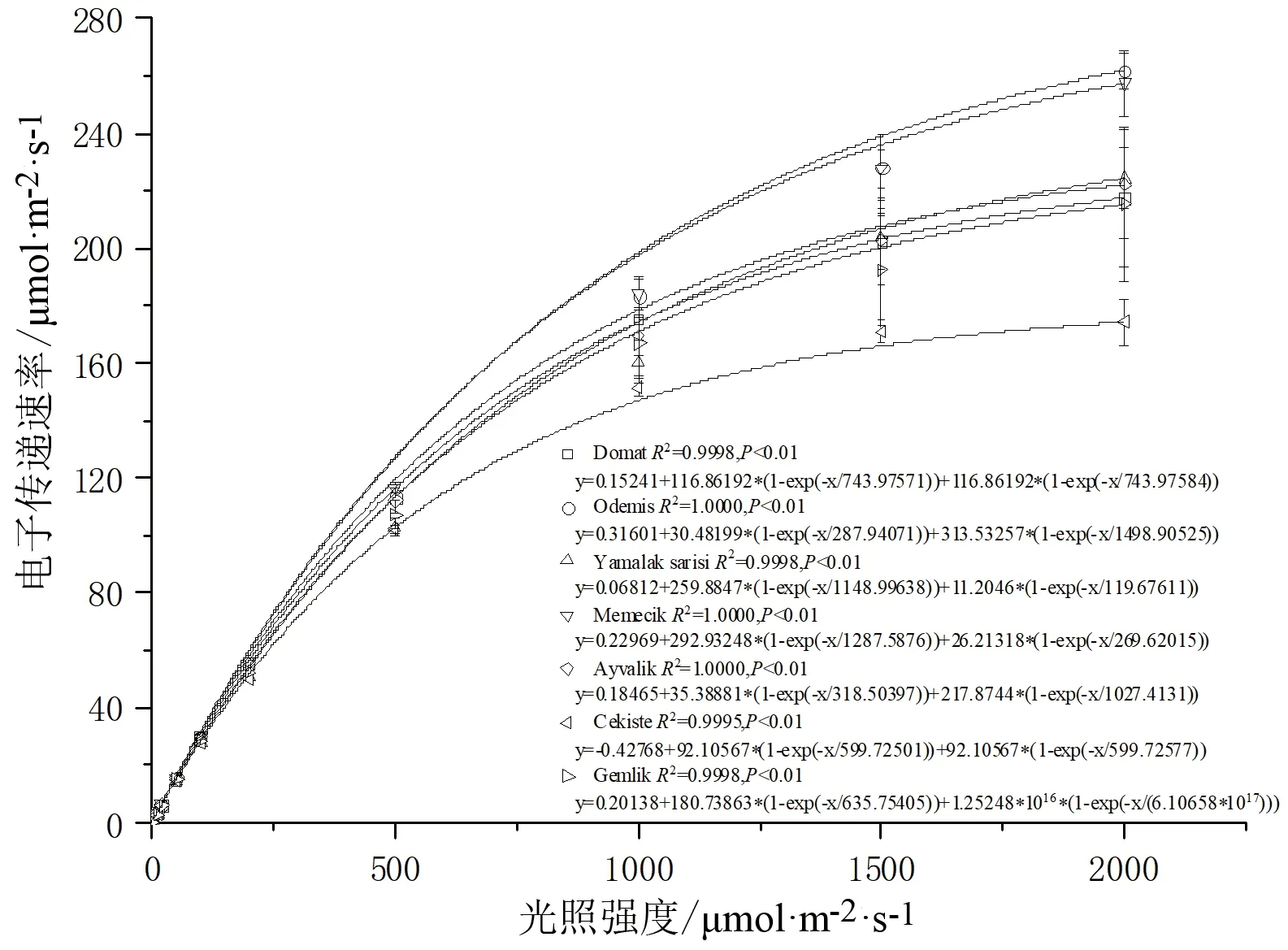
图3 7个油橄榄品种的电子传递速率光响应曲线Fig.3 Response of electron transfer rate to light intensityin seven olive cultivars
3 讨论与结论
3.1 讨论
光响应曲线是植物光合作用随着光照强度改变的系列反应曲线,测定该曲线可获得植物光合特性的相关生理参数,有助于判定植物的光合效率,且不同植物有其最佳的光响应曲线模型[7]。因为,RH、NRH和SEM的拟合方程是无极值的渐近线,不能直接得出植物的光饱和点,不能合理地拟合植物在光抑制条件下的光响应曲线。SEM和DEM通常只用于藻类、浮游植物和少数陆生植物,且DEM拟合值远高出实际值的结果[8,26-27]。在模型拟合的研究中,MRH拟合方程存在极值,能较好地拟合植物存在PSII动力学下调或光抑制时的光响应曲线,且拟合曲线与实测值高度符合[28-31]。本研究通过分析各模型的拟合优度,得出MRH是7个油橄榄品种的最佳拟合模型(表3)。同时,各油橄榄品种MRH模型拟合曲线与实测值的符合程度高达95%以上(图1)。因此,上述理论在本研究中也得到了验证。
光合电子流对光的响应曲线一般包括以下3个阶段:①ETR随PAR的增加而几乎线性增加;②ETR随PAR的增加缓慢增加到一个平台;③ETR随PAR的增加而下降,出现光PSII的动力学下调或光抑制现象阶段[16]。本研究中,在PAR为0~2 000 μmol/(m2·s)条件下,Domat、Odemis、Yamalak sarisi、Memecik、Ayvalik和Gemlik叶片的ETR随PAR的增强而快速增加。在设定最大光照条件2 000 μmol/(m2·s)下,各品种叶片的ETR均未出现下降趋势,即均无PSII的动力学下调现象。Cekiste叶片ETR随PAR增强缓慢增强并趋于饱和,这表明除Cekiste,其它6个油橄榄品种都可以承受较高的光辐射。
已有研究表明,植物Fv/Fm在正常情况下为0.80~0.85[19]。本研究中,7个油橄榄品种的Fv/Fm值接近于0.80或大于0.80,而小于0.83,说明7个油橄榄品种苗木生长状况良好,也侧面说明了其他荧光指标的可靠性。在7个油橄榄品种中,Odemis、Memecik和Ayvalik的PSII反应中心处于完全开放和完全关闭时的荧光产量较高,具有较高的PSII活性,但Yamalak sarisi和Cekiste与之相反。
本研究中,随着光强增强各油橄榄品种的PhiPS2呈下降趋势,最低达到了0.18,推测高光强迫使各油橄榄品种叶片PSII反应中心受到了光抑制,降低了各油橄榄品种的实际光能转化效率,使PSII潜在活性受损,光合作用原初反应过程受到了抑制。Memecik的PhiPS2整体水平高于各品种,Cekiste低于其它6个品种(图2)。随着光强增强各油橄榄叶片的ETR都明显增大,一定程度的高光强有利于提高各油橄榄品种的PSII反应中心开放程度,将更多的光能用于推动各油橄榄叶片的光合电子传递,提高了各油橄榄品种的光合效率。Memecik的ETR值明显高于各品种,Cekiste低于其它6个品种(图3),说明在相同光照条件下,Memecik对光照的响应能力最强,具有较大的潜在光合能力,Cekiste则相反。
胁迫导致植物 PSII 开放反应中心开放程度减少,引起 PSII 有效光量子产量的下降,从而导致光合电子传递效率的明显降低,电子传递过程明显受到抑制。本研究中,Memecik叶片PhiPS2值整体水平明显高于其它品种,且随光强的变化幅度最小,其ETR值明显也高于其它品种。因此,可说明Memecik具有较好的光合能力;Cekiste叶片PhiPS2值低于其它品种,随光强的变化幅度最大,且其ETR值显著低于其它品种,则说明Cekiste光合效率较差。
3.2 结论
MRH是7个油橄榄品种RLCs的最佳拟合模型,R2值最接近1,RMSE和AIC值最小,拟合曲线与实测值高度符合。Memecik具有较好的光合特性,将光能转化为电子流的能力和潜在光合能力较强,且具有较广的光辐射范围。Cekiste的光合特性较差,将光能转化为电子流的能力和潜在光合能力最弱,对高光强的适应能力较差。
猜你喜欢
环境保护与循环经济(2021年7期)2021-11-02
阅读(科学探秘)(2020年8期)2020-11-06
哈尔滨轴承(2020年1期)2020-11-03
中国奶牛(2019年10期)2019-10-28
中国果业信息(2019年1期)2019-01-05
电子制作(2018年23期)2018-12-26
中成药(2018年4期)2018-04-26
中国果业信息(2017年8期)2017-09-12
生物学教学(2017年9期)2017-08-20
绿色科技(2015年5期)2015-07-05
