
伴矿景天耐镉内生菌的筛选
2021-08-09 08:11:50周毅吉徐远芳李文革张祺玲
湖南农业科学 2021年6期
郭 峰,周毅吉,徐远芳,李文革,彭 玲,张 勇,邓 超,张祺玲
(湖南省核农学与航天育种研究所,湖南 长沙 410125)
随着城市化、工业化、农业集约化的快速发展,环境污染问题日渐突出[1]。2014 年 4 月我国环境保护部和国土资源部公布的《全国土壤污染调查公报》显示,我国土壤主要遭受以重金属为代表的无机物污染,其中镉是主要的重金属污染物之一,存在于各种土地类型中,含量分布呈现从西北到东南、从东北到西南方向逐渐升高的态势,点位超标率达到7%[2]。研究高效便捷的镉污染修复技术刻不容缓。
目前,大部分物理化学修复方法不仅成本高昂,而且会对土壤生态系统造成一定破坏[3]。这使得重金属镉污染土壤生态修复技术研究成为多个科技研究领域的热点和难点。虽然已发现一些超累积植物能吸收高浓度的重金属镉[4-5],但是这些植物大多数生物量极低且生长缓慢,同时对重金属存在偏好,从而制约了其在土壤重金属污染中的修复效率[6]。因此,制定适宜的植物修复强化策略对修复重金属镉污染的土壤具有重要意义。为了提高植物修复效率,许多学者研究了植物-微生物-重金属之间的相互作用,发现某些微生物具有减轻重金属诱导的植物毒性和增加生物量的作用[7-9]。从此,增强植物修复体系中微生物的功能成为生物修复技术研究的一个重要方向。
植物和内生菌通过长期协同进化形成互惠互利的关系,彼此构成了稳定的生态关系,维持着自然系统生态平衡[10]。研究表明,内生菌产生的代谢物,或能刺激植物的生长发育,提高宿主植物对生物胁迫或非生物胁迫的抵抗能力,在污染土壤植物修复技术领域展现出巨大的应用潜力[11]。该研究从镉污染区域生长的超富集植物伴矿景天植株中分离筛选获得2 株具有耐镉特性的细菌菌株,通过形态观察、生理生化特性及16S rDNA 序列和系统发育树将其鉴定为无色杆菌和伯克霍尔德氏菌,同时对菌株的耐镉能力进行了评估,为耐镉内生菌株的开发应用提供依据。
1 材料与方法
1.1 试验材料
供试植株伴矿景天(Sedum plumbizincicola)采集自湖南省长沙市长沙县高桥镇镉污染区域,取样后于48 h 内用其根茎叶健康组织进行植物内生菌的富集和分离。用于细菌培养的LB 培养基配方为:胰蛋白胨10 g、酵母提取物 5g、NaCl 10 g、固体培养基另加琼脂粉15~20 g,加水至1 000 mL,pH 值7.2,121℃灭菌30 min。耐镉细菌筛选培养基,即在LB 培养基中加入 CdCl2溶液。
1.2 试验方法
1.2.1 耐镉内生菌株的分离 将伴矿景天植株根茎叶用清水洗净,75%乙醇浸泡1 min,5%NaClO 浸泡1 min,无菌水冲洗4 次,经试验验证消毒效果后在无菌条件下研磨植株组织,置于Cd2+浓度为100 mg/L的LB 培养基中,30℃下150 r/min 振荡培养3 d,采用稀释涂布法分离耐镉内生菌,再分别进行划线纯化。
1.2.2 耐镉内生菌株的筛选 取分离纯化菌株一环置于LB 培养基中,30℃下150 r/min 振荡培养24 h,分别取0.2 mL 培养液涂布于Cd2+浓度分别为200~1 000 mg/L 的LB 固体培养基,将培养基置于30℃的恒温培养箱中培养48 h,观察菌株生长情况,筛选出耐受能力较高的菌株,斜面保存。
1.2.3 耐镉菌株的生理生化特性分析 挑取耐镉菌株的单菌落接种于LB 培养基中,30℃下150 r/min 振荡培养24 h,参照《微生物学实验技术》[12]进行革兰氏染色;参照《常见细菌系统鉴定手册》[13]及《伯杰细菌鉴定手册》[14]进行细菌常规的生理生化鉴定试验。
1.2.4 耐镉菌株16S rRNA 序列分析及系统发育树构建 采用DNA 提取试剂盒提取细菌DNA,采用细菌通 用 引 物27F(5-′AGAGTTTGATCCTGGCTCAG-3′)和1492R(5′-TACGGYTACCTTGTTACGACTT-3′)进行16S rRNA 扩增。PCR 反应体系(50 μL)为:PCR Mix 25 μL,27F(10 μM)2 μL,1492R(10 μM)2 μL,DNA 模板1 μL,ddH2O 20 μL。PCR 反应条件为:98℃2 min; 98℃ 10 s,54℃ 10 s,72℃ 10 s,35 个循环;72℃5 min 终止。PCR 产物经凝胶电泳检测,采用凝胶回收试剂盒切胶回收后,交由生工生物工程(上海)股份有限公司测序。测序结果在NCBI 数据库进行Blast同源比对分析,利用MEGA 软件构建16S rRNA 基因系统发育树。
1.2.5 镉离子对菌株生长的影响 耐镉菌株分别接种于LB 培养基中,30℃下150 r/min 振荡培养24 h,以2%的接种量分别接种于Cd2+浓度为0、50、100、200 和400 mg/L 的200 mL LB 培养基中振荡培养,每24 h取样1 次,用紫外可见分光光度计测定不同培养时间的培养液在600 nm 处的OD 值,研究Cd2+浓度对菌株生长特性的影响。
1.2.6 耐镉菌株对不同抗生素耐性研究 采用K-B 药敏纸片扩散法对耐镉菌株进行药物敏感性试验。吸取浓度为108CFU/mL 耐镉菌液0.1 mL 涂布于LB 固体培养基上,待培养基上菌液稍干,用无菌镊子夹取药敏片平放在培养基上,每个培养基均匀放置3 片药敏片,30℃下培养48 h,测量菌抑菌圈大小,参照美国临床和实验室标准协会 (Clinical and Laboratory Standards Institute,CLSI)推荐的纸片扩散法标准进行判定。抗生素药敏片包括:氯霉素、四环素、环丙沙星、卡那霉素、诺氟沙星、红霉素、链霉素、氨苄西林、青霉素和庆大霉素。
1.2.7 耐镉菌株能谱分析 将耐镉菌株分别接种于LB 培养基中,30℃下150 r/min 振荡培养24 h,以2%的接种量分别接种于Cd2+浓度为0、100 和500 mg/L的LB 培养基中,30℃下振荡培养24 h,细菌菌体经去离子水洗涤3 次,真空干燥后应用能谱仪对菌体元素分布进行定性定量分析。
1.2.8 耐镉菌株红外光谱分析 耐镉菌株分别接种于LB 培养基中,30℃下150 r/min 振荡培养24 h,以2%的接种量分别接种于Cd2+浓度为0、100 和500 mg/L的LB 培养基中,30℃下振荡培养24 h,细菌菌体经去离子水洗涤 3 次后,真空干燥,取1 mg 干燥样品与100 mg KBr(光谱纯)磨细混匀,在10 t/cm2下压成薄片,用 FTIR 光谱仪测定并记录红外光谱[15-17]。
2 结果与分析
2.1 耐镉内生细菌菌株的分离筛选
经Cd2+浓度为100 mg/L 的LB 固体培养基上多次分离纯化,共分离出8 种菌落形态不同的耐镉内生菌株,8 种菌株经Cd2+浓度梯度筛选培养,最高耐受浓度见图1。从图1 可以看出,8 株细菌均能在Cd2+浓度为400 mg/L 的固体培养基上生长,其中菌株JGY61 和JGG81 的最高耐受浓度分别达800 和850 mg/L,因此选择菌株JGY61和JGG81进行下一步研究。

图1 耐镉内生细菌菌株最高耐受浓度
2.2 耐镉菌株生理生化特性及16S rRNA 序列分析
2.2.1 耐镉菌株的生理生化特性 菌株JGY61 在光学显微镜下菌体呈短杆状,革兰氏染色阴性;菌株JGG81 在光学显微镜下菌体呈杆状,革兰氏染色阳性。2 个菌株部分生理生化特征见表1。

表1 耐镉内生细菌菌株生理生化试验结果
2.2.2 耐镉菌株的16S rRNA 序列分析 2 个菌株DNA 经过PCR 扩增出来的产物大小约为1 500 bp,其中菌株JGY61 的16S rDNA 序列扩增长度为1 461 bp,菌株JGG81 的16S rDNA 序列扩增长度为1 465bp。测序序列通过Blast 同源比对分析后利用MEGA软件构建16S rRNA 基因系统发育树如图2 所示,菌株JGY61 与无色杆菌属的细菌菌株亲缘关系最近,菌株JGG81 与伯克霍尔德氏菌属的细菌菌株亲缘关系最近。综合菌株生理生化特性、16S rRNA 序列及系统发育树可知,分离到的耐镉内生菌株JGY61 和JGG81 分别为无色杆菌和伯克霍尔德氏菌。
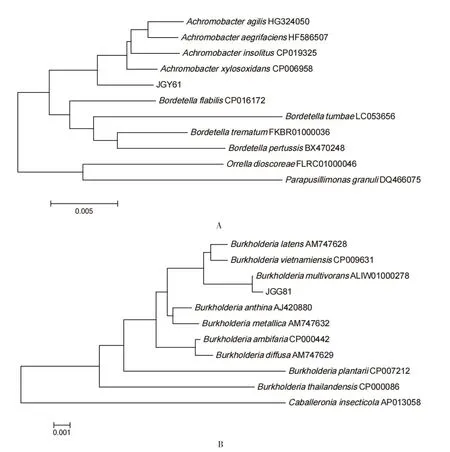
图2 耐镉内生细菌菌株的16S rDNA 序列系统发育树
2.3 Cd2+浓度对耐镉内生菌株生长的影响
由图3 可知,随着Cd2+浓度的增加,菌株的生长期逐渐延长,但当Cd2+浓度达400 mg/L 时,菌株稳定期OD 值均未受明显影响,说明菌株可能通过某些方式对Cd2+浓度进行适应性调整,减弱Cd2+对菌株的影响,使菌株具有较强的耐镉能力,从而不影响菌株的生物量。

图3 不同Cd2+浓度处理下菌株的生长曲线
2.4 耐镉菌株对不同抗生素的耐性
由表2 可知,菌株JGY61 对四环素类抗生素四环素敏感,对糖肽类抗生素卡那霉素、β-内酰胺类抗生素氨苄西林、四环素类抗生素氯霉素和喹诺酮类抗生素环丙沙星耐性中介,对其余抗生素耐性较强;菌株JGG81 对四环素类抗生素氯霉素和四环素敏感,对糖肽类抗生素卡那霉素和喹诺酮类抗生素环丙沙星耐性中介,对其余抗生素耐性较强。

表2 耐镉内生菌株对 10 种抗生素的敏感性结果
2.5 耐镉内生菌株菌体能谱分析
由图4 可以看出,菌株JGY61 和JGG81 在有Cd2+条件下菌体细胞壁均能检测出镉元素存在,在Cd2+浓度为100、500 mg/L 条件下培养后,菌株JGY61 菌体表面镉元素分别占总元素的0.6%、40.2%(原子百分比),菌株JGG81 菌体表面镉元素分别占总元素的1.5%、2.8%(原子百分比),表明菌株可能对镉离子存在不同程度的表面吸附作用。

图4 耐镉菌株的菌体能谱分析图谱
2.6 耐镉内生菌株菌体红外分析
从图5 可以看出,Cd2+处理后耐镉细菌细胞壁表面化学基团发生变化的主要有:3 300 cm-1左右来自蛋白质C—H 键、N—H 键的伸缩以及碳水化合物中结合水O—H 的伸展振动;2 927 cm-1左右饱和C—H 键;1 650 cm-1左右酰胺(O=CN—N)Ⅰ带C=O 的伸缩振动;1 540 cm-1左右蛋白质酰胺II 带N—H 的弯曲振动与C—N 伸展振动的叠加;1 395 cm-1左右—CH2、CH3、COO—等 基 团;1 240 cm-1左 右P=O 和C—S 的伸缩振动以及C—O 与O—H 的叠加吸收峰;1 080 cm-1左右—PO43-、胺基中的C—N 的伸缩振动和糖环的振动吸收带;550 cm-1左右M—O 和O—M—O (M—金属离子) 振动吸收峰。低浓度Cd2+处理下菌体细胞壁表面化学集团主要发生轻微位移,高浓度Cd2+处理下1 080 cm-1左右—PO43-、胺基中的C—N的伸缩振动和糖环的振动吸收带以及550 cm-1左右—M—O 和O—M—O( M—金属离子)振动吸收峰变得强而宽,1 540 cm-1左右蛋白质酰胺Ⅱ带N—H 的弯曲振动与C—N 伸展振动的叠加吸收峰变弱,说明参与积累重金属的化学官能团主要有—PO43-、—M—O(O—M—O)、胺基中的C—N 和酰胺基(—CO—NH—)基团。

图5 耐镉菌株的菌体红外光谱图
3 结论与讨论
研究以镉污染区域生长的伴矿景天为材料,从中分离到8 种菌落形态不同的耐镉内生菌株,经Cd2+浓度梯度筛选得到2 株耐镉能力较强的菌株,最高耐受浓度分别达800 和850 mg/L,经形态观察、生理生化特性研究以及16S rDNA 序列和系统发育树分析,将2 株菌株分别鉴定为无色杆菌和伯克霍尔德氏菌。耐镉菌株在不同镉浓度培养基中的生长曲线测定结果表明,菌株随着Cd2+浓度的增加生长期逐渐延长,说明在Cd2+影响下菌株生长存在适应性调整过程;但经适应性调整后在培养基中Cd2+浓度达400 mg/L 时2 株菌株的生物量均未受明显影响,该调整机理有待进一步研究。综上所述,筛选获得的2 株伴矿景天内生细菌有较好的耐镉能力,为镉污染土壤植物-微生物联合修复提供了宝贵的微生物资源。
对2 株耐镉内生菌进行了抗生素耐性测定,结果显示,菌株JGY61(无色杆菌)对四环素敏感,对卡那霉素、氨苄西林、氯霉素和环丙沙星耐性中介,对青霉素、庆大霉素、链霉素、红霉素、诺氟沙星耐性较强;菌株JGG81(伯克霍尔德氏菌)对氯霉素和四环素敏感,对卡那霉素和环丙沙星耐性中介,对青霉素、氨苄西林、庆大霉素、链霉素、红霉素、诺氟沙星耐性较强。
相关研究表明,微生物与重金属镉的相互作用方式主要有4 种:表面吸附、胞外络合、胞外沉淀和微生物积累[18]。对该研究分离获得的耐镉内生菌株进行能谱分析,发现菌株在Cd2+存在条件下菌体细胞壁均能检测出镉元素存在,表明菌株可能对镉离子存在不同程度的表面吸附作用。红外分析结果表明,—PO43、—M—O(O—M—O)、胺基中的C—N 和酰胺基(—CO—NH—)基团是参与菌株表面吸附的主要官能团。由此初步证实,菌体细胞壁及其官能团在吸附镉中的作用,其耐性与解毒机理有待进一步加以揭示。
猜你喜欢
当代水产(2022年1期)2022-04-26 14:35:38
黑龙江大学自然科学学报(2021年4期)2021-11-19 07:05:02
快乐语文(2020年26期)2020-10-16 03:37:00
湖北农机化(2020年4期)2020-07-24 09:07:16
世界农药(2019年4期)2019-12-30 06:25:10
今日农业(2019年11期)2019-08-15 00:56:32
乡村地理(2018年2期)2018-09-19 06:44:02
中国调味品(2017年2期)2017-03-20 16:18:21
现代检验医学杂志(2016年3期)2016-11-15 01:59:48
植物营养与肥料学报(2010年4期)2010-11-06 07:28:54
