
荒漠绿洲过渡带一年生草本植物对干旱胁迫的响应
2021-08-06 10:23席璐璐缑倩倩王国华
生态学报 2021年13期
席璐璐,缑倩倩,*,王国华,2,宋 冰
1 山西师范大学,地理科学学院,临汾 041004 2 中国科学院西北资源与环境研究院 中国生态系统研究网络临泽内陆河流域研究站,兰州 730000
一年生草本植物是干旱半干旱荒漠生态系统植物区系的恒有层片[1],广泛分布于荒漠区土质、沙质、砾质、石质、盐碱土等生境,对于荒漠生态系统生物多样性的维持和生态系统功能稳定发挥着不可替代的作用[2]。目前,国内外关于荒漠地区一年生草本植物的研究已有大量报道。国外主要对一年生草本植物的生态意义和生存对策、物种丰富度和多样性、生长、繁殖和生产力以及季节动态等方面开展了研究[3- 6]。例如,Madon和Medail[3]对地中海一年生草本植物研究发现一年生植物具有高度的耐胁迫能力;Brown[4]在科威特荒漠发现,在三个生长季节一年生草本植物的生物量受初雨的发生时间,生长期的降雨量及时间分布影响;Krieger等[5]发现一年生草本植物生命周期短,繁殖率高,对于干旱有很强的适应能力;Robinson[6]发现在阿曼干旱林地荒漠,多年生植物可以通过叶片截留并重新分配降雨对一年生草本植物的生长和丰度的空间格局产生影响。我国学者主要研究了荒漠地区一年生植物层片组织格局、生态适应模式[1]、物种多样性[7]、分布特征[8- 9]、丰富度的季节变化及不同生活型植物生物量特征[10]、叶片对凝结水响应[11],更多关注干旱与半干旱地区荒漠群落一年生层片对于荒漠生态系统稳定和防止土地荒漠化的重要作用[1,7- 18]。同时学者们还对荒漠地区一年生草本植物种子萌发[12- 14]和土壤种子库[15- 16]、一年生植物的生理生态特征,例如光合速率[17]、蒸腾速率和气孔导度[18]进行研究,阐释了一年生植物在生理特性上对环境的生态适应性,而目前探讨一年生植物对干旱胁迫的适应机制的研究还相对有限。
在干旱荒漠地区,降水稀少、不连续、不可预测,且小降水事件发生频率高、降水间隔期长[19],一年生草本植物作为机会主义者,高度顺应气候波动,对干旱胁迫和水分供应高度敏感,并能够在高温干旱时期通过调节生理、生长和繁殖策略,快速完成生活史,虽植株生长矮小,但结实较多[7,18]。生长在干旱半干旱荒漠生态系统的一年生草本植物,在长期进化中形成了复杂多样的干旱胁迫适应对策,因此,一年生草本植物对干旱胁迫的响应机制研究一直是干旱荒漠生态系统研究的重点内容。例如,宋士伟等[20]通过对野大麦(Hordeumbrevisubulatum)幼苗进行生理指标测定,研究表明,野大麦在遭受干旱胁迫的情况下,野大麦叶片脯氨酸含量呈连续上升趋势,可溶性糖呈先上升后下降趋势;郭郁频[21]也通过设置不同干旱胁迫条件,研究表明,随着干旱胁迫的加剧,早熟禾(Poa)幼苗叶片叶绿素含量呈下降趋势,脯氨酸含量、丙二醛含量总体呈上升趋势;李秋艳等[22]通过人工控制降水量处理水平模拟幼苗生长对生长季节内降水量变化的响应,研究表明,5种荒漠植物能够调节对地上地下生物量的分配来适应环境变化;徐贵青等[23]对3种荒漠植物根系研究表明,根系在一定程度上具有向土壤湿润区域发展的向水特性。目前,更多学者关注植物对增温和控制降水量导致的干旱胁迫的响应,而天然环境下过长的降水间隔是影响植物的生理和生长的直接因素,但目前相关研究鲜有报道,导致我们还无法系统全面了解一年生草本植物对气候干旱的响应。
河西走廊地处我国西北干旱内陆地区,是重要的农业生产基地,但由于深居内陆、干旱少雨、风沙活动频繁,绿洲沙漠化严重,该区也是中国西北主要的沙漠化防治区[24]。该区通过在荒漠绿洲过渡带建立人工固沙植被,保持绿洲生态环境稳定。经过近50年人工固沙植被的建立,人工林内灌木和多年生草本植物入侵稀少,而大量一年生草本植物入侵并定居,并成为人工固沙植被群落中草本层片的优势植物层片,对防治近地表风沙活动和保持沙面稳定发挥着至关重要的作用[25]。该地区一年生植物主要由一年生禾本科、一年生藜科、十字花科组成,其中一年生禾本科虎尾草(Chlorisvirgata)和狗尾草(Setariaviridis)与一年生藜科雾冰藜(Bassiadasyphylla)是主要的优势种。由于降水匮乏,蒸发强烈,年际降水量波动大,年内降水分布极度不均匀,干旱胁迫成为该地区一年生草本植物最关键的影响因素。本文以河西走廊荒漠绿洲过渡带典型一年生草本植物(雾冰藜、虎尾草和狗尾草)为研究对象,通过模拟不同降水间隔期对其进行胁迫处理,研究3种一年生草本植物在不同干旱胁迫下生理、生长和繁殖的响应规律,探讨植物生理指标之间的关系,以期揭示干旱半干旱地区一年生草本植物的抗旱机制,进而为荒漠生态系统的科学管理提供理论支撑。
1 研究区概况与方法
1.1 研究区概况
研究区位于甘肃河西走廊中部临泽县北部荒漠绿洲过渡带(39°22′—39°23′N,100°07′—100°08′E),是临泽绿洲和巴丹吉林沙漠的交汇处[26],该地海拔约1386m,气候类型属于典型的温带大陆性荒漠气候,年平均气温为7.6℃,多年平均降水量为118.4mm,降水稀少,且分布不均匀,集中在5—8月,年平均蒸发量为2400mm,干旱指数达到20[27]。地带性土壤均为灰棕漠土、沙壤土及沙土。地貌景观类型有流动、半流动、固定、半固定沙丘以及丘间低地。为了防止风沙侵袭及恢复绿洲边缘生态环境,在当地种植梭梭(Haloxylonammodendron)、柠条锦鸡儿(CaraganakorshinskiiKom),红柳(Tamarixramosissima)等人工固沙植被,在人工林内雾冰藜、虎尾草和狗尾草等一年生草本植物不断侵入,成为人工固沙植被草本层的优势植物种,并在防止近地表风蚀发挥着不可忽略的作用。
1.2 试验设计
试验用种子于2018年9月采自河西走廊中国科学院临泽内陆河流域研究站附近的半固定沙丘上,同一物种的种子在20—30个植株个体上收集,自然风干并充分混合(图1)。实验室盆栽试验时间为2019年6月1日—9月15日。花盆内直径为30cm,深20cm。每盆装中沙(粒径0.5—0.25mm)3kg,将纱布铺在花盆底部,阻止沙子漏出,同时可保持通气。选取子粒饱满、大小基本一致且无病虫的3种一年生草本植物种子,每种单独播种于花盆内,每种植物播种20盆,共60盆,每盆播种50粒,行距3cm,播深2cm(图2),为了防止系统误差,60个花盆随机摆放,并在每盆上用标签纸进行标号。用烧杯等量浇水至花盆底部渗出水,控制实验室温度为15/25℃,以保证种子顺利出苗。自播种后每天记录出苗数,在停止出苗3天后,每盆留10株长势良好、生长情况相近的植物进行正常浇水种植。

图1 三种一年生草本植物采集种子照片Fig.1 Seed photos of three annual herbaceous plants

图2 盆栽种植(行距、株距)示意图 Fig.2 The schematic diagram of pot planting (row spacing, plant spacing)
根据临泽站40年降水资料统计,该区多年平均降水量为118.4mm,因此按照多年平均降水量设定试验期间正常水分条件(对照);此外,资料还显示,<10d的降水间隔期占比率最大,为年无降水期的67.56%,10—20d间隔期占13.23%,且频率基本稳定,但>20d的间隔期频率明显下降且变异较大,因而本试验设定5d间隔期(轻度干旱)、10d间隔期(中度干旱)、15d间隔期(重度干旱)来模拟自然降水频率[28]。为减少水分蒸发,尽量保证土壤接受的实际降水量与设定的模拟降水量一致,模拟降水均在同一天的19:00—20:00内完成,并将试验设定的降水量均匀地洒在花盆中。3种植物正常生长至生长期与繁殖期转换期时开始进行干旱胁迫模拟试验并停止浇水,每隔5d采样一次,干旱处理到15d为止,为了测定植物在重度干旱胁迫下的恢复能力,对干旱胁迫15d的植物进行自然复水(DS)并于复水第二天取样。取样时先用水将盆土充分浸泡,将土壤连同植株轻轻倒出,然后用流水慢慢冲洗,洗净根系上所有附泥,样品迅速带回实验室,放入4℃以下低温冰箱保存,测定各项指标。
1.3 测定指标与方法
生理参数:采用TTC法测定根系活力[29];采用80%丙酮法测定叶绿素含量[29];用硫代巴比妥酸法测定丙二醛含量[29];采用酸性茚三酮法测量脯氨酸含量[29];采用蒽酮比色法测定可溶性糖含量[30];用考马斯亮蓝G- 250染色法测定可溶性蛋白质含量[30]。
形态参数:茎长、根长用直接测量法,用精确到0.01cm的直尺测量。茎长是测量根部以上至穗以下的部分;根长是测量植物的主根根长。
繁殖参数:收集全部种子,于实验室晾干至恒重,测定百粒重;并测出单株结种数。以上指标均重复3次,取平均值。
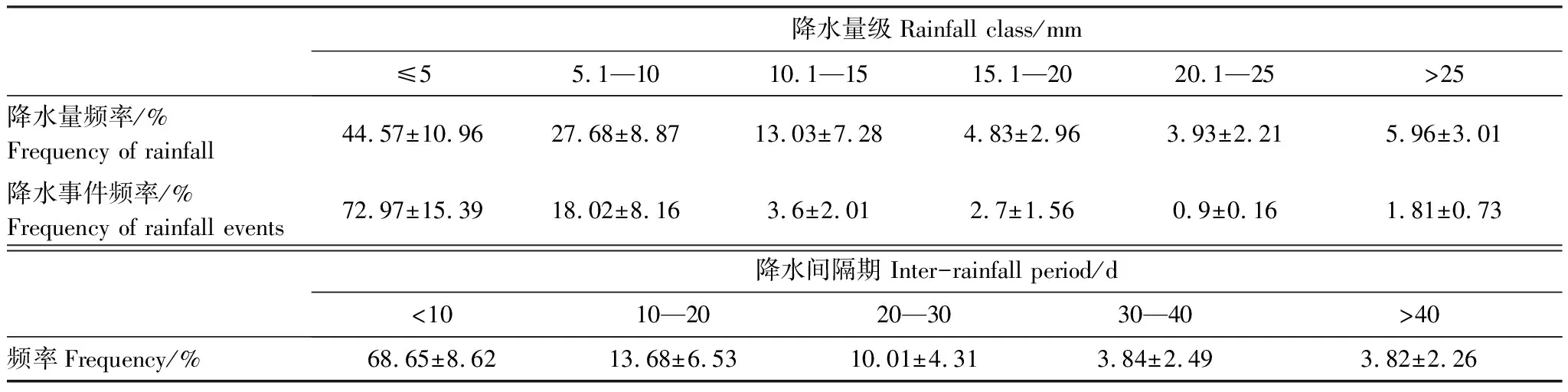
表1 研究区40年降水量、降水事件、降水间隔期频率分布
1.4 数据分析
采用SPSS 21.0进行单因素方差分析和Duncan′s多重比较分析同一植物不同胁迫水平各个参数的差异显著性(P<0.05),用相关性分析方法分析植物各生理之间的相关性。利用Origin 2018作图软件完成图形绘制。
2 结果与分析
2.1 3种一年生草本植物对干旱胁迫的生理响应
在不同干旱胁迫下叶片渗透调节物质(脯氨酸、可溶性糖和可溶性蛋白)差异显著(P<0.05),脯氨酸、可溶性糖含量基本趋势为随着干旱胁迫天数的增加而增加,可溶性蛋白含量随干旱胁迫天数增加呈先增大后减小:干旱胁迫5d时,叶片脯氨酸、可溶性糖和可溶性蛋白含量均上升,脯氨酸含量上升幅度最大(190.81%、184.62%、51.80%);干旱胁迫10d时,与对照相比可溶性蛋白含量增幅(652.80%、1045.85%、1333.48%)较大,并达到峰值;干旱胁迫15d时,叶片可溶性蛋白含量有所下降,可溶性糖增幅(584.49%、197.12%、635.47%)较大(图3)。

图3 雾冰藜、虎尾草和狗尾草渗透调节物质对干旱胁迫的响应Fig.3 Response of osmoregulation substances in Bassia dasyphylla,Chloris virgata and Setaria viridis to drought stress不同黑色字母代表雾冰藜不同干旱胁迫处理差异显著(P<0.05),相同黑色字母代表雾冰藜不同干旱胁迫处理差异不显著(P>0.05);不同红色字母代表虎尾草不同干旱胁迫处理差异显著(P<0.05),相同红色字母代表虎尾草不同干旱胁迫处理差异不显著(P>0.05);不同蓝色字母代表狗尾草不同干旱胁迫处理差异显著(P<0.05),相同蓝色字母代表狗尾草不同干旱胁迫处理差异不显著(P>0.05);CK:对照; DS:重度干旱胁迫后复水
不同干旱胁迫下叶片丙二醛含量差异显著(P<0.05),随着干旱胁迫天数增加呈上升趋势:干旱胁迫15d时,叶片丙二醛含量增大到最高值(7.41μ mol/g、14.16μ mol/g、10.34μ mol/g)(图4)。
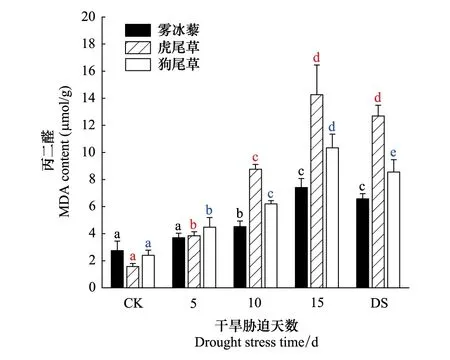
图4 雾冰藜、虎尾草和狗尾草丙二醛含量对干旱胁迫的响应 Fig.4 Response of Malondialdehyde (MDA) contents in Bassia dasyphylla,Chloris virgata and Setaria viridis to drought stress
在不同干旱胁迫下叶片叶绿素含量差异显著 (P<0.05),随着干旱胁迫天数的增加,呈下降趋势:在对照组,叶绿素含量分别为0.83mg/g、0.48mg/g和0.58mg/g;干旱胁迫15d时,叶片叶绿素含量降至最低值,与对照相比分别下降了46.21%、39.69%和33.34%(图5)。

图5 雾冰藜、虎尾草和狗尾草叶绿素含量对干旱胁迫的响应 Fig.5 Response of chlorophyll contents in Bassia dasyphylla,Chloris virgata and Setaria viridis to drought stress
在不同干旱胁迫下根系活力差异显著(P<0.05),呈先上升后下降趋势:在干旱胁迫5d时,各植物根系活力有所上升并达到最大值,分别升高31.77%、17.89%和20.91%;随后逐渐降低,干旱胁迫15d时,根系活力下降至最低值(图6)。

图6 雾冰藜、虎尾草和狗尾草根系活力对干旱胁迫的响应 Fig.6 Response of root activity in Bassia dasyphylla,Chloris virgataand Setaria viridis to drought stress
15d干旱胁迫复水后,叶片脯氨酸含量、可溶性糖含量和丙二醛含量均有所下降,叶绿素含量和根系活力均有所上升。
2.2 3种一年生草本植物对干旱胁迫的形态响应
随着干旱胁迫天数增加,3种植物根长伸长;虎尾草和狗尾草的茎长呈现出下降趋势,雾冰藜茎长呈上升趋势;雾冰藜根长与茎长度的比呈现下降趋势,虎尾草和狗尾草根长与茎长度的比呈现上升趋势。干旱胁迫15d时,虎尾草、狗尾草茎长分别减少了25.25%和26.59%,雾冰藜茎长上升了44.61%,3种植物根长较对照处理下伸长了18.86%、26.38%和21.74%(图7)。
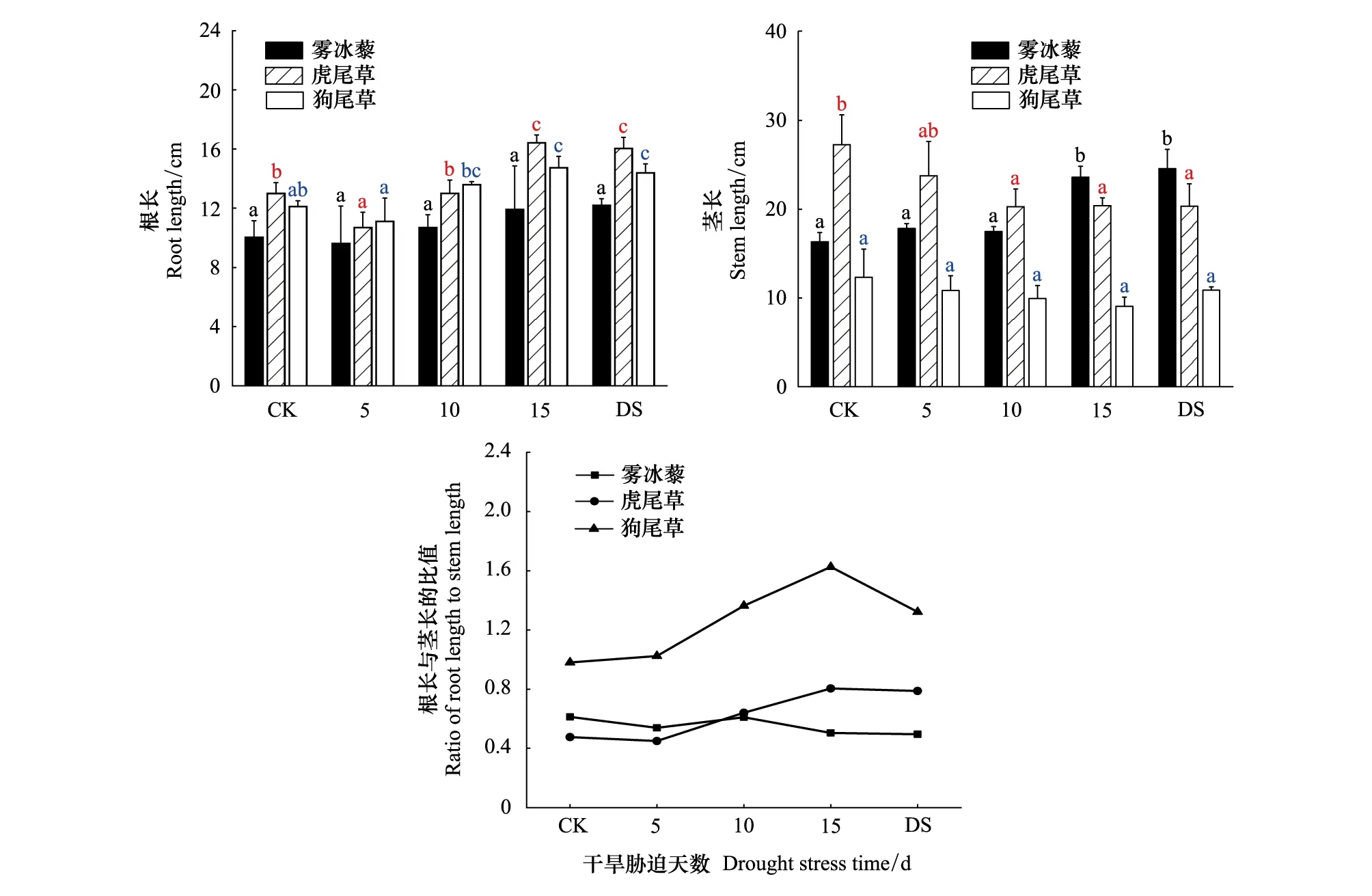
图7 雾冰藜、虎尾草和狗尾草生长对干旱胁迫的响应Fig.7 Response of the growing in Bassia dasyphylla,Chloris virgataand Setaria viridis to drought stress
2.3 3种一年生草本植物对干旱胁迫的繁殖响应
在不同干旱胁迫下,结种数和百粒重均差异显著(P<0.05):随着干旱胁迫天数的增加,结种数和百粒重总体呈下降趋势。干旱胁迫15d时,结种数均降为最小值,与对照相比,分别降低了67.82%、45.82%和46.97%,百粒重降幅为52.53%、61.52%和55.49%(图8)。
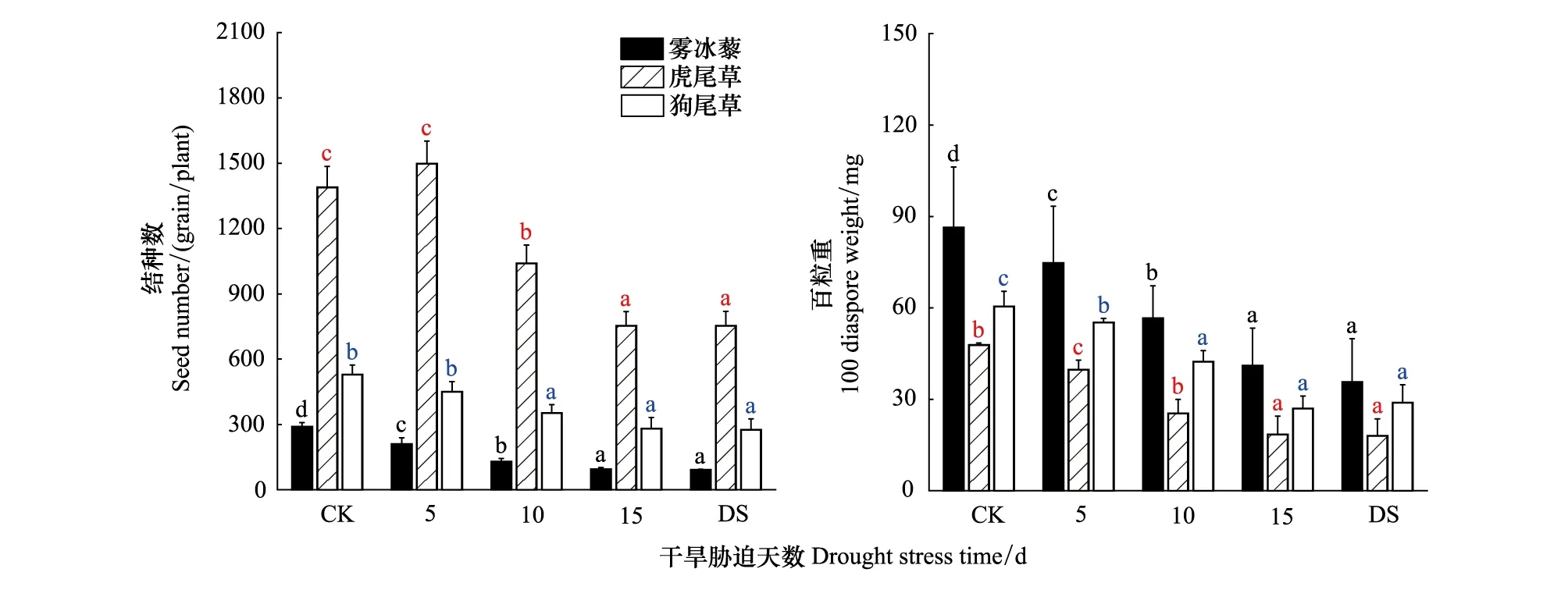
图8 雾冰藜、虎尾草和狗尾草繁殖体对干旱胁迫的响应Fig.8 Response of propagules in Bassia dasyphylla,Chloris virgata and Setaria viridis to drought stress
3 讨论
3.1 3种一年生草本植物对干旱胁迫的生理响应
本研究发现,当3种一年生草本植物受到干旱胁迫时,叶片内会产生大量的渗透调节物质(脯氨酸、可溶性蛋白和可溶性糖),轻度干旱脯氨酸含量增加190.81%、184.62%、51.80%,中度干旱可溶性蛋白含量增加652.80%、1045.85%、1333.48%;而重度干旱复水后,干旱胁迫被解除,3种一年生草本植物的脯氨酸和可溶性糖含量向正常水平恢复,这说明这3种一年生草本植物在叶片渗透调节方面对复水表现出明显的响应,对于重度干旱胁迫的损伤仍具有较高的修复能力。很多研究表明,植物受到干旱胁迫时,叶片的细胞通过吸收无机离子(例如,Na+、Ca2+)[31]或产生有机质溶质[32- 34]维持渗透势,保持细胞继续吸水,维持叶片细胞组织具有一定的持水力或免于脱水,从而对干旱胁迫起到了缓冲保护的作用。3种一年生草本受到干旱胁迫时,在植物叶片内会主动积累脯氨酸、可溶性蛋白和可溶性糖来提高提高渗透调节能力,维持细胞内外渗透平衡,保证细胞的保水能力,增强其抗旱性,尤其是脯氨酸,是一年生草本植物对干旱胁迫反应最为敏感的一种渗透调节物质[35]。在本研究区,年降水量只有大约100mm,<10d降水间隔期占比率(68%),降水发生时间和持续时间多变,荒漠一年生草本植物在荒漠区长期处于干旱胁迫状态,增加叶片的脯氨酸、可溶性蛋白和可溶性糖含量,维持稳定渗透势,是荒漠一年生草本应对荒漠环境中干旱的关键生理适应策略。
同时,本研究发现3种一年生草本植物叶片内丙二醛含量也会随着干旱程度增加而增加(169.13%、804.65%、330.69%);3种一年生草本植物叶片内叶绿素含量也会随着干旱程度增加而减少(下降46.21%、39.69%和33.34%)。一般认为,当干旱对植物的胁迫程度较轻时,植物体内丙二醛在较低水平,一旦超过植物的耐受极限,导致脂质过氧化产物丙二醛的大量产生[36]。丙二醛含量上升会加速叶绿素的分解,从而抑制植物光合作用和个体生长[37]。在本研究中,轻度干旱胁迫下,丙二醛与渗透调节物质、叶绿素含量关系不显著,而随着干旱胁迫程度增加,丙二醛与渗透调节物质、叶绿素含量之间呈显著负相关关系(表2),这是由于轻度干旱时一年生草本植物叶片渗透调节物质上升,叶片丙二醛含量维持在较低水平,叶绿素含量增加;而中度和重度干旱胁迫时,丙二醛含量逐渐增加,丙二醛具有细胞毒性,与膜结构上的蛋白质结合,使蛋白质含量下降[21],导致一年生草本植物的渗透调节作用不显著,同时叶绿素分解加速,从而使植物生长受到抑制。
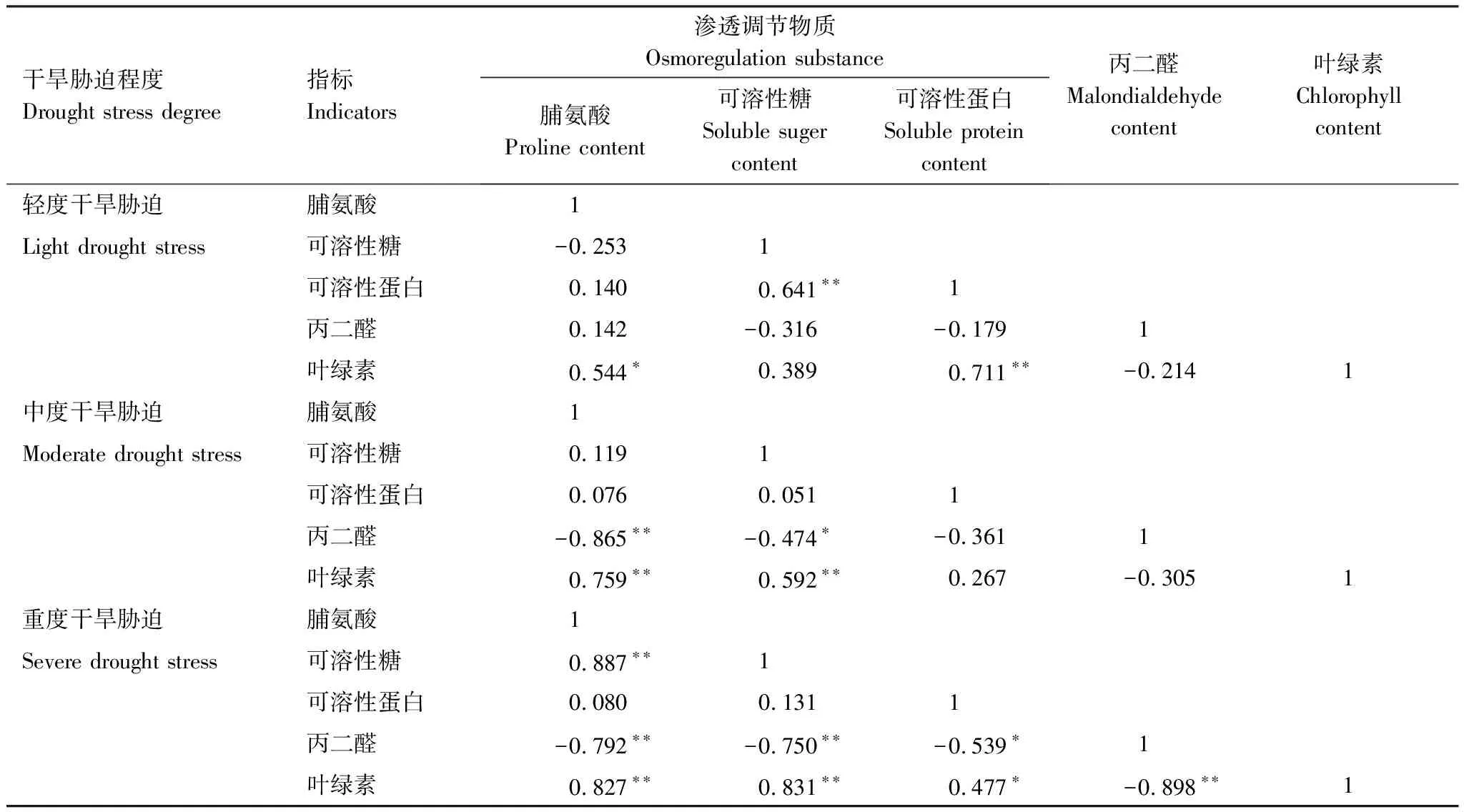
表2 干旱胁迫下3种一年生草本植物生理指标间的相关性分析
本研究还发现,在轻度干旱条件下,3种一年生草本植物根系活力为最大值,而随着干旱程度增加,根系活力逐渐降低。根系活力影响水分吸收,活力越强,越有益于水分吸收[38]。一年生草本植物在轻度干旱胁迫时,除了进行渗透调节以及光合作用调节,还会通过增加根系活力,保证根系从土壤中吸收更多水分,以缓解干旱对生长的抑制,从而表现出适应干旱环境的能力。
3.2 3种一年生草本植物对干旱胁迫的形态响应
本研究发现,当降水的时间间隔增大,干旱程度加剧时,3种一年生草本植物还可以通过个体形态调节进行适应,尤其在重度干旱胁迫时,茎长下降明显,植物增加地下根系投入,通过延伸主根系长度,增加根长来适应严重干旱。在荒漠生态系统,许多旱生植物对资源限制都可以做出类似的个体形态反应,根系会向较深土层延伸,在深层土壤吸收土壤水分[22- 23,39]。而对于浅根系的一年生草本植物,根系对降水和浅层土壤水分变化极为敏感,最先感知干旱胁迫,因而,在干旱胁迫时,根系最先反应,这是一种典型的“开源”策略[40]。
3.3 3种一年生草本植物对干旱胁迫的繁殖响应
本研究还发现,轻度干旱和中度干旱时,一年生植物最大限度地提高繁殖输出,百粒重及结种数均维持在较高水平,而重度干旱胁迫时,一年生草本植物株的地上部生长受到抑制作用,直接导致了百粒重和结种数量下降,结种数分别下降67.82%、45.82%和46.97%,百粒重分别下降52.53%、61.52%和55.49%。有研究表明,植物受到严重干旱胁迫时,繁殖分配低即可保证其存活,如果提高繁殖分配,则会危及一年生草本植物的生存。资源利用学说认为,生境中的可利用资源是影响植物种群繁殖分配比例的重要因素,当资源不足时,植物会将更多的资源分配给营养结构以提高资源的获取能力[41]。有研究表明,荒漠植物权衡种子数量和质量资源分配,是荒漠植物适应干旱环境的一种机制[42]。许多研究都预测干旱荒漠地区未来的极端干旱和极端降雨事件将更频繁[43-44],使降水间隔期更大,干旱荒漠地区植物可能经受更为严重干旱胁迫。在全球气候变化的背景下,一年生草本植物作为机会主义者,比多年生草本植物可能更具有优势,一年生草本植物在应对干旱时具有更灵活的生长和繁殖适应策略,在荒漠生态系统中作用将更加明显。
4 结论
本研究发现轻度和中度干旱胁迫时一年生草本植物主要通过生理调整:增加叶片脯氨酸含量、可溶性蛋白含量和可溶性糖含量维持叶片渗透压,提高保水能力,并增加叶绿素含量,使丙二醛含量维持在较低水平,从而使植物维持正常体内生长环境;同时,轻度干旱时根系活力增强,有效促进根系水分吸收与生长。由于生理调整作用,轻度和中度干旱时百粒重及结种数均维持在较高水平。而在重度干旱胁迫下,叶片丙二醛含量迅速增加达到最高值,加速叶绿素分解使可溶性蛋白含量下降,渗透调节作用达到了极限,从而抑制植物有效生长,植物开始启动个体形态响应(增加根系投入,减少地上茎干投入),根系延伸反应出一年生草本植物适应干旱逆境的一种“开源”策略。同时,重度干旱胁迫时,一年生草本植物植物地上部生长受到抑制,从而导致结种数和百粒重下降。
猜你喜欢
小哥白尼(趣味科学)(2022年5期)2022-08-15
作文周刊·小学一年级版(2020年40期)2020-10-19
绿色中国(2019年14期)2019-11-26
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
儿童故事画报·智力大王(2016年7期)2017-02-08
文史春秋(2016年3期)2016-12-01
小学生作文选刊·低年级版(2016年6期)2016-06-24
食品与生活(2014年2期)2014-03-04
中国火炬(2012年9期)2012-07-25
