
基于遗传图谱的班章古茶树遗传多样性分析
2021-07-29 01:48:54谭思敏高彬林胤
广东茶业 2021年3期
谭思敏 高彬 林胤
(1东莞中国科学院云计算产业技术创新与育成中心 广东省东莞市 523808)
(2东莞市春福润茶叶有限公司 广东 东莞 523570)
(3东莞中国科学院云计算产业技术创新与育成中心 广东东莞 523808)
一 引言
植物多样性作为地球上较为宝贵的财富,是人类的发展与生长的物质源泉。遗传多样性的丰富程度影响着品种培育与改良的潜力。中国作为茶的原产地,具有漫长的种茶历史,对于古茶树的种质资源探究则成为古茶树培育的关键。为此,不少研究学者针对班章古茶树的生长特征进行班章古茶树遗传多样性分析,促进茶树的种群生长与繁殖[1]。
为维护班章古茶树的生长多样性,提高古茶树的生长质量,本文对其遗传多样性进行研究分析,探索适宜班章古茶树生长的遗传规律,由此保持古茶树的可持续发展。目前的古茶树遗传多样性研究针对古茶树的生长特点分析不同古茶树的染色体信息,并根据获取的染色体信息分类,利用标记法对染色体信息标记,同时调整标记的信息数据,聚类分析不同古茶树的种群,并研究其内部种源关系。初步构建不同性状间的线性关系,记录线性关系生成条件,设置相应的线性规律表,对比线性规律表调节整体古茶树生长态势与生长结构,进而提高古茶树的生长质量。但传统分析研究对于古茶树的环境特征分析力度较小,无法满足古茶树遗传多样性的整体分析需求,为此,针对上述问题,本文提出一种新式基于遗传图谱的班章古茶树遗传多样性分析对以上问题进行分析与解决[2]。
本文分析方法利用不同的研究试剂对古茶树的生长特征进行数据分析,同时调节古茶树之间的遗传特征关系,构建遗传图谱信息,将染色体信息录入图谱中,并保留试剂样本状态,确保分析数据的完整程度,实现对班章古茶树的遗传多样性分析,具有较强的操作性,能够更好的推动班章古茶树的发展与生长[3]。
二 材料与方法
1.试剂
班章古茶树树种、丙烯酰胺、二甲双丙烯酰胺、TEMED、Taq酶。
2.提取方法
选用100g的新鲜茶叶,将茶叶放入设置好的容器中,等待后续容器研究处理。进行氮液研磨,同时加入1ml预热的分离溶液,将溶液提取物与茶叶物质相结合,同时充分蒸馏过多的提取物水分。65℃水浴15min,同时保证茶叶内部的物质处于平衡状态,分解茶叶机制,将有机混合物分离,加入等体积的氯仿,静置5min,离心10min,转移上清至新试管中,将试管中的溶液充分分解,并选用80%乙醇洗涤沉淀两次,最后静置,以免溶液重悬[4]。
将放置的溶液放入新备容器中,37℃水浴5min,重复抽提内部溶液,沉淀内部溶液,并将内部溶液与茶叶溶液分离,获取新式溶液,由此实现对茶叶内部溶液的提取。
3.古茶树分析设备调整
在进行茶树研究的同时进行内部分析板的设置,构建茶树茶叶叶分析塑料板、栓钉以及茶叶装置连接板有限元模型,连接不同组建的内部结构,并调整结构信息的操作方向,将数据模型集中控制在操控范围之内[5]。组建模型各部分元件,在完成组建后,构建设备装置材料库,破除固有的设备材料模式,并添加新的材料属性于材料库中,茶树茶叶分析塑料板材料在初始行为输出中的模型数量设置为1560MPa,其泊松比为0.53。构建的材料板截面采用实体板质,根据不同的材料特性进行元件截面指定制造,构建设备结构图:

图1设备结构图
在试验过程中,为更加便利地计算出所需的茶叶遗传性特征信息,固定约束外层分析板的四个板脚,同时加强对分析板的数据看护力度,压制其四周的数据位移,并将位移数据转化为内部节点操控数据,及时改变不同的分析板形式,并存储茶叶数据遗传信息板此时的数据状况,将此些数据集中收集至内部存储空间中实现内部管理操作[6]。
利用遗传分析板栓钉与土层之间的相互作用压制栓钉的位移方向,并转动栓钉的位移位置,构建有限元模型边界条件收录空间。创造分析板承载负荷基础数据库,将分析板承载负荷信息全部录入数据信息库中,及时检验数据库中的数据存储状态,时刻保证古茶树在进行遗传信息调节的过程中能够保持调节安全性。加强对古茶树的染色体处理,分析古茶树的茶谱间距,同时将茶谱间距进行适当调整[7]。控制茶谱边距处于分析板内部,根据茶树分析板载荷类型划分为不同的遗传数据分析空间,优化遗传分析板的几何截面,将其载荷方向控制在古茶树树种调节板的垂直方向上,并加强对调节板的管理力度,防止茶树遗传数据的外泄。
分析板内部压强初始数值设置为0.02MPa,在试验初始阶段,将茶树茶谱边距板的压力承载梯度增加0.02MPa,并随着试验时间的增加而逐渐增加该梯度数值,分析板的压力承载梯度增加0.01MPa[8]。在实现各组件重组后,在中心网络划分模块中将有限元模型数据的划分为不同的数据模型空间,规定网络模型的具体尺寸,并按照尺寸大小匹配操作的数据模型,管理此时的茶树分析板状态,在中心网络操作阶段,执行单元格式选择命令,及时转变分析板形状,并设置网络划分格式标准,按照标准执行试验操作,选取不同的单元形状,实现初始试验操作,构建古茶树内部存储板结构图:

图2古茶树内部存储版结构图
在整体结构与系统模型空间的变化中,根据此时的班章古茶树内部茶树状态,设置蠕动形变测试器,记录垫片在茶树遗传特征分析情况下的蠕动形变程度,并对模型的整体应变能力进行分析,判断其是否符合模拟器中的本模型应具备的条件,并在结构固定的同时选用C173单元进行预计截面采集,由于模型的形变自由度较大,接触单元中选择弹性形变较好的接触编辑器,在分析结构与模型预计截面结构不符时,接触编辑器将自动改变模型形态,提高垫片的弹性形变性能,在目标与模型相接触的同时进行时由系统的自由对接,并完成相应反应动作[9]。
由于班章古茶树树种具有较强的预测复杂性,需要对分析设备进行整体调整,调节设备内部元件结构,在合金垫片与法兰的公共节点上添加节点平滑装置,利用装置的磨合性将整体系统进行节点磨合处理,使模型中的垫片与法兰相吻合,在运用过程中,要适当忽略垫片与法兰接触时产生的物理摩擦力,并进行相应的模拟系统操作,将此时的垫片应力与法兰承受力记录,作为应力分析数据的一部分而存在,此外,对合金垫片的材料非线性形变进行分析处理[10]。
螺栓法兰合金垫片的材料具备较多的特性,其各元件材料主要分为两个特性:线性形变弹性与非线性形变弹性。本文设置法兰接触系统中的各螺栓以及垫片的材料均为线性形变弹性材料,并设置其法兰的弹性模量为2.21,泊松比为0.3μ,其屈服强度为522σ。
由此实现对班章古茶树的分析装备的设置,同时将配备好的溶液与设置的装备相结合,进行后续遗传图谱构建操作[11]。
4.遗传图谱构建
针对遗传图谱构建的特点,本文选择相应的亲本数据,将DNA形态较为丰富的古茶树样本作为亲本,增加内部标记数量。进一步纯化亲本特征,避免古茶树遗传特征研究过程中产生数据分析失误。着重考虑树种间的杂交繁育情况,加大对班章茶树的管理力度,防止繁育失败现象的产生[12]。在实现亲本选配后,筛选选择的亲本,将不符合遗传性分析的亲本数据清除,保留与分析特征相似度较高的数据信息。加强班章古茶树的茶谱判断操作力度,同时根据茶谱的茶树收集特征分析不同区域位置的班章茶树生长特点[13]。标记班章古茶树的树种点,设置5个树种查找区域点。在1至3区域点范围内,班章古茶树平均有效等位基因数为1.8135。若变化范围符合基础实验需求,则可进行下一步骤的茶树遗传特征分配研究,同时对其内部信息进行处理,构建处理图:
图3内部处理图
由于连锁图谱为单交群体,在对班章古茶树群体进行图谱构建的过程中需调整构建的阶段特征,分析不同阶段内产生的图谱分析状况,并及时转化图谱的茶树遗传结构,掌控茶树茶谱内部繁殖材料信息[14]。标记单株繁殖特征,并分辨不同标记符号下的标记类型,保持茶树的遗传结构特征,确保在特征分析过程中茶树的基础遗传特征不受影响。不同的培育方法将对茶树的遗传特征产生不同程度的影响,在进行实验的同时针对目标权衡情况选择合适的班章古茶树树种大小,构建图谱群体集体操作机制,将古茶树的操作机制与其相结合,并对比不同机制条件下古茶树的配比情况,实现对古茶树的内部遗传多样性分析调整操作,结束研究操作部分[15]。
三 结果与分析
根据上述研究数据进行实验研究,并获取相应的班章古茶树遗传多样性特征。
选用3对设计的引物组合对班章古茶树品种资源进行结构性扩增,同时分离出240条图像清晰的操作条带,检验条带内部的多态性位点,检测条带点数量为300多个,每个引物组合连接的多态性位点能够连接不同的古茶树特征点状图谱,在进行图谱连接的同时不断分离图谱的内部距离,并按照调整的距离信息缓冲古茶树遗传信息的多样性信号,结束图谱差异内部的调节模式,在获取的每组引物组合中,揭示引物分子的基因状况,同时将现实的基因对比图像扩充至操作界面中,按照操作流程配置界面数据信息,并加强对内部数据界面信息的管理力度,构建不同引物组合检测效率对比表:
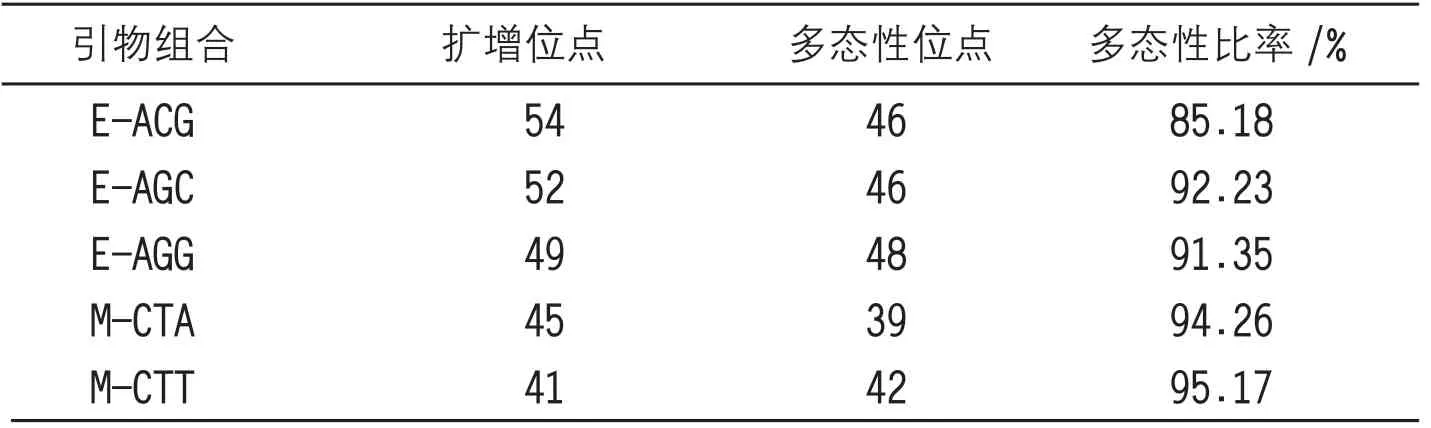
表1引物组合检测效率对比表
在对引物组合检测效率进行对比后,对比不同区域环境条件下古茶树的相似性系数,并构建相似性系数表如下所示:

表2相似性系数表
根据上表可知,在不同的区域环境影响下,班章古茶树的遗传特点随着环境的变化而变化。不同种群间的差异较大,将会对后期班章古茶树的生长状况产生影响。为此,在进行实验调节的后期阶段,需调整班章古茶树的环境区域条件,并结合不同区域条件特征,执行相应的环境区域缓解指令,并对不同区域条件下的环境指令做出空间反应,缓和区域条件与古茶树遗传多样性特征间的关系,加强对班章古茶树的树种分析力度,将属于相同区域的树种信息记录于相同的空间中,等待空间的后续研究与处理,同时处理其内部遗传链信息,构建传输图:

图4遗传链传输图
综上所述,依据不同的引物组合顺序进行数据排列,完整分析班章古茶树的遗传多样性特征,由此分析不同区域环境下对古茶树遗传多样性系数的影响得出,班章古茶树的遗传多样性与所处区域环境相关度较高。
四 讨论
在实现上述研究后,与前人的研究结果进行对比,本文分析方法对于班章古茶树的相似性调查程度较深,获取的茶树生长特征较为完整,针对班章古茶树的生长习性研究结论相同,皆认为班章古茶树的生长特征受生长环境的影响,进而影响其内部遗传特征。在进行内部特征试剂检验的过程中,加强了对古茶树的染色体分析,获取了不同茶树树种之间的遗传相似性差异受自体染色体的变化而变化的结论,完善了古茶树遗传多样性研究原理,能够更好地为古茶树的遗传多样性研究提供良好的理论基础,为验证本文研究的有效性,将本文分析结果与传统研究分析结果进行实验对比,得出如下实验对比图:
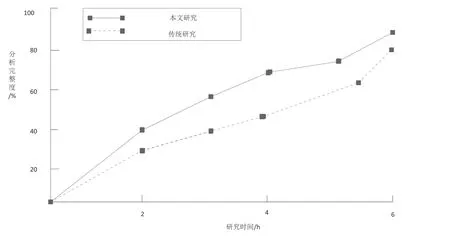
图5xin’x实验对比图
根据以上图示可以看出,在研究时间为2小时时,本文基于遗传图谱的班章古茶树遗传多样性分析的分析完整度为40%,在研究时间为4小时时,本文基于遗传图谱的班章古茶树遗传多样性分析的分析完整度为65%,在研究时间为6小时时,传统班章古茶树遗传多样性分析的分析完整度为92%。
造成此种差异的主要原因在于本文研究针对班章古茶树的内部特征进行数据分析,同时加强对其内部染色体信息的管理,并加大管理强度,在获取相关的染色体研究数据的同时分析不同区域环境条件下的班章古茶树的遗传特性,有效缓解不同区域对古茶树研究的压力,并增强对班章古茶树的管理性能,有效获取相关的古茶树遗传多样性信息,分析完整度较高,能够更好地为后续研究提供强有力的数据支撑。
五 结论
根据上述分析研究,得出以下结论:
(1)在进行班章古茶树遗传多样性分析的同时应注意对古茶树周围环境区域的信息分析,减少周边区域环境对分析结果的影响。选用不同的聚类方法聚合不同的古茶树信息,有效避免因数据收集完整度不高造成的分析精准度过低的状况的产生。
(2)班章古茶树内部遗传相似性较高,可被分为单独的一类进行培育,在茶树培育的过程中需选择特定培育环境,提升培育的质量与茶树的生长优势,进而促进班章古茶树的有效生长。
(3)班章古茶树作为普洱茶树种的一种,其生长特点应区别于一般茶树,在生长观察阶段需注重对古茶树本身的特征分析力度,及时缓解古茶树的自身生长压力,并匹配相关的生长试剂,提升班章古茶树的遗传生长效率。
猜你喜欢
电站辅机(2021年2期)2021-08-19 00:33:16
少先队活动(2020年12期)2021-01-14 01:47:40
乡村地理(2018年2期)2018-09-19 06:44:00
中成药(2017年3期)2017-05-17 06:09:01
领导科学论坛(2016年9期)2016-06-05 14:59:58
湖南农业(2016年3期)2016-06-05 09:37:36
音乐天地(音乐创作版)(2016年11期)2016-02-05 01:10:59
金属加工(热加工)(2015年11期)2015-11-16 06:04:26
橡胶工业(2015年8期)2015-07-29 09:22:46
锻压装备与制造技术(2015年2期)2015-06-26 09:00:24
