
鳜不同亚型瘦素受体基因克隆与表达分析
2021-07-26 13:53:24魏君冉梁旭方徐晶蔡文静
华中农业大学学报(自然科学版) 2021年4期
魏君冉,梁旭方,徐晶,蔡文静
华中农业大学水产学院/华中农业大学鳜鱼研究中心/农业农村部淡水生物繁育重点实验室/湖北省名优鱼育种与健康养殖工程技术研究中心,武汉 430070
瘦素(leptin)在1994年首次在小鼠(Musmusculus)中被发现,并描述为一种由肥胖基因(obese gene,ob)编码的、由脂肪细胞分泌的多肽激素[1]。在哺乳动物中,leptin由脂肪组织合成并分泌进入血液,通过血脑屏障后到达下丘脑并与瘦素受体(leptin receptor,lepR)结合,导致食欲降低和脂质代谢增加[2]。继leptin被发现后,Tartaglia等[3]随即在小鼠中克隆出lepR基因。已有的研究表明,哺乳动物只有一个lepR基因的旁系同源物,lepRmRNA通过3′端的可变剪切形成6种不同的亚型,这6种不同的亚型可分为长型(lepRb)、短型(LepRa、LepRc、LepRd、LepRf)、分泌型(lepRe)3类[4]。自首次发现小鼠的lepR基因十多年后[3],Wong等[5]首次在海洋青鳉(Oryziasmelastigma)中发现鱼类的lepr基因。不同于哺乳动物的是,在大多数鱼类中只克隆出长型lepr,如日本青鳉(Oryziaslatipes)[6]、斑马鱼(Daniorerio)[7]和斜带石斑鱼(Epinepheluscoioides)[8]等。在少数鱼类中通过cDNA末端快速扩增技术(rapid amplification of cDNA,RACE)克隆出多个由leprmRNA 3′端可变剪切产生的受体亚型,如虹鳟 (Oncorhynchusmykiss)[9],黑鲫(Carassiuscarassius)[10]和欧洲鲈 (Dicentrarchuslabrax)[11]。
鱼类长型受体具备哺乳动物lepR高度保守的结构,短型受体亚型的胞内结构、氨基酸组成及序列长度均存在差异,分泌型受体通常缺乏胞内区和跨膜区。与哺乳动物一致,鱼类的lepr只有长型受体亚型具备完全的重要功能域。在虹鳟[9]、大西洋鲑(Salmosalar)[12]及黑鲫[10]中的研究发现短型受体亚型可能与能量稳态有关。对于鱼类可溶性leptin受体的研究发现其与leptin-a的结合能力高于leptin-b,随着对多种鱼类的lepr基因的鉴定和研究的深入,发现lepr及其leptin结合域(leptin binding domain,LBD)在鱼类进化过程中相对保守[7-8,11,13]。
在哺乳动物及鱼类中,leptin通过其专一性受体结合传导抑食欲信号[14-15]。使用哺乳动物同源重组的leptin蛋白处理金鱼可导致其摄食量下降[16],与在哺乳动物中观察到的结果一致,但类似的处理并不能影响银大马哈鱼(Oncorhynchuskisutch)[17]、鲶(Silurusasotus)[18]和绿海鲂(Lepomiscyanellus)[19]的摄食行为。这些不同的结果可能是鱼类和哺乳动物的leptin及lepr氨基酸序列差异较大引起的。因此,研究鱼类leptin及lepr的结构和功能对于研究鱼类摄食行为来说至关重要。鳜是我国特有的名贵淡水鱼,其食性奇特,自开口起终身以活饵为食,但经长期驯化才可食人工饲料。本课题组在前期研究氨基酸、脑肠肽(PYY)对鳜摄食调控等[20-21]工作的基础上,利用3′ RACE 技术,克隆鳜由mRNA的3′端可变剪切4个不同的lepr受体亚型,检测长型受体亚型在鳜组织中的表达情况,以及注射鳜同源重组leptin蛋白对其在脑中表达的影响,以期为后续深入研究鳜不同亚型的lepr在摄食调控、生理代谢中的功能及leptin-a和leptin-b的基因功能提供理论依据。
1 材料与方法
1.1 试验鱼及饲养条件
试验鱼来自华中农业大学鳜鱼研究中心,于农业农村部鳜鱼育种创新基地的循环水养殖系统中暂养14 d,水温为(25±1) ℃,pH为7.53~7.70,氨氮、亚硝酸氮、硝酸氮质量浓度均低于0.1 mg/L,每天09:00和17:00各饱食投喂鲜活麦鲮(Cirrhinusmrigala)1次。
1.2 鳜leptin A和leptin B同源重组蛋白的表达、纯化及腹腔注射
鳜leptin A和leptin B同源重组蛋白的表达和纯化方法参照Yuan等[22]的方法进行。暂养结束后,随机选取9尾健康、体表无伤、质量为(230±16) g的鳜,饥饿24 h后,使用MS-222麻醉后称取质量。采用腹腔注射的方法,试验组注射1 000 ng/g的溶解于DPBS的leptin A或leptin B同源重组蛋白,对照组注射等量的DPBS溶液,每组注射12尾试验鱼。
1.3 组织的获取及RNA的提取
取样前,手术刀、手术剪及1.5 mL离心管均于37 ℃下质量分数为0.1%的焦碳酸乙二酯(DEPC)浸泡12 h,随后使用高压灭菌锅于120 ℃下处理30 min以去除DEPC,烘干后密封保存备用。鳜经MS-222麻醉后,解剖提取3尾试验鱼的垂体组织,所获得的垂体组织置于1.5 mL离心管并立即冻存于液氮中,取样结束后冻存于-80 ℃,用于后续lepr的3′- RACE克隆。取6尾试验鱼的端脑、中脑、小脑、垂体、下丘脑、延脑、肝脏、肠、肾、脂肪组织、鳃、脾和心组织置于1.5 mL离心管并立即冻存于液氮中,用于后续检测lepr基因的组织表达。分别在注射鳜leptins同源重组蛋白2 h及4 h后取每组鳜的脑组织,置于1.5 mL离心管并立即冻存于液氮中。
将用于lepr的3′- RACE克隆的垂体组织混合,其余组织取适量放入干净的1.5 mL离心管中,向装有组织的离心管中加入1 mL TRIzol 试剂,并装载入-20 ℃预冷的组织破碎仪夹板中,充分匀浆后按照有机溶剂抽提RNA法提取组织RNA。RNA经1.2%琼脂糖凝胶电泳检测完整性后用于后续试验。
1.4 3′ RACE 引物设计及lepr cDNA全长克隆
从鳜基因组数据库中调取lepr基因序列,根据TaKaRa公司的3′-Full RACE Core Set with PrimeScriptTMRTase试剂盒的说明和lepr基因的核心序列设计外引物Lepr- Outer,以及内引物Lepr-Inner,用于扩增lepr的cDNA全长序列。用于3′RACE克隆的引物如表1所示。
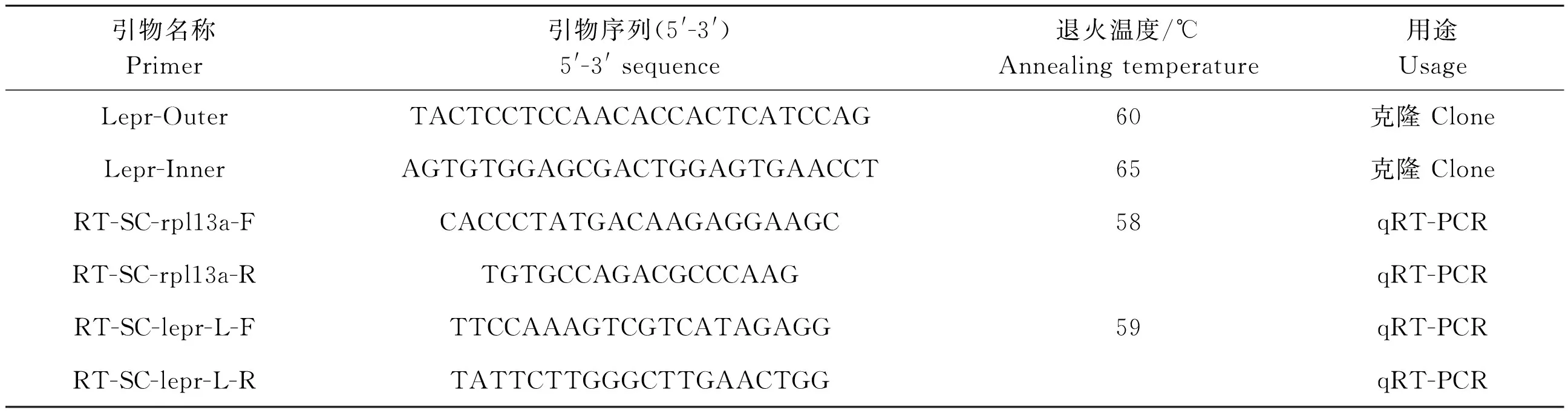
表1 本研究中所使用的克隆及定量引物 Table 1 Primers used for clone and qRT-PCR in this study
使用TaKaRa公司的3′-Full RACE Core Setver2.0以及DNA聚合酶(TaKaRa,日本)合成鳜lepr基因cDNA全长序列。根据试剂盒的使用说明操作,获得PCR产物,经1.2%琼脂糖凝胶电泳检测无误后用于连接、转化。
1.5 lepr基因3′RACE克隆产物的连接、转化及测序
PCR产物经1.5%的琼脂糖凝胶电泳分离,切胶并使用Omega公司的EZNA Gel Extraction Kit试剂盒进行纯化回收,并连接入T克隆载体(北京全式金公司,pEASY©-T1 Cloning Kit)。将克隆载体转入DH5α感受态细胞(北京全式金公司),用不含有氨苄的LB培养液在37 ℃的摇床中培育1 h后,吸取200 μL菌液涂于含有氨苄的LB固体培养基平板上,37 ℃过夜培养。挑选阳性克隆送武汉生工生物工程有限公司测序。
1.6 不同亚型的lepr基因的生物信息学分析
使用NCBI网站的BLAST及ORF finder工具将测序结果与鳜基因组调取的lepr序列进行比对,并预测其氨基酸序列。分别使用在线软件Signal P 4.1 Server (http://www.cbs.dtu.dk/services/Signal P/),TMpred Server (https://embnet.vital-it.ch/software/TMPRED_form.html)及SMART:Main page (http://smart.embl-heidelberg.de/)预测测序所得的lepr序列的信号肽、跨膜区和结构域。从GenBank数据库(https://www.ncbi.nlm.nih.gov/genbank/)获取以下15个物种的lepr氨基酸序列:人(Homosapiens)、小鼠(Musmusculus)、大鼠(Rattusnorvegicus)、红鳍东方鲀(Takifugurubripes)、爪蟾(Xenopustropicalis)、大黄鱼(Larimichthyscrocea)、尖吻鲈(Latescalcarifer)、欧洲鲈、罗非鱼(Dreochromisniloticus)、斜带石斑鱼(Epinepheluscoioides)、虹鳟、大西洋鲑、日本青鳉、斑马鱼、黑鲫,利用Bioedit软件对上述氨基酸序列进行多重比对,并使用MEGA 6.0软件构建上述物种lepr蛋白质的系统进化树(bootstrap=1 000)。
1.7 qRT-PCR
通过qRT-PCR技术检测DNA的表达水平。20 μL反应体系:ChamQ SYBR qPCR Master Mix(诺唯赞,南京)10 μL,F/R引物0.4 μL,模板cDNA 1 μL,双蒸水8.2 μL。qRT-PCR反应条件:95 ℃预变性5 min; 95 ℃变性15 s,引物特异性退火温度下30 s,72 ℃延伸45 s,共40个循环。溶解曲线以0.5 ℃/s的速度从95 ℃降低至65 ℃,每6 s采集1次信号。
2 结果与分析
2.1 lepr cDNA 3′RACE克隆
使用3′ RACE技术克隆获得leprcDNA,发现4个不同大小的DNA片段。经测序发现这4个DNA片段均为leprmRNA经3′可变剪切产生的不同亚型,长度分别为3 474、1 512、945和915 bp。
2.2 lepr基因与氨基酸序列分析
将使用3′RACE技术克隆得到的4条序列与从鳜基因组中获取的lepr序列进行比对分析,发现1个长型受体亚型lepr-L,3个由mRNA 3′端可变剪切产生的短型受体亚型:lepr-S1、lepr-S2、lepr-S3(图1)。lepr-L的CDS长度为3 474 bp,编码1 157个氨基酸;短型受体亚型lepr-S1、lepr-S2和lepr-S3的CDS长度分别为1 512、945和915 bp,分别编码503、314和304个氨基酸。序列分析发现,鳜lepr-L蛋白具有1个信号肽结构、1个跨膜区、1个免疫球蛋白(Ig) C2-like结构域、1个LBD结构域,2个FNIII结构域和2个重复的色氨酸/丝氨酸基序(WSXWS),含有JAK和STAT功能域等全部胞内结构。除lepr-S1蛋白具备不完整的LBD结构域外,其余2个短型受体亚型均不具备LBD结构域,同时,所有短型受体亚型均丢失跨膜区和所有胞内结构域(图2)。如表2所示,鳜lepr-L的氨基酸序列与鱼类同源性高(42%~86%),与哺乳动物同源性低(26%~28%)。

下划线部分氨基酸序列为lepr保守结构。The amino acid sequence of lepr gene conserved domains are underlined.
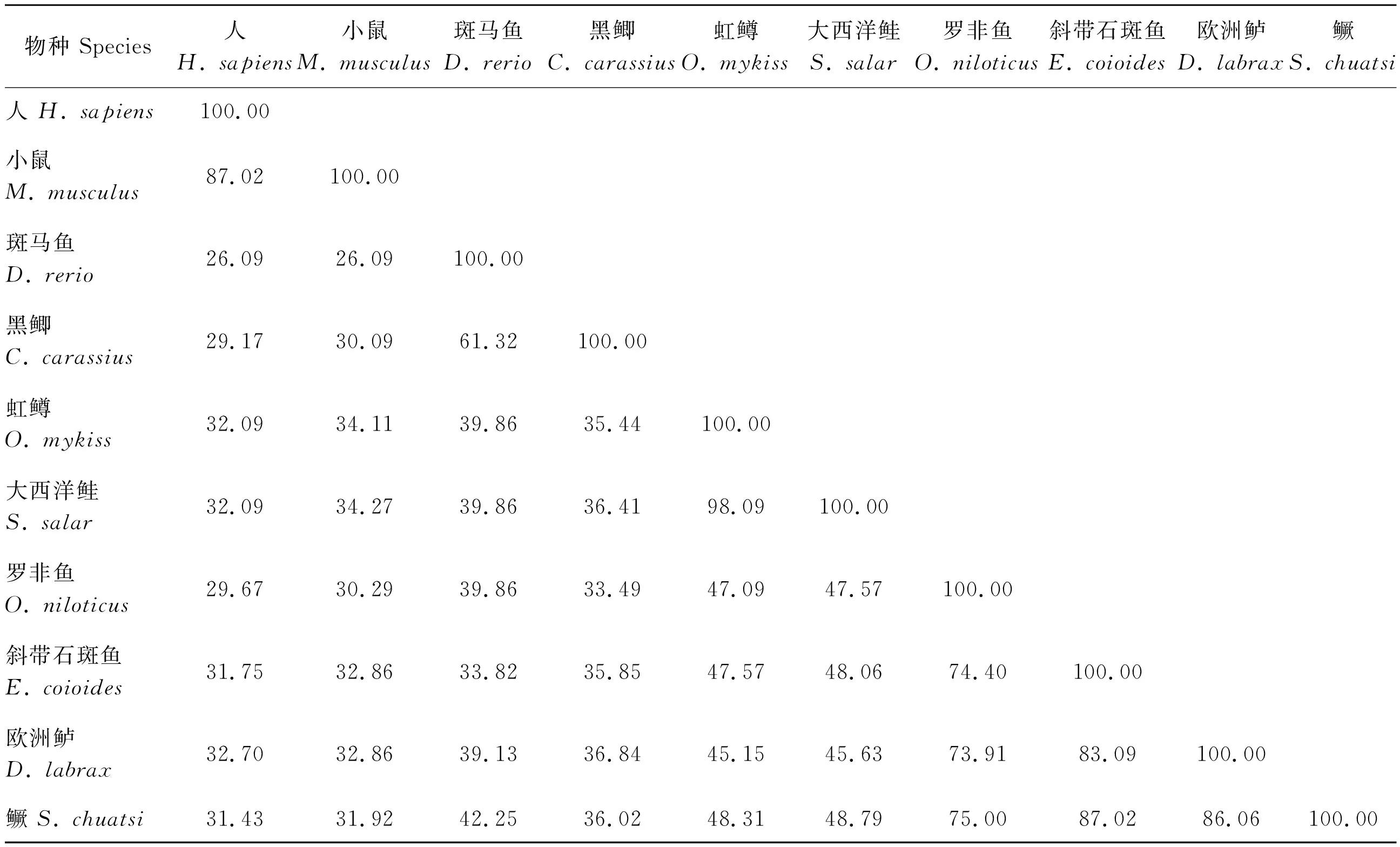
表2 鳜与其他脊椎动物LBD结构域氨基酸序列同源性比较 Table 2 Amino acid homology comparisonof LBD domain between Chinese perch and other vertebrates

黑色方块表示 “WSXWS”基序;箭头位置指示终止密码子; TM:跨膜区。The black bar means “WSXWS” motif; arrows indicates the location of stop codon; TM:Transmembrane domain.
2.3 lepr氨基酸序列多重比对和进化树分析
鳜lepr-L亚型的LBD序列与其他物种氨基酸序列多重比对发现,鳜与石斑鱼、罗非鱼和欧洲鲈同源性最高(75%~87%),其次是虹鳟和大西洋鲑(48%),与斑马鱼、黑鲫、小鼠和人同源性最低(31%~42%)(图3)。

黑色底纹表示氨基酸序列一致 Black shading indicates that the amino acid sequence is consistent.
比对16个不同物种的lepr氨基酸序列并构建系统进化树。如图4所示,lepr进化树整体分为2支,哺乳动物和两栖动物的lepr聚为一支,鱼类的lepr聚为另一支。鳜与鲈形目鱼类的lepr的亲缘关系最近,其次为鲑鳟类,与鲤科鱼类的亲缘关系最远。

图4 脊椎动物lepr氨基酸系统进化树
2.4 lepr在鳜各组织中的表达情况
图5所示为lepr-L在鳜各组织器官中的表达情况。在鳜中,lepr-L在鳃中表达量最高,其次是肾、垂体、肠、脾、脂肪组织、心、肝、中脑、下丘脑和小脑,在小脑和端脑中表达量最低。

T:端脑 Telencephalon; M:中脑 Midbrain; C:小脑 Cerebellum; P:垂体 Pituitary; Ht:下丘脑 Hypothalamus; MO:延脑 Medulla oblongata; L:肝脏 Liver; I:肠 Intestines; K:肾脏 Kidney; A:脂肪组织 Adipose; G:鳃 Gill; S:脾脏 Spleen; H:心脏 Heart.
2.5 腹腔注射鳜leptin A和leptin B同源重组蛋白对脑lepr-L表达的影响
图6所示为腹腔注射鳜同源重组leptin A和leptin B 2 h和4 h后,脑组织中lepr-L的表达情况。注射leptin A 2 h和4 h对鳜脑组织中lepr-L的表达均无显著影响(P>0.05);注射leptin B 2 h鳜脑组织中lepr-L的表达水平显著上调(P<0.05),注射leptin B 4 h后未观察到鳜脑组织中lepr-LmRNA丰度的显著变化(P>0.05)。

含有不同字母的组别之间存在显著性差异(P <0.05),数据以平均值±标准误表示(n=6)。Values within the same row with superscripted letters represent significant difference (P<0.05),data are expressed as mean±S.E (n=6).
3 讨 论
本研究采用3′ RACE克隆的方法,获得了由mRNA 3′端可变剪切产生鳜的4种lepr亚型,包括1个长受体亚型和3个短受体亚型。氨基酸多重比对和进化树分析发现,鳜lepr及其LBD区域均较为保守,与同属鲈形目的欧洲鲈和石斑鱼的同源性最高,亲缘关系最近,其次是鲤科鱼类,与两栖动物和哺乳动物的同源性较低,亲缘关系最远。这与在其他鲈形目鱼类中得到的结果相一致[8,11,24]。
本研究中使用3′ RACE克隆得到鳜的4种亚型的lepr,与哺乳动物和其他鱼类一致的是,长型受体亚型lepr-L具有完整的功能结构域,包括:信号肽、LBD、FNIII,重复的WSXWS、Ig-C2-like、跨膜区、细胞膜内的JAK-STAT结构域。鳜lepr-S1含有LBD,完全缺失了跨膜结构域和胞内域,推测其可能是可溶性leptin受体。在哺乳动物和鱼类中,可溶性受体与循环瘦素结合,避免瘦素被降解,进而调节瘦素的浓度和活动[9,23,25]。鳜短受体亚型lepr-S1结构与哺乳动物和虹鳟的可溶性受体相似,我们推测鳜短受体亚型lepr-S1应当具备分泌型受体的功能,可能具备调节血浆leptin浓度的功能。而鳜短型受体亚型lepr-S2和lepr-S3仅含有一个WSXWS,未见其他重要结构域,在其他鱼类中均未见报道类似的短型受体亚型,其生理功能还需进一步探究。
在鱼类中,lepr在各组织中的表达分布情况与物种相关,例如在金鱼[26]、欧洲鳗鲡[27]和大西洋鲑[12]的端脑、下丘体和性腺中lepr表达量最高,在罗非鱼的垂体中lepr表达最多,其次是肌肉和头肾[26],日本青鳉的lepr则在肌肉中表达量最高,其次是皮肤和鳃[6]。在我们的研究中,鳜lepr-L在鳃中表达最高,其次是肾脏和垂体。此类lepr组织表达分布的差异可能说明leptin在不同的物种中发挥的作用不尽相同。
在哺乳动物中,leptin作为一种饱食因子,通过与专一性受体lepr结合发挥其抑制食欲的生理功能[14-15]。草鱼[28]、鳜[22]、罗非鱼[29]等鱼类腹腔注射同源重组的leptin蛋白后均表现出食欲降低,因此,leptin在鱼类中也被认为具有抑制食欲的作用。同时,敲除lepr基因的日本青鳉也表现出食欲增强[30],也验证了上述观点。然而,lepr敲除的斑马鱼却未表现出食欲的增加[31]。在我们的研究中发现,腹腔注射鳜leptin B同源重组蛋白2 h后,鳜食欲显著降低[22],脑中lepr的表达量随之升高,在注射4 h后,鳜食欲恢复的同时[22],脑中lepr的表达量也随之降低;而注射鳜leptin A同源重组蛋2 h及4 h后,既未观察到食欲明显降低[22],也未观察到脑组织中lepr表达的变化。由此,我们推测,鱼类不同的leptins在不同的组织中对lepr表达的影响不一致,从而行使不同的生理功能。
本研究首次克隆出由mRNA 3′端可变剪切产生的鳜lepr的4个受体亚型。经序列分析发现,仅长型受体lepr-L具有完整的功能结构域,且各功能结构域的氨基酸序列高度保守。短型受体亚型lepr-S1完全缺失跨膜区域和胞内结构域,推测其为可溶性受体亚型,发现了2个在其他鱼类中未见报道的、仅具备1个WSXWS结构的lepr短型受体亚型。鳜lepr基因在鳃、肾和垂体中表达量较高;注射鳜leptin B而非leptin A的同源重组蛋白会引起脑lepr表达量上升。这些结果说明鳜leptins能引起组织中lepr的表达的不同变化而发挥独特的生理功能。
猜你喜欢
——紫 苏
河南农业(2024年1期)2024-01-19 01:56:54
华人时刊(2023年1期)2023-03-14 06:43:36
汉字汉语研究(2021年2期)2021-08-30 08:58:46
小学科学(学生版)(2018年5期)2018-06-15 08:48:00
奇妙博物馆(2018年12期)2018-04-25 02:16:04
奇妙博物馆(2018年12期)2018-04-25 02:16:02
高中生·天天向上(2016年11期)2017-02-28 07:40:18
河北书画研究(2016年3期)2016-04-28 08:55:35
中国病理生理杂志(2015年8期)2015-12-21 12:38:10
中国当代医药(2015年30期)2015-03-01 02:08:19
