
异源表达番茄蔗糖磷酸合成酶基因SlSPS促进拟南芥株型增大研究
2021-07-24 04:36:06刘雅慧朱龙英杨学东朱为民张迎迎
河北农业大学学报 2021年3期
刘雅慧,朱龙英,杨学东,朱为民,张 辉,张迎迎
(上海市农业科学院 园艺研究所/上海市设施园艺技术重点实验室,上海 201403)
番茄(Solanum lycopersicum)是世界范围内广泛种植的重要的蔬菜作物和模式植物,具有重要的营养价值和经济价值[1]。糖度是影响番茄品质性状的主要因子,也是影响其风味品质的重要因素[2],番茄重要性状基因已成为目前的研究热点之一。
蔗糖作为植物光合作用的主要产物,是植物生长、发育及防护的主要碳资源。高等植物中,蔗糖的合成受多种酶控制,其中包括蔗糖磷酸合成酶(Sucrose Phosphate Synthase, SPS),它以尿苷二磷酸葡萄糖(UDPG)为供体,催化6-磷酸果糖合成6- 磷酸蔗糖;6- 磷酸蔗糖再在蔗糖磷酸化酶(Sucrose Phosphate Phosphatase, SPP)的催化下水解脱磷酸形成蔗糖。该反应中的SPS 是植物体内控制蔗糖合成的限速酶[3],SPS 参与植物产量和品质的形成,其活性直接反应了植物体内蔗糖合成的能力[4]。在柑橘、甘蔗、玉米和荔枝中已证实,植物体内蔗糖的积累与SPS 的活性呈正相关[5-8]。最初,认为SPS 是以单基因形式存在于植物体内,随着研究的深入,发现高等植物中存在多个SPS 同工酶,而且属于不同的家族,单子叶植物中含4 个SPS 家族,双子叶植物中含有3 个SPS 家族,由此暗示着不同的SPS 在植物中的功能产生了分工与合作[4, 9-11]。
研究发现,植物内不同的SPS 家族基因的表达特性不同,所发挥的功能也存在着共性和差异性。反义表达CmSPS1的转基因甜瓜蔗糖含量下降、果实变小[12];转菠菜SPS基因的棉花表现出更高的蔗糖/淀粉比率,棉花纤维细胞次生壁增厚,品质提高[13];转玉米SPS基因的马铃薯叶片SPS活性提高,叶片抗衰老,产量增加、蔗糖含量提高[14]。南极发草在南极夏天长白昼的环境下,SPS 活性极高,蔗糖超量积累从而适应极端寒冷气候[15]。低温处理下猕猴桃果实的SPS基因转录水平提高[16]。干旱促进水稻中SPS 活性增加[17]。高盐处理下的玉米秧苗的SPS活性增加[18]。本研究将番茄中的1 个SlSPS基因转化拟南芥进行异源过量表达,并对转基因的拟南芥进行表型观察发现,揭示SlSPS基因影响植物生长发育和蔗糖积累的生物学功能,对番茄的产量和品质改良及其生产应用提供理论依据。
1 材料与方法
1.1 试材和培养方法
以拟南芥(Arabidopsis thaliana)野生型Col-0生态型作为遗传转化的材料和对照组。野生型和转基因植株的种子经消毒后在1/2 MS 培养基上萌发,1 周后移栽至穴盘中,于25 ℃,湿度50%~60%的人工气候室培养。
1.2 基因克隆、蛋白比对与载体构建
番茄蔗糖磷酸合成酶基因SlSPS的编码序列CDS从茄科基因组网站上获得(Solyc07g007790, https://solgenomics.net/),拟南芥同源基因序列从拟南芥信息网站上获得(TAIR, https://www.arabidopsis.or g/)。蛋白比对通过ClustalW(https://www.genome.jp/tools-bin/clustalw) 和ESPript3.0(http://espript.ibcp.fr/ESPript/cgi-bin/ESPript.cgi)完成。从cDNA文库中扩增该基因后,以内切酶KpnI 和XbaI 为酶切位点,设计酶切引物 SlSPS-CDS-F-KpnI,SlSPSCDS-R-XbaI(表 1),通过酶切连接构建到植物双元表达载体pCAMBIA1300-35S 的多克隆位点上(该载体以pCAMBIA1300 为骨架,插入35S 启动子于MCS 前,载体由本实验室保存),得到以CaMV 35S 启动子和SlSPS基因的CDS 融合的表达载体。
1.3 遗传转化及鉴定
SlSPS基因过表达载体pCAMBIA1300-35SSlSPS 转入农杆菌GV3101 后,通过蘸花法转化拟南芥野生型Col-0,获得独立转化株系。经过潮霉素抗性培养基的筛选后,利用PCR 扩增潮霉素基因、SlSPS基因(鉴定引物见表1)鉴定阳性幼苗后,随后通过实时荧光定量PCR 鉴定SlSPS1基因的表达量。所获得的3 个独立转化株系用于后续的试验研究。

表1 鉴定转基因植株的PCR 引物Table1 Primer sequence for identification of transgenic lines
1.4 表达量鉴定
取10 d 期野生型和T3 代转基因拟南芥的整株幼苗,经液氮速冻后研磨,采用翊圣植物RNA 提取试剂盒提取RNA,然后使用翊圣反转录酶Hifair™II 1st Strand cDNA Synthesis Super Mix for qPCR(gDNA digester plus) 反转录得到cDNA。 针对SlSPS基因的CDS 设计引物(表达量鉴定引物见表1),以Actin2基因为对照,在Applied Biosystems QuantStudio 5 系统上进行qPCR 检测。通过相对定量2—ΔΔCT法分析结果。
1.5 蔗糖、可溶性糖及酶活测定
蔗糖磷酸合成酶催化果糖-6-磷酸形成蔗糖磷酸,蔗糖磷酸与间苯二酚反应可呈现颜色变化,在480 nm 下有特征吸收峰,酶活力大小与颜色的深浅成正比。在野生型和T3 代转基因拟南芥幼苗生长4周时,将同一独立转化株系的3 株长势一致的莲座叶同一部位的叶片混合取样,液氮研磨匀浆。蔗糖测定采用间苯二酚法测定,可溶性糖测定采用蒽酮比色法,蔗糖磷酸合成酶酶活测定采用间苯二酚法测定。上述测定采用苏州科铭生物技术有限公司的试剂盒,于可见光分光光度计下测定。
1.6 表型鉴定
表型鉴定采用野生型和T3 代转基因植株进行分析。拟南芥放在人工气候室(温度25 ℃,14 h 光照/10 h 黑暗),从种子萌发后3 d 开始观察分析,转基因植株和野生型的成苗高度、根长、果荚通过直尺或游标卡尺进行测量。同时将不同的材料在黑暗下培养7 d 测定下胚轴长度。取长势一致的野生型和转基因植株各10 株,进行比较和测量。成苗高度和果荚在6 周时测量,根长和下胚轴长度在1 周时测量。
2 结果与分析
2.1 拟南芥过表达番茄蔗糖磷酸合成酶基因SlSPS的克隆及转基因植株的获得和鉴定
根据已报道的蔗糖磷酸合成酶的蛋白序列,在茄科植物基因组网站中进行了同源比对,结合最新的番茄基因组注释ITAG4.0(ftp://ftp.solgenomics.net/tomato_genome/annotation/ITAG4.0_release/),获得了番茄中蔗糖磷酸合成酶基因的基因号(Solyc07g007790)及其序列。对其序列和拟南芥中SPS基因(At5g11110)进行蛋白同源性比对发现,其蛋白相似度达到69.29%(图1A)。通过基因合成,将SlSPS基因的CDS 区域经PCR 扩增后通过酶切连接的手段克隆到植物双元表达载体pCAMBIA 1300-35S 上,并测序验证,得到pCAMBIA1300-35S-SlSPS载体(图1C)。载体转化到农杆菌后进行遗传转化,得到T0 代幼苗。对T0 代幼苗的基因组DNA 进行PCR 扩增,利用潮霉素抗性基因和目标基因SlSPS基因,筛选阳性幼苗。对T3 代幼苗进行了SlSPS基因表达量鉴定,其中,获得了3 个表达量显著上调的独立转化株系SlSPS-OE20,SlSPSOE44 和SlSPS-OE60(图1B),用于后续的表型分析和生理指标检测。

图1 SlSPS 蛋白比对及表达量分析Fig.1 Alignment of SlSPS protein sequence and expression level analysis
2.2 过表达SlSPS 基因拟南芥转基因植株中蔗糖磷酸合成酶活性提高
对转基因植株蔗糖磷酸合成酶的酶活测定后,进一步验证在拟南芥中过表达SlSPS基因的酶活性(图2A)。过表达转基因株系均呈现出蔗糖磷酸合成酶活性升高的趋势。3 个独立转化株系的酶活与野生型相比提高了20%~30%。该结果表明过表达番茄SlSPS基因能够提高拟南芥转基因植株的蔗糖磷酸合成酶的活性,正常行使SlSPS基因预测的生物学功能。
2.3 过表达SlSPS 基因拟南芥转基因植株的蔗糖、可溶性糖的含量显著上升
蔗糖磷酸合成酶是蔗糖合成过程的关键限速酶。可溶性糖包括葡萄糖、果糖、蔗糖,是番茄品质的重要构成性状之一。通过对过表达SlSPS基因在转基因植株中蔗糖、可溶性糖含量的测定,进一步挖掘SlSPS基因的直接生化功能。对拟南芥生长4 周幼苗叶片中的蔗糖、可溶性糖进行测定(图2),结果表明:与野生型Col-0 相比(0.673 μmol/mgprot),过表达转基因植株中的蔗糖含量(1.269 μmol/mgprot,1.148 μmol/mgprot,1.104 μmol/mgprot) 显 著 提高;转基因植株中的可溶性糖的含量(7.155 mg/g,7.692 mg/g,7.302 mg/g)也显著高于野生型Col-0(5.962 mg/g)。这表明过表达SlSPS基因提升了拟南芥中蔗糖的含量,同时可溶性糖的含量也随之升高。
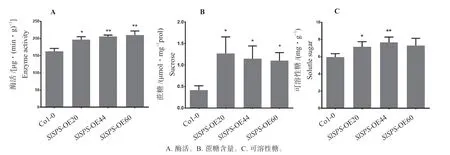
图2 转基因和野生型植株中蔗糖磷酸合成酶酶活、蔗糖和可溶性糖水平分析Fig.2 Analysis of sucrose phosphate synthase activity and content of sucrose and soluble sugar in transgenic lines and wild type Col-0
2.4 过表达SlSPS 基因的拟南芥转基因植株的株型增大
过表达SlSPS基因不仅提高了拟南芥中SPS 的活性,蔗糖和可溶性糖的含量,还进一步促进了转基因植株的株型。选取种子萌发后5 d(5 days after germination, 5DAG)( 图3 A),1 周(1 week)(图3E),10 d(10 days after germination, 10DAG) (图3B),2 周(2 week)(图3F),4 周(4 weeks)(图3C)和6 周(weeks)(图3D)的植株观察叶片和株型大小。发现转基因植株从营养生长期至生殖生长期,其植株大小,包括叶片大小、根长、莲座和成苗高度,相较于野生型都增大。5DAG 时即出现明显的叶片增大的表型。1 周时测量根长,相较于野生型其根长显著伸长了12.15%(图3H)。2 周时,其株型大小和每片叶的大小都增大。到了6 周时,其成苗高度升高了24.69%,显著高于野生型(图3G)。以上结果表明过表达SlSPS基因促进了拟南芥整个生育期的株型增大,其叶片、根长和成苗高度都显著增大。

图3 过表达SlSPS 基因转基因植株的表型分析Fig.3 Phenotype analysis of transgenic lines with overexpression of SlSPS gene
2.5 过表达SlSPS 基因的拟南芥转基因植株黑暗下下胚轴显著伸长
为了探究过表达SlSPS基因对拟南芥暗形态建成的影响,将消毒后的种子培养于1/2 MS 培养基上避光垂直培养,观察和测量其下胚轴(图4)。如图所示,暗培养下转基因植株的下胚轴显著长于野生型,增加了17.09%。表明过表达SlSPS基因能促进暗形态建成。
2.6 过表达SlSPS 基因的拟南芥转基因植株花和果荚增大,种子也变大
通过对花蕾(图5A、B)、果荚(图5D)和种子大小(图5C)的观察和测量,以探究过表达SlSPS基因对于生殖器官发育的影响。结果显示,转基因植株的花蕾更大,其果荚长度显著长于野生型,增长了25.17%(图5E)。此外,转基因植株的种子也大于野生型(图5)。以上结果表明,过表达SlSPS基因能够促进拟南芥中生殖器官的变大。
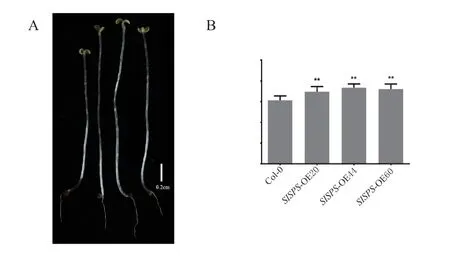
图4 暗培养下过表达SISPS 基因转基因植株下胚轴长度差异Fig.4 The significant elongation of hypocotyls of transgenic lines with overexpression of SISPS gene in dark culture
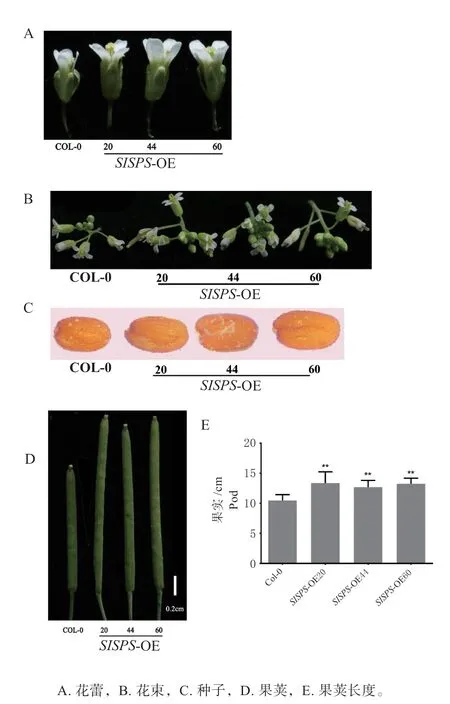
图5 过表达SlSPS 基因转基因植株的生殖器官表型Fig.5 Phenotypes of reproductive organs of transgenic lines with overexpression of SlSPS gene
3 结论与讨论
蔗糖是植物光合作用的重要产物,同时参与生长发育过程中的合成和代谢过程。番茄中蔗糖参与其产量和风味品质的形成。而蔗糖磷酸合成酶是蔗糖合成过程中的关键限速酶,其活性与植物体内蔗糖合成能力直接相关。本研究在拟南芥中异源表达番茄蔗糖磷酸合成酶基因SlSPS,发现转基因株系中,SlSPS基因表达量显著上调,且SPS 酶活性、蔗糖和可溶性糖含量显著升高;表型分析结果显示拟南芥转基因植株增大,其中包括根长伸长、叶片增大、莲座大小增大、成苗高度增高;暗形态建成中下胚轴显著伸长;生殖器官(花蕾、果荚、种子)大小增大。以上结果表明,在拟南芥中过表达番茄SlSPS基因能够提高SPS 酶活、蔗糖和可溶性糖含量,增强暗形态建成,促进植株的营养器官和生殖器官的增大,全面提高植株大小。
异源表达SlSPS基因促进了拟南芥中SPS 酶活增加、蔗糖含量提高,这与基因注释的功能吻合。同时,已报道的研究中,证实了在柑橘、甘蔗、玉米和荔枝中,植物体内蔗糖的积累与SPS 的活性呈正相关[5-8];反义表达SPS基因的转基因甜瓜蔗糖含量下降[12];转菠菜SPS基因的棉花表现出更高的蔗糖/淀粉比率,棉花纤维细胞次生壁增厚,品质提高[13];转玉米SPS基因的马铃薯叶片SPS 活性提高,产量增加、蔗糖含量提高[14]。本试验结果进一步说明,SlSPS基因与其他物种中的同源基因具有相同或相似的功能,均能够提高SPS 活性,促进蔗糖的合成。此外,本研究发现可溶性糖水平也得到提高,说明蔗糖含量提高并不影响其他可溶性糖的合成,比如果糖、葡萄糖。我们进一步推测SPS 催化蔗糖合成对其他可溶性糖合成没有反馈抑制的影响,说明SPS基因可以应用于提高作物中蔗糖和可溶性糖的含量,从而提高糖分积累,改良产量和品质。
为了适应土壤中的生长环境,幼苗形态发育为暗形态建成,表现为下胚轴快速向上伸长,子叶扩展被严格抑制并闭合,减少向上生长所遇到的土壤阻力。幼苗出土后,下胚轴伸长抑制,子叶快速打开和扩展,以吸收更多阳光,实现光合自养生长。幼苗出土前后的形态建成巨变,与所处土壤环境的剧变紧密联系,是研究植物适应环境的经典模型[19-20]。本研究发现过表达SlSPS基因的拟南芥转基因幼苗,相较于野生型,具有更加突出的暗形态建成的表型,具有更显著的下胚轴伸长。这表明SlSPS基因可以促进暗形态建成,可能使得种子破土而出的存活能力提高,活力增强。
过表达SlSPS基因使得拟南芥植株的营养器官和生殖器官都变大。在其营养生长时期,其幼苗根变长,莲座叶片增大,成苗高度增高;而在其生殖生长时期,其花蕾、果荚和种子的大小都增大。这表明SlSPS基因能够在整个生育期促进植株株型变大。SPS 可作为高糖积累的生化指标[20]。在幼苗期,通过促进根的伸长提高对水分和无机盐的吸收,通过促进叶片的增大提高了光合作用的效率,由此可能促进能量的累积,增强了碳的固定,进而促进了生殖器官的增大。因此,我们推测过表达SlSPS基因通过促进根的伸长、叶片增大从而提高对营养物质的固定,促进生殖器官的发育,从而使得植株生长更加旺盛,提高产量。
综合本研究结果,在拟南芥中异源过表达SlSPS基因能够提高SPS 酶活性、蔗糖和可溶性糖含量,增强暗形态建成,推测可能提高了能量的累积,同时促进植株的营养器官和生殖器官的增大。本研究有助于对SlSPS的基因功能及其参与蔗糖合成以及下游代谢机制的研究,从而挖掘该基因对植物生长发育的影响,并说明SlSPS基因可以用于改良番茄的产量和品质,为分子设计育种提供遗传资源。
猜你喜欢
中学生天地(A版)(2023年1期)2023-02-17 00:33:04
广西糖业(2020年3期)2020-09-25 08:32:44
中国化肥信息(2019年12期)2020-01-16 08:40:06
中国化肥信息(2018年7期)2018-08-23 09:12:32
中国化肥信息(2018年6期)2018-08-23 09:11:42
生命科学研究(2018年1期)2018-05-29 01:12:47
浙江工业大学学报(2017年5期)2018-01-22 02:03:33
中国化肥信息(2017年7期)2017-12-13 08:46:28
上海农业学报(2017年3期)2017-04-10 12:39:14
中国糖料(2016年1期)2016-12-01 06:49:06
