
2020年CHINET中国细菌耐药监测
2021-07-21 09:38胡付品朱德妹蒋晓飞徐英春张小江张朝霞王传清王爱敏徐元宏孙自镛陈中举倪语星孙景勇褚云卓田素飞胡志东0金0俞云松郭素芳魏莲花邹凤梅胡云建艾效曼苏丹虹郭大文赵金英黄湘宁刘文恩0李艳明0邵春红徐雪松王山梅楚亚菲张利侠周树平孟晋华郑红艳胡芳芳周万青伟0刚0吴劲松卢月梅李继红段金菊康建邦马晓波郑燕萍郭如意陈运生王世富胡雪飞沈继录汪瑞忠0华0俞碧霞温开镇张贻荣刘江山廖龙凤顾洪芹薛顺虹佼0岳春雷
中国感染与化疗杂志 2021年4期
胡付品,郭 燕,朱德妹,汪 复,蒋晓飞,徐英春,张小江,张朝霞,季 萍,谢 轶,康 梅,王传清,王爱敏,徐元宏,黄 颖,孙自镛,陈中举,倪语星,孙景勇,褚云卓,田素飞,胡志东0,李 金0,俞云松,林 洁,单 斌,杜 艳,郭素芳,魏莲花,邹凤梅,张 泓,王 春,胡云建,艾效曼,卓 超,苏丹虹,郭大文,赵金英,喻 华,黄湘宁,刘文恩0,李艳明0,金 炎,邵春红,徐雪松,鄢 超,王山梅,楚亚菲,张利侠,马 娟,周树平,周 艳,朱 镭,孟晋华,董 芳,郑红艳,胡芳芳,沈 瀚,周万青,贾 伟0,李 刚0,吴劲松,卢月梅,李继红,段金菊,康建邦,马晓波,郑燕萍,郭如意,朱 焱,陈运生,孟 青,王世富,胡雪飞,沈继录,汪瑞忠0,房 华0,俞碧霞,赵 勇,龚 萍,温开镇,张贻荣,刘江山,廖龙凤,顾洪芹,姜 琳,贺 雯,薛顺虹,冯 佼0,窦 睿,岳春雷
近年来,临床上重要耐药细菌,尤以革兰阴性杆菌为代表的多重耐药细菌的检出率呈快速上升趋势,为临床的抗感染治疗带来巨大挑战[1]。细菌耐药监测工作是了解耐药菌变迁、遏制耐药菌进一步流行播散最重要的基础工作之一。现将2020年CHINET中国细菌耐药监测结果报道如下。
1 材料与方法
1.1 材料
1.1.1 细菌来源 收集2020年1月1日—12月31日来自全国29个省市或自治区52所医院(45所综合性医院和7所儿童专科医院) 的临床分离株,剔除同一患者分离的重复菌株,按CHINET 2020统一方案进行细菌对抗菌药物的敏感性试验,剔除非无菌体液标本分离的凝固酶阴性葡萄球菌和草绿色链球菌。
1.2 方法
1.2.1 药敏试验 参照2020年美国临床和实验室标准化协会(CLSI)推荐的药敏试验要求[2],按CHINET中国细菌耐药监测网技术方案、采用纸片扩散法或自动化仪器法进行。药敏试验质控菌为:金黄色葡萄球菌(金葡菌)ATCC 25923、大肠埃希菌ATCC 25922、铜绿假单胞菌ATCC 27853、肺炎链球菌ATCC 49619和流感嗜血杆菌ATCC 49247。
1.2.2 判断标准 参照2020年CLSI M100-S30文件推荐的判断标准[2]。其中磷霉素的判断标准仅针对尿标本分离的大肠埃希菌和粪肠球菌,多黏菌素的判断标准采用《多黏菌素药物敏感性检测及临床解读专家共识》[3],替加环素按美国食品和药品监督管理局(FDA)推荐的判断标准[4]。
1.2.3 β内酰胺酶检测 采用头孢硝噻吩试验定性检测流感嗜血杆菌中的β内酰胺酶。按CLSI推荐的纸片法进行酶抑制剂增强试验[2],确证大肠埃希菌、肺炎克雷伯菌、产酸克雷伯菌和奇异变形杆菌中产超广谱β内酰胺酶(ESBL)菌株。
1.2.4 青霉素不敏感肺炎链球菌的检测 经1 μg/片苯唑西林纸片法测定抑菌圈直径≤19 mm的肺炎链球菌菌株,采用青霉素E试验条测定其最低抑菌浓度(MIC),脑膜炎株和非脑膜炎株分别按2020年CLSI M100文件相关标准[2]判定为青霉素敏感(PSSP)、中介(PISP)或耐药(PRSP)。
1.2.5 糖肽类不敏感革兰阳性球菌检测 常规药敏试验显示对万古霉素、利奈唑胺或替考拉宁不敏感(包括中介和耐药)的菌株,采用其他方法测定这些细菌对上述药物的MIC值,加以复核确认。
1.2.6 特殊耐药菌株定义 碳青霉烯类耐药肠杆菌目细菌(CRE)定义为对亚胺培南、美罗培南或厄他培南中任一种抗生素耐药者[5]。其中摩根菌属、变形杆菌属等细菌对除了亚胺培南之外的任一碳青霉烯类抗生素耐药者。
1.2.7 数据统计分析 采用WHONET 5.6软件进行数据统计分析。
2 结果
2.1 细菌分布
2020年共收集临床分离株251 135株,其中革兰阳性菌和革兰阴性菌分别占28.1% (70 624株)和71.9%(180 511株);住院患者和门急诊患者分离的菌株分别占87.7%(220 143株)和12.3%(30 992株)。标本分布中痰液等呼吸道分泌物占36.2%、尿液20.8%、血液15.0%、伤口脓液6.7%、脑脊液1.1%、其他无菌体液5.7%、生殖道分泌物1.2%、粪便1.1%和其他标本12.2%。肠杆菌目细菌占全部分离菌株的45.8%(114 970株),此菌群中分离率占前三位的是大肠埃希菌(43.0%)、肺炎克雷伯菌(32.3%)和阴沟肠杆菌(6.4%)。不发酵糖革兰阴性杆菌占总分离菌株的23.8%(59 703株),其中分离率占前三位的是铜绿假单胞菌(35.4%)、鲍曼不动杆菌(33.3%)和嗜麦芽窄食单胞菌(12.5%)。70 624株革兰阳性菌中最多见者依次为金葡菌(32.9%)、屎肠球菌(15.5%)、粪肠球菌(13.5%)和肺炎链球菌(7.4%)。主要细菌菌种分布见表1。

表1 2020年主要监测细菌菌种分布Table 1 Distribution of bacterial species in 2020
2.2 革兰阳性球菌对抗菌药物的敏感率和耐药率
2.2.1 葡萄球菌属 金葡菌中甲氧西林耐药株(MRSA)的检出率为31.0%,表皮葡萄球菌中甲氧西林耐药株(MRSE)的检出率为81.7%,其他葡萄球菌属细菌(除假中间葡萄球菌和施氏葡萄球菌外)中甲氧西林耐药株(MRCNS)的检出率为77.5%。MRSA和其他MRCNS对大环内酯类、氨基糖苷类和喹诺酮类等抗菌药物的耐药率均显著高于甲氧西林敏感株(MSSA和其他MSCNS)。但MRSA对甲氧苄啶-磺胺甲唑的耐药率低于MSSA(6.4% 对14.5%)。MRSE对甲氧苄啶-磺胺甲唑的耐药率明显高于MRSA(56.1%对6.4%),但对克林霉素的耐药率则显著低于MRSA(32.0%对58.6%)。葡萄球菌属中未发现万古霉素耐药株,极少数MRCNS对替考拉宁或利奈唑胺耐药。见表2。

表2 葡萄球菌属对各种抗菌药物的耐药率和敏感率Table 2 Susceptibility of Staphylococcus spp. to antimicrobial agents(%)
2.2.2 肠球菌属 肠球菌属细菌中粪肠球菌和屎肠球菌分别占43.2%(9 555/22 135)和49.4% (10 931/22 135),其他肠球菌占7.4%(1 649/22 135)。粪肠球菌对绝大多数受试抗菌药物的耐药率均显著低于屎肠球菌,其对呋喃妥因、磷霉素和氨苄西林的耐药率较低,分别为2.7%、3.6%和3.8%,但屎肠球菌对呋喃妥因和氨苄西林的耐药率均较高,分别为53.5%和90.6%。两者对高浓度庆大霉素的耐药率分别为36.6%和45.0%。粪肠球菌和屎肠球菌中均有少数万古霉素、替考拉宁和利奈唑胺耐药株,利奈唑胺耐药菌株粪肠球菌(3.3%)多于屎肠球菌(0.6%),而万古霉素耐药菌株屎肠球菌(1.1%)多于粪肠球菌(0.1%)。见表3。

表3 粪肠球菌和屎肠球菌对抗菌药物的耐药率和敏感率Table 3 Susceptibility of E. faecalis and E. faecium to antimicrobial agents (%)
2.2.3 链球菌属 儿童患者分离的3 836株肺炎链球菌中,PSSP、PISP和PRSP的检出率分别为96.7%、2.6%和0.7%;成人患者中分离的1 264株肺炎链球菌中,PSSP、PISP和PRSP分别为95.5%、3.8%和0.7%。见表4。药敏试验结果显示儿童株和成人株对红霉素、克林霉素和甲氧苄啶-磺胺甲唑耐药率均较高。儿童患者分离的PSSP 中出现极少数左氧氟沙星或莫西沙星的耐药株(耐药率<0.3%),但较成人株(2.8%~4.7%)为少。未发现万古霉素和利奈唑胺耐药株。见表5。

表4 儿童和成人患者中非脑膜炎肺炎链球菌的分布Table 4 The distribution of nonmeningitis S. pneumoniae isolates from children and adults
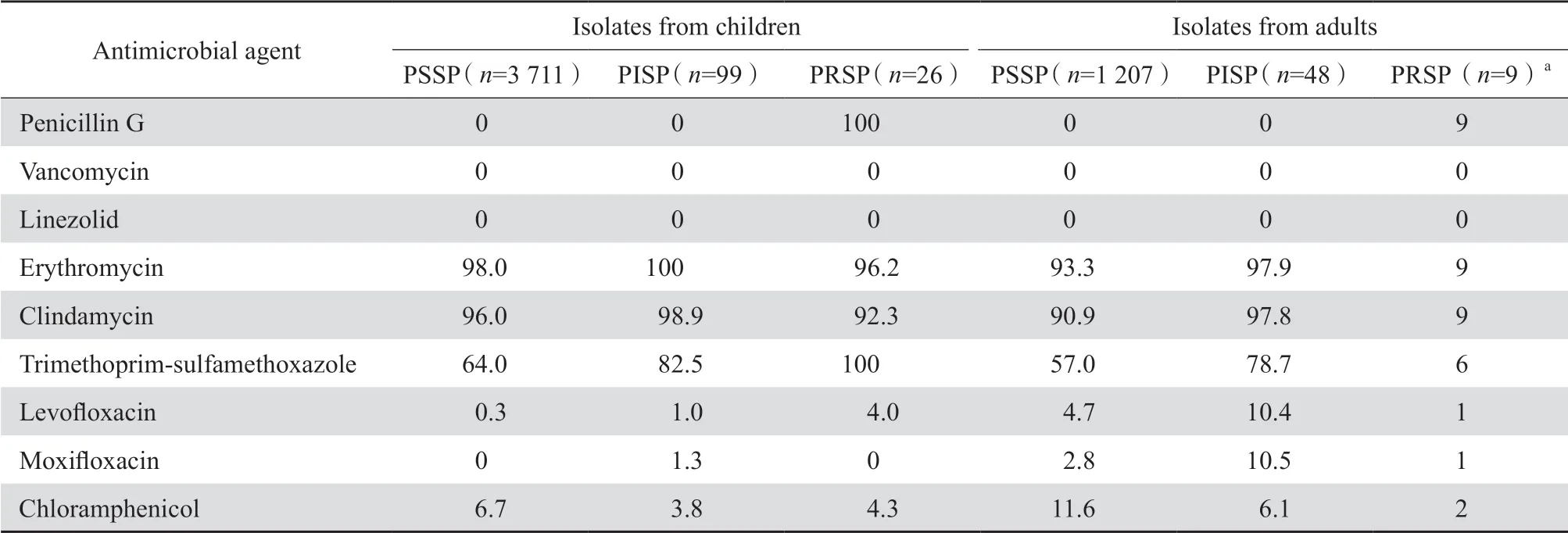
表5 儿童和成人患者中分离的非脑膜炎肺炎链球菌的耐药率Table 5 Susceptibility of nonmeningitis S. pneumoniae isolated from children and adults(%)
监测到A、B、C、F、G各组β溶血链球菌分别为917、4 391、372、31、31株,以及血液或脑脊液等无菌体液标本中的草绿色链球菌2 489株。除草绿色链球菌对青霉素的耐药率为6.7%外,其他链球菌属细菌未发现对青霉素耐药株。大部分链球菌属对红霉素和克林霉素的耐药率超过75%。除B组β溶血链球菌对左氧氟沙星的耐药率为46.2%,为最高,其他β溶血链球菌对左氧氟沙星均高度敏感,耐药率为0.9%~4.1%。未发现万古霉素和利奈唑胺耐药的链球菌属细菌。见表6。

表6 链球菌属对抗菌药物的耐药率Table 6 Resistance rates of Streptococcus spp. to antimicrobial agents (%)
2.3 革兰阴性杆菌对抗菌药物的敏感率和耐药率
2.3.1 肠杆菌目细菌 大肠埃希菌对头孢曲松、哌拉西林、甲氧苄啶-磺胺甲唑、环丙沙星和左氧氟沙星的耐药率均超过50%;肠杆菌目细菌对3种碳青霉烯类的耐药率仍较低,除克雷伯菌属细菌对碳青霉烯类的耐药率范围为17.4%~22.4%外,其他细菌的耐药率多在12%以下。见表7。2005—2020年16年的监测显示,肺炎克雷伯菌对亚胺培南和美罗培南的耐药率从2005年的3.0%和2.9%持续上升至2018年的25.0%和26.3%,但从2019年和2020年开始呈连续下降趋势 (图1)。鼠伤寒沙门菌和肠沙门菌对氨苄西林的耐药率均超过80%,但伤寒沙门菌和副伤寒沙门菌对氨苄西林的耐药率仅为44.6%;所有沙门菌属细菌对碳青霉烯类均100%敏感。见表8。肠杆菌目细菌对常用抗菌药物的总耐药率和敏感率见表9,其中细菌对替加环素、多黏菌素、2种碳青霉烯类和阿米卡星的耐药率最低,为2.4%~10.7%,对哌拉西林-他唑巴坦和头孢哌酮-舒巴坦的耐药率均为14.0%,对头孢他啶和环丙沙星的耐药率分别为28.2%和41.5%。
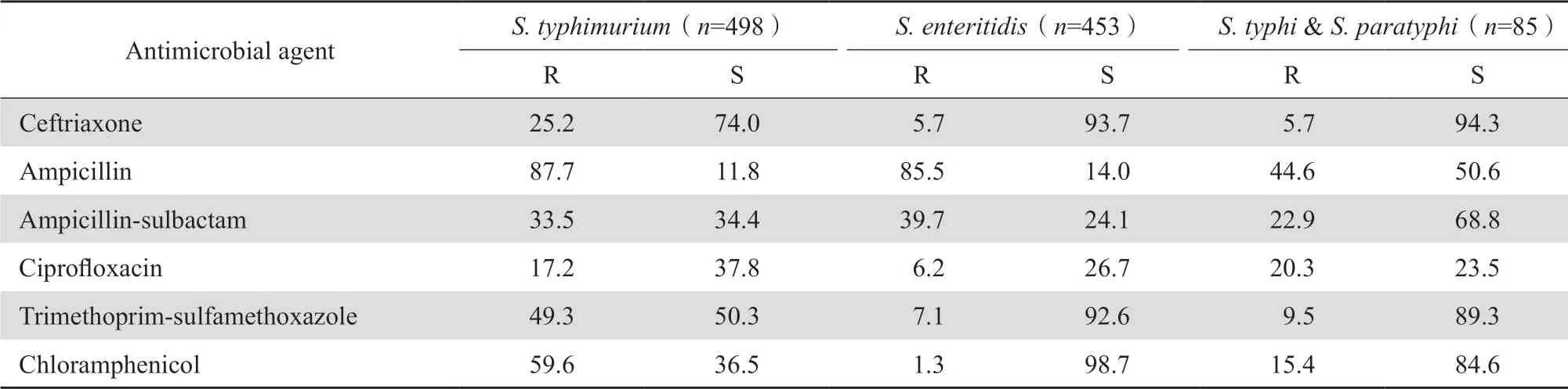
表8 沙门菌属对抗菌药物的耐药率和敏感率Table 8 Susceptibility of Salmonella spp. to antimicrobial agents (%)

表9 肠杆菌目细菌对抗菌药物总耐药率和敏感率Table 9 Susceptibility of all Enterobacterales strains to antimicrobial agents

图1 2005—2020年肺炎克雷伯菌对亚胺培南和美罗培南耐药变迁Figure 1 The changing trend of imipenem and meropenem resistance in K. pneumoniae isolates from 2005 to 2020

表7 肠杆菌目细菌对抗菌药物的耐药率和敏感率Table 7 Susceptibility of Enterobacterales strains to antimicrobial agents(%)
2.3.2 不发酵糖革兰阴性杆菌 铜绿假单胞菌对亚胺培南和美罗培南的耐药率分别为23.2%和19.3%;对多黏菌素B、黏菌素和阿米卡星的耐药率分别为0.7%、2.0%和4.5%;对所测试的两种酶抑制剂合剂、庆大霉素、环丙沙星、左氧氟沙星、头孢他啶、头孢吡肟和哌拉西林的耐药率<17%。铜绿假单胞菌对亚胺培南和美罗培南的耐药变迁见图2。不动杆菌属细菌对亚胺培南和美罗培南的耐药率分别为68.1%和69.0%;对头孢哌酮-舒巴坦和米诺环素的耐药率分别为46.3%和23.7%,对多黏菌素B、黏菌素和替加环素的耐药率较低(1.0%~2.9%),对其他受试药的耐药率均在50%以上。嗜麦芽窄食单胞菌对甲氧苄啶-磺胺甲唑和米诺环素的耐药率低于10%,对左氧氟沙星的耐药率为10.8%。洋葱伯克霍尔德菌对美罗培南、头孢他啶、米诺环素和甲氧苄啶-磺胺甲唑的耐药率范围为7.0%~13.4%,但对左氧氟沙星的耐药率21.6%。见表10。

表10 不发酵糖革兰阴性菌对抗菌药物的耐药率和敏感率Table 10 Susceptibility of non-fermentative gram-negative bacilli to antimicrobial agents(%)
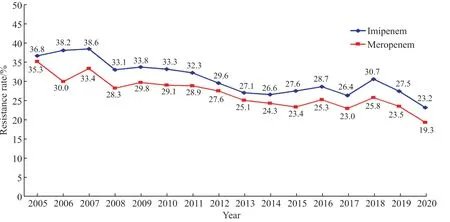
图2 2005—2020年铜绿假单胞菌对亚胺培南和美罗培南耐药变迁Figure 2 The changing trend of imipenem and meropenem resistance in P. aeruginosa isolates from 2005 to 2020
2.4 其他革兰阴性杆菌
2.4.1 流感嗜血杆菌 4 065株流感嗜血杆菌中,儿童分离株和成人分离株分别占60.1%( 2 443株)、39.9%(1 622株);β内酰胺酶的检出率分别占各自分离株的56.3%和45.6%。流感嗜血杆菌对氨苄西林的耐药率均超过50%。大多流感嗜血杆菌对头孢曲松、美罗培南和左氧氟沙星、以及氯霉素均高度敏感,但对前三者均已出现少数不敏感株(儿童株0.3%~1.8%,成人株2.2%~5.4%)。相比较而言,儿童分离株对氨苄西林、头孢呋辛、阿奇霉素和甲氧苄啶-磺胺甲唑的耐药率高于成人株。见表11。

表11 流感嗜血杆菌对抗菌药物的耐药率和敏感率Table 11 Susceptibility of H. influenzae to antimicrobial agents(%)
3 讨论
2020年CHINET中国细菌耐药监测结果具有以下特点:①2020年收集的总菌株数为251 135株,较2019年的270 497株减少7.2%[6-7]。前五位的细菌排名与往年一样,分别为大肠埃希菌、肺炎克雷伯菌、金葡菌、铜绿假单胞菌和鲍曼不动杆菌。②金葡菌中MRSA菌株的检出率继续下降,由2019年的31.4%继续下降至31.0%,表皮葡萄球菌中MRSE菌株的检出率由2019年的82.4%下降至81.7%,其他凝固酶阴性葡萄球菌中MRCNS的检出率由2019年的78.3%下降至77.5%。③肺炎克雷伯菌对亚胺培南和美罗培南的耐药率分别 从2005年 的3.0%和2.9%上 升 到2018年 的25.0%和26.3%,耐药率上升幅度超过8倍,但2019年和2020年呈现持续下降。然而在革兰阴性杆菌中该菌的检出率占比从2019年的20.2%上升至20.6%。④美罗培南耐药铜绿假单胞菌的检出率连续两年呈下降趋势,从2019年的23.5%下降至19.3%;美罗培南耐药鲍曼不动杆菌的检出率首次出现下降,从2019年的79.0%下降至73.4%。
2020年CLSI文件对原来的肠杆菌科细菌的药敏试验标准涉及的细菌重新分类,升级为包含7个科的肠杆菌目细菌,分别是布杰约维采菌科、肠杆菌科、欧文菌科、哈夫尼亚菌科、摩根菌科、溶果胶菌科和耶尔森菌科[2]。其中埃希菌属、克雷伯菌属、肠杆菌属、枸橼酸杆菌属、沙门菌属、志贺菌属等仍然归为新命名的肠杆菌科细菌。当前,产ESBL和碳青霉烯酶仍是包含肠杆菌科细菌在内的肠杆菌目细菌最重要的耐药机制[1]。由于碳青霉烯酶的产生及流行,使得CLSI推荐的ESBL酶抑制剂增强试验因无法获取ESBL正确的检出率或发生率而受到严重挑战;再者2010年以来CLSI进行了头孢菌素和碳青霉烯类对肠杆菌科细菌折点的系列调整和修改,新折点下的药敏试验结果的“敏感”或“耐药”的判断无需再考虑菌株ESBL的检出,故目前CLSI已不再规定实验室常规进行ESBL的检测并报告。由于我国上述临床分离菌株产生的ESBL主要为水解头孢噻肟或头孢曲松的CTX-M型ESBL,有文献报道可用细菌对头孢曲松或头孢噻肟的耐药率来反映细菌产生的ESBL[8]。2020年监测结果显示,大肠埃希菌、肺炎克雷伯菌和奇异变形杆菌中产ESBL菌株的检出率分别为55.5%、44.8%和39.0%[9]。对2005—2020年分离的肺炎克雷伯菌对碳青霉烯类的耐药变迁结果发现,2005—2018年,肺炎克雷伯菌对碳青霉烯类的耐药率呈持续上升趋势,2019年和2020年肺炎克雷伯菌对碳青霉烯类的耐药率虽然呈连续下降趋势,但其检出率仍超过23%。与肺炎克雷伯菌相似的是,铜绿假单胞菌对亚胺培南和美罗培南的耐药率亦连续两年呈下降趋势,鲍曼不动杆菌对亚胺培南和美罗培南的耐药率首次出现下降现象。
为应对碳青霉烯类耐药革兰阴性杆菌带来的重大挑战,临床急需新型广谱有效的抗菌药物以挽救患者的生命。替加环素、多黏菌素(包括多黏菌素B和黏菌素)和头孢他啶-阿维巴坦被认为是治疗碳青霉烯类耐药革兰阴性杆菌所致感染最有效的“三剑客”。但研究显示,实验室做好上述三种药物的药敏试验并不容易。①替加环素:由于替加环素含有多酚基团容易被空气中的氧气氧化降解,常规药敏试验方法容易出现假中介或假耐药结果[10],此时需以标准的肉汤微量稀释法或含替加环素复敏液的方法进行复核确认[11]。②多黏菌素B和黏菌素:CLSI和EUCAST明确规定,肉汤微量稀释法是测定多黏菌素类药物唯一可靠的药敏试验方法,且2020年CLSI文件取消了此类药物的敏感标准,仅设置中介和耐药标准。由于多黏菌素类药物在治疗碳青霉烯类耐药革兰阴性杆菌中仍发挥重要的作用,建议实验室在报告药敏试验结果时,根据杨启文等[3]的《多黏菌素药物敏感性检测及临床解读专家共识》的建议,MIC≤2 mg/L时报告为敏感,≥4 mg/L时报告为耐药,并按要求添加相应的备注信息。③头孢他啶-阿维巴坦:按CLSI文件要求,头孢他啶-阿维巴坦对肠杆菌目细菌的抑菌圈直径为20~22 mm的菌株,需补充MIC试验确认,以避免报告假敏感或假耐药结果[2]。除上述三种抗菌药物外,实验室应尽可能开展其他抗菌药物的药敏试验,包括磷霉素、氯霉素和舒巴坦等,必要时开展联合药敏试验,以筛选其他有效的抗感染治疗方案[12],在尽量减少替加环素、多黏菌素和头孢他啶-阿维巴坦使用的同时,亦可减少经济困难患者的医疗开支。产生碳青霉烯酶是肠杆菌目细菌对碳青霉烯类耐药最主要的耐药机制[13],由于不同碳青霉烯酶抑制剂复合制剂对不同碳青霉烯酶的抑制作用不同,如阿维巴坦可抑制A类和D类丝氨酸碳青霉烯酶的活性,但不能抑制B类金属酶的活性;韦博巴坦和瑞来巴坦可抑制A类丝氨酸碳青霉烯酶的活性,但不能抑制B类金属酶和D类丝氨酸碳青霉烯酶的活性。因此,为应对碳青霉烯类耐药革兰阴性杆菌所致感染带来的挑战,实验室应按喻华等[14]倡议的《肠杆菌目细菌碳青霉烯酶的实验室检测和临床报告规范专家共识》开展碳青霉烯酶表型或基因型的检测并进行临床报告。
纵观2020年全国52所医院临床分离菌的分布特征和对受试抗菌药物的耐药性监测,显示我国临床分离菌中甲氧西林耐药菌(MRSA、MRSE和MRCNS)、万古霉素耐药肠球菌(VRE)、青霉素不敏感的肺炎链球菌(PNSP)、肠杆菌目细菌中产ESBL菌、大肠埃希菌中氟喹诺酮类耐药性(FQR)、碳青霉烯类耐药革兰阴性杆菌(CRO)等多年来仍以较高的检出率存在于我国临床,但监测资料同时也显示,多种重要耐药细菌的检出率呈连续下降趋势,提示加强细菌耐药性监测,结合医院感染预防控制措施,对控制耐药细菌的流行播散是行之有效的。为遏制细菌耐药性,必须继续加强细菌耐药性监测。
猜你喜欢
传染病信息(2022年2期)2022-07-15
广东医科大学学报(2020年4期)2020-08-24
天津医科大学学报(2019年3期)2019-08-13
广东饲料(2016年6期)2016-12-01
中国当代医药(2015年26期)2015-03-01
现代检验医学杂志(2015年4期)2015-02-06
现代检验医学杂志(2015年2期)2015-02-06
中国感染与化疗杂志(2015年5期)2015-01-23
郑州大学学报(理学版)(2014年2期)2014-03-01
现代检验医学杂志(2014年1期)2014-02-06
