
北方主栽花生品种的源库特征及其分类
2021-07-19 12:02刘兆新赵继浩潘小怡赖华江李向东杨东清
作物学报 2021年9期
高 芳 刘兆新 赵继浩 汪 颖 潘小怡 赖华江 李向东 杨东清
北方主栽花生品种的源库特征及其分类
高 芳 刘兆新 赵继浩 汪 颖 潘小怡 赖华江 李向东*杨东清*
山东农业大学农学院/ 作物生物学国家重点实验室, 山东泰安 271018
大田栽培条件下, 以中国北方主栽的13个花生品种为试验材料, 对单株叶面积、开花数、成果率等18个源库性状进行测定和计算, 利用主成分分析、聚类分析等统计方法, 筛选花生源库性状评价指标, 对比不同品种的源库性状差异和产量差异, 并进行源库类型划分。结果表明, 结荚期和饱果期的叶面积、开花数量、成果率和荚果充实度可以作为评价花生源库关系的主要指标。根据源库特征及产量表现可以把花生品种分为源库协调型、源大库小型、源足库少型和源足库多型4类。源库协调型品种叶片净同化率高, 花期持续时间和开花数量适中, 有效果比例和荚果充实度高, 易获得高产。源大库小型品种叶面积过大, 叶片净同化率和单位叶面积荚果产量低, 荚果充实度低, 限制产量提高。源足库多型品种产量限制因素为花期长, 花数多, 成果率低, 无效果针和荚果消耗营养, 有效荚果饱满度不足。源足库少型品种产量限制因素为开花持续时间短, 花量少。因此, 在花生生产中, 应该针对不同源库类型品种, 采取相应措施控制叶源大小和开花量, 防止叶源冗余、花多不实和果多不饱, 提高有效果比例和荚果饱满度, 增加荚果产量。
花生; 源; 库; 评价指标; 产量
近代作物栽培生理研究中常用源、库理论来阐明作物产量形成的规律。作物产量既取决于光合源同化物的生产能力, 又取决于库的大小和库容能力[1]。协调的源库关系对提高作物产量有重要意义[2], 有学者认为, 更高的生物量产量和优化的源库比将推动作物产量的下一次巨大飞跃[3], 但关于源库在产量形成中相对作用的大小, 各国学者的研究结果尚不一致。有研究认为, 大库容类型品种产量较高, 小库容类型产量较低, 产量与库容量的关系极为密切[4]; 也有研究认为, 作物产量与生殖生长期叶面积指数呈极显著正相关[5]; 多数研究结果认为, 协调好源库关系, 扩源增库才能保证作物的高产[6-8]。另外在相同生态环境及栽培条件下, 不同品种源库特征表现不一[9], 说明作物的源库关系存在明显的基因型差异, 因此可将品种按照源库关系进行分类研究。
在水稻中, 前人按照不同的源库特征指标, 把水稻现有品种分为源限制型、库限制型和源库互作型或库大源强协调型、库小源小协调型、库大源弱源限制型、库小源足库限制型[10-11], 这对花生源库关系的研究提供了一定参考。与禾谷类作物不同, 花生具有无限开花结实特性, 单株开花数量多, 持续时间长, 但成针、成果率低, 且饱果率变幅大(20%~80%)[12], 冗余的无效荚果库是限制花生产量提高的主要因素, 另外, 花生又具有无限生长习性和发生分枝的特性[13], 生育前期叶源展开过快、叶片数量多而冗余, 生育后期叶片早衰, 脱落速度快。叶片的前期冗余和后期过早衰老也是花生生产中难以解决的关键问题。叶源与荚果库关系不协调已成为花生产量提高和品质改善的瓶颈。关于花生源库关系及其对产量品质的影响, 已有研究报道, 但研究内容多采用单一或几个品种, 且对花生源库性状的评价指标和评价标准单一, 无统一结论。本研究以北方13个主推花生品种为试验材料, 从多个源库指标中筛选主要评价因子, 阐明描述花生源库特征的表观指标, 并对花生品种进行源库特征分类, 以期为花生的高产优质栽培及新品种选育提供理论基础和技术指导。
1 材料与方法
1.1 供试材料
收集近几年北方主栽大花生品种13个, 分别来自于山东农业大学、河北省农林科学院、中国农业科学院油料作物研究所等8个育种单位。品种名称及来源见表1。

表1 供试花生品种及来源
1.2 试验设计
大田试验于2018—2019年在山东省泰安市山东农业大学农学实验站(36°09′N, 117°09′E)进行, 该地属温带大陆性季风气候, 2018、2019年全年平均气温均为14.6℃, 花生生育期降水量分别为531.1 mm和365.3 mm。试验田为冬闲田沙壤土, 2018年试验田0~20 cm耕层土壤含碱解氮45.3 mg kg-1、速效磷45.6 mg kg-1、速效钾59.4 mg kg-1、有机质10.6 g kg-1, 2019年试验田0~20 cm耕层土壤含碱解氮82.9 mg kg-1、速效磷51.4 mg kg-1、速效钾64.7 mg kg-1、有机质14.8 g kg-1。花生种植前基施复合肥750 kg hm-2,深耕、耙平、起垄(垄距0.9 m, 每垄种植2行, 垄面行距0.3 m), 覆膜种植。各品种按小区种植, 小区面积2.7 m × 5 m, 穴距16 cm, 每穴2粒, 采取随机区组排列, 每品种重复3次。花生生长期内田间管理同一般高产田, 及时浇水、排涝, 除草、防治虫害。2018年5月6日播种, 9月15日收获; 2019年5月7日播种, 9月15日收获。
1.3 测定指标及数据计算
1.3.1 叶面积和干物重 分别于开花后20 d开始每隔10 d取各品种长势一致植株3棵, 采用LI-3100C台式叶面积仪(Li-Cor, USA)测定单株叶面积(leaf area, LA), 于花后30 d (花针期)、50 d (结荚期)、80 d (饱果期)取各品种长势一致植株3棵, 记录单株果针数、幼果数、秕果数、饱果数, 并按根、茎、叶、果分样, 105℃杀青30 min、80℃烘干至恒重后测定不同器官的干物质重量。
1.3.2 开花数、成针率、成果率 各品种挂牌标记长势相近的连续10穴植株, 现花后(6月6日左右)于每日清晨人工计数标记植株当日新开花数量, 直至平均单株开花数连续5 d<0.5。收获期时, 按穴收获, 并记录果针数、幼果数、秕果数、饱果数, 计算成针率、成果率、有效果比例。
1.3.3 产量及构成因素 成熟期, 每小区收获2.7 m × 2.0 m, 晾干, 去杂, 称重。室内测定各品种百果重、千克果数、出仁率。
1.3.4 数据计算 荚果生物量指果针、幼果、秕果、饱果生物量之和; 有效果数指秕果、饱果数量之和, 总果数指幼果、秕果、饱果数量之和;
式中荚果生物量指果针、幼果、秕果、饱果生物量之和;
式中有效果数指秕果、饱果数量之和, 总果数指幼果、秕果、饱果数量之和;
式中, LAIa、LAIb指2个时期叶面积指数, Wa、Wb指2个时期干物质量, Ta、Tb指2个时期相差天数。
1.4 数据处理
采用SPSS 23软件对源库相关性状进行主成分分析和聚类分析; 采用DPS 7.05软件最小显著极差法(LSD)检验平均数显著性; 采用Microsoft Excel软件计算品种间源库指标标准差及变异系数; 采用SigmaPlot 10.0软件作图。
2 结果与分析
2.1 供试花生品种源库指标变异幅度
将花生源库指标按源性状、库性状、源库综合性状及产量性状进行分类, 并计算各指标变异范围和变异系数。由表2可知, 源性状指标中, 单株叶面积(leaf area, LA)和叶干物质(leaf dry matter, LDM)变异系数最大值均出现在结荚期。该时期不同品种单株叶面积变异幅度为1717.7~2910.9 cm2, 相差169.5%; 单株叶干物质的变异幅度为9.47~ 15.63 g, 相差165.20%。库性状指标中, 成果率的变异系数最大, 变异幅度为8.66%~24.20%, 相差279.4%; 其次是单株开花数, 变异幅度为67~124朵, 相差184.0%; 开花持续时间的变幅为29~46 d, 相差158.6%。源库综合性状方面, 单位叶面积负荷量和单位叶面积对产量贡献的变异系数都比较大, 变幅相差分别为274.1%和280.0%, 收获指数变异系数为12.14%, 变幅相差150.6%。产量性状方面, 荚果充实度和有效果比例的变异系数分别为19.63%和16.1%, 产量变异系数为7.88%, 变异幅度为6024.69~7944.44 kg hm-2, 相差131.9%。说明供试的13个品种源库差异较大, 可以代表不同源库类型花生品种。
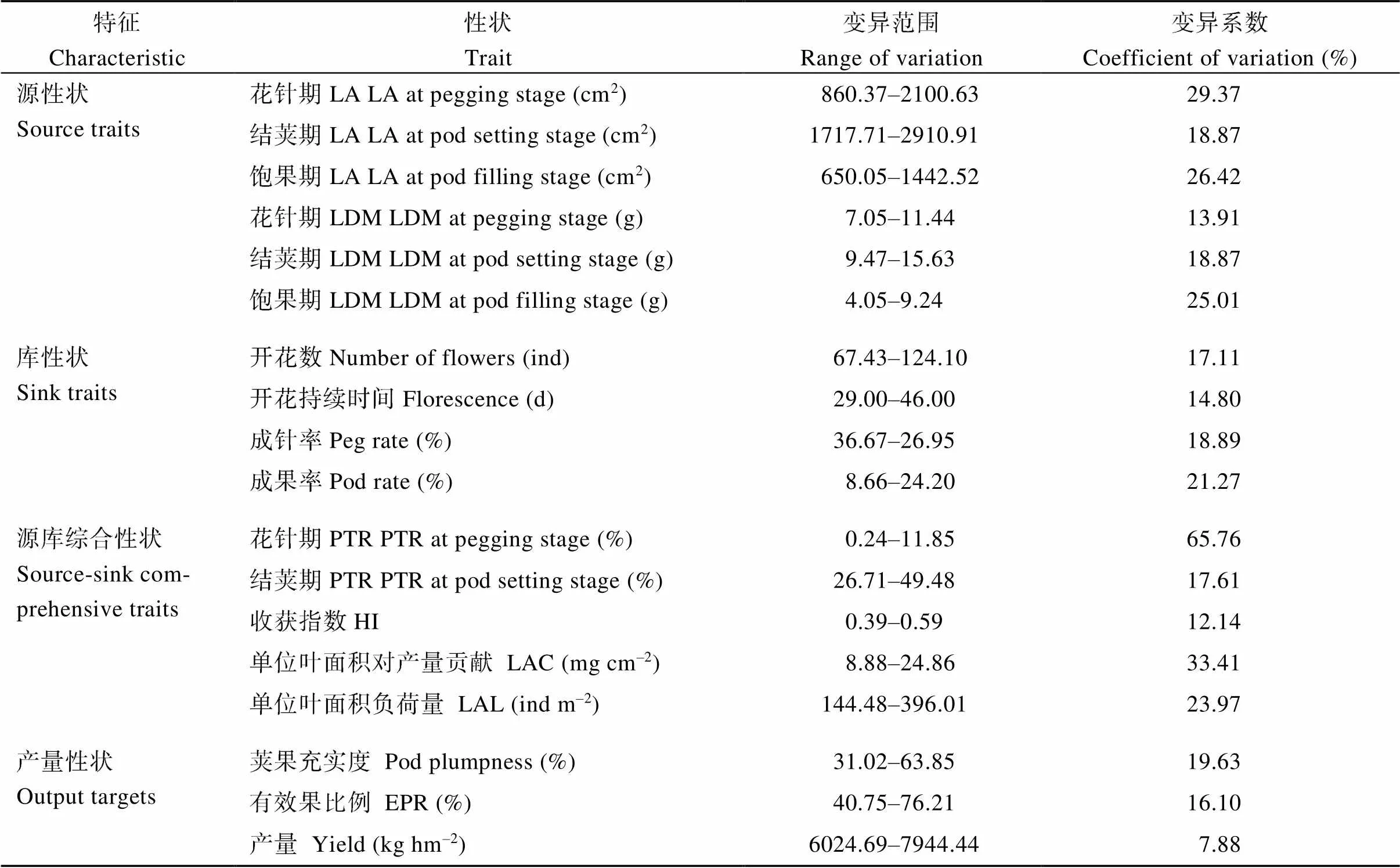
表2 花生各源、库指标变异幅度(2019年)
表中源、库性状数据为单株性状数据。LA: 叶面积; LDW: 叶干物质; PTR: 荚果生物量比例; HI: 收获指数; LAC: 单位叶面积对产量贡献; LAL: 单位叶面积负荷量; EPR: 有效果比例。
The source and sink traits in the table is the data of per plant. LA: leaf area; LDW: leaf dry matter; PTR: pod biomass/total biomass; HI: harvest index; LAC: unit-leaf-area contribution to yield; LAL: unit-leaf-area load; EPR: economic pod ratio.
2.2 花生品种源库性状主成分分析
利用主成分分析法, 对叶面积、叶干物质等18个花生源库性状进行降维分析, 由表3可以看出, 第1主成分的特征值是5.155, 贡献率为28.640%; 第2主成分的特征值是2.935, 贡献率为16.305%; 第3主成分的特征值是2.915, 贡献率为16.197%; 第4主成分的特征值是2.563, 贡献率为14.239%; 第5主成分的特征值是2.397, 贡献率为13.315%。前5个特征向量的累计贡献率为88.7%, 说明这5个主成分可以反映出18个源库性状的绝大部分信息, 因此可以选取这5个主成分作为花生源库性状的综合评价指标。
由表4可以看出, 第1主成分的性状中结荚期叶面积的特征向量值最大, 结荚期叶干物质次之, 其特征向量与叶面积接近。生产中常用叶面积和叶干物质作为源的评价指标, 因此可以称第1主成分为前期源因子, 当第1主成分的各特征向量值高时, 单位叶面积对产量的贡献有降低趋势。第2主成分的性状中荚果充实度的特征向量值最大, 有效果比例次之。荚果充实程度和有效果比例可以影响产量的高低, 因此可以称第2主成分为产量因子。第3主成分的性状中单株开花数的特征向量值最大, 开花持续期次之。花生单株开花数量及开花持续期是荚果库形成的第1阶段, 可以称第3主成分为前期库因子。当第3主成分的各向量值高时, 成果率及有效果比例有下降趋势。第4主成分的性状中饱果期叶面积的特征向量值最大, 饱果期叶干物质次之。可以称第4主成分为后期源因子。第5主成分的性状中成针率特征向量值最大, 成果率次之, 成针率和成果率是荚果库形成的后2个阶段, 因此可以称第5主成分为后期库因子。表明花生源库性状的主成分因子包含源因子、库因子、产量形成因子, 能较全面地评价花生源库关系, 对花生源库评价体系的建立提供理论依据。

表3 花生源库性状主成分分析的方差贡献率

表4 花生源库性状的主成分载荷矩阵
缩写同表2。Abbreviations are the same as those given in Table 2.
2.3 花生品种源库性状主成分得分
根据主成分因子的特征值和相应的特征向量, 计算13个品种的主成分得分。从表5可以看出, 13个品种中, 冀花5号和潍花8号前期源因子得分较高, 花育33号和花育36号前期源因子得分较低; 冀花5号和潍花8号的产量因子得分最高, 山花9号和日花1号的产量因子得分较低; 山花9号的前期库因子得分最高, 花育36号的前期库因子得分最低; 日花1号、山花9号和丰花1号的后期源因子得分较高, 冀花18155的后期源因子得分最低; 中花24号的后期库因子得分最高, 日花1号的后期库因子得分最低。从综合性状方面看, 冀花5号综合主成分得分最高, 其次是潍花8号,花育36号综合主成分得分最低。
2.4 花生品种聚类分析
利用表5中各品种的5个主成分得分进行K-means聚类分析, 将供试花生品种聚分为4类, 结果如表6所示。第I类有2个品种冀花5号和潍花8号; 第II类有4个品种, 分别为潍花16号、冀花18155、中花24、豫花15号; 第III类有3个品种, 分别为丰花1号、山花9号、日花1号; 第IV类有4个品种, 分别为青花7号、花育33号、花育36号、豫花9326。根据聚类分析结果, 选择各类中代表性品种进行源库性状及产量性状比较, 以明确源库性状对花生产量形成的影响。代表性品种分别为冀花5号、潍花8号、中花24、山花9号、日花1号、花育33号、花育36号。
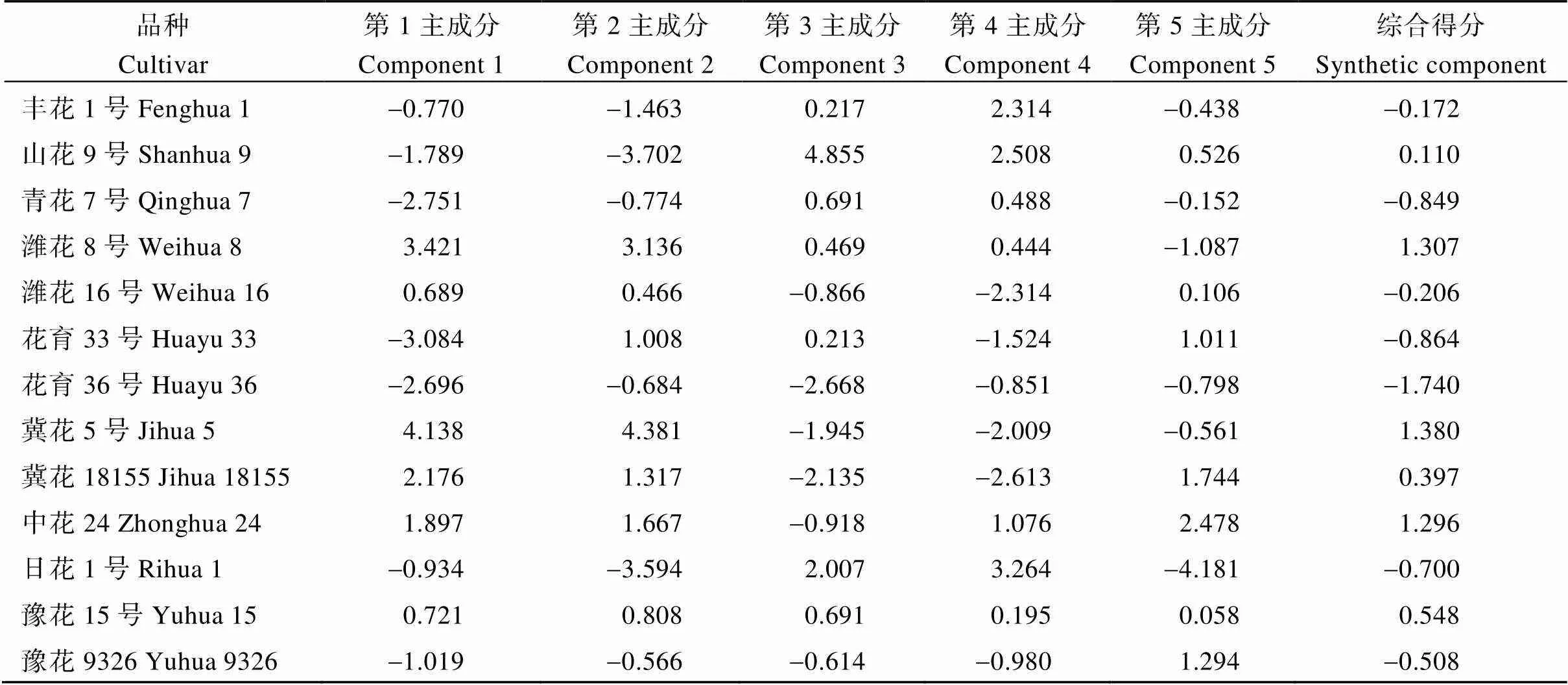
表5 主成分得分
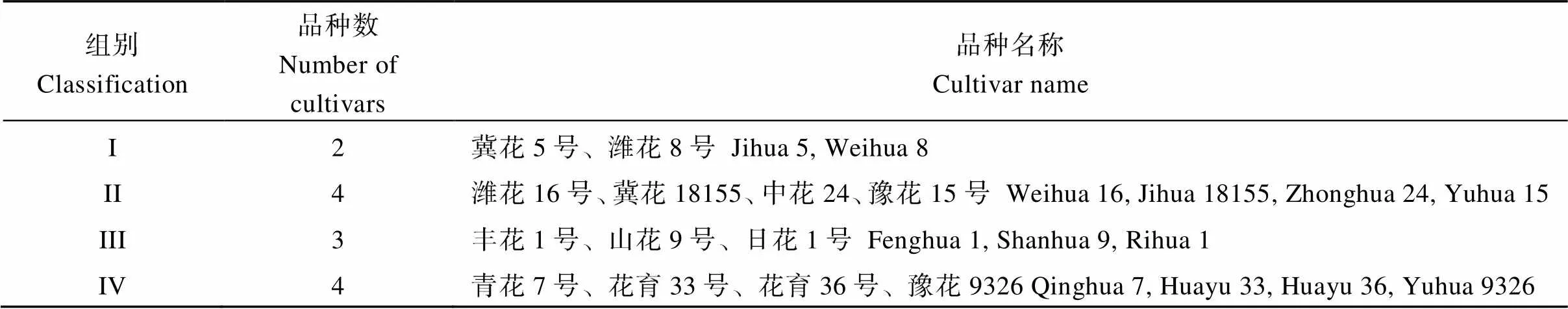
表6 利用K-means动态聚类分析法划分源库类型
2.5 花生品种间源性状差异
2.5.1 叶面积差异 由图1可以看出, 自花后30 d开始, 不同类型花生品种单株叶面积开始出现差异, 花育33号和花育36号的单株叶面积显著高于其他品种。2年数据平均值显示, 二者单株叶面积最大值2788 cm2, 为冀花5号和潍花8号的1.8倍。之后随着生育期的推进, 基部落叶数量开始增多, 叶片的落叶速度超过新叶展开速度, 花生单株叶面积日益减少。花育33号和花育36号的叶面积日消亡率平均值为47.9 cm2d-1, 显著大于其他品种, 其他品种间叶面积日消亡率差异不大。
2.5.2 净同化率差异 结荚期是花生营养生长和干物质积累的最盛期, 也是荚果形成的关键期。由表7可以看出, 2018年冀花5号和潍花8号的结荚期净同化率平均值为5.4 g m-2d-1, 显著高于其他品种, 花育33号和花育36号的平均值为3.3 m-2d-1。所有品种中, 潍花8号的净同化率最高, 花育33号最低, 两者相差75.2%。2019年冀花5号和潍花8号的净同化率平均值为6.3 g m-2d-1, 分别比山花9号和中花24号高18.6%和32.9%, 比花育33号和花育36号的平均值高76.7%。2年结果说明, 花生净同化率与叶面积大小有关, 随着叶面积的增大, 净同化率呈下降趋势。
2.5.3 单位叶面积对产量贡献差异 由表7可以看出, 潍花8号的单位叶面积荚果产量最高, 其次是冀花5号, 2018年二者单位叶面积荚果产量平均值为21.7 mg cm-2, 2019年为24.1 mg cm-2, 均显著高于其他品种。花育33号和花育36号的单位叶面积荚果产量最低, 2018年平均值为10.9 mg cm-2, 2019年为10.4 mg cm-2。说明不同类型品种间单位叶面积对产量贡献的差异较大, 随着叶面积的增大单位叶面积对产量的贡献呈减小趋势。
2.6 花生品种间库性状差异
2.6.1 开花特性差异 由表8可以看出, 不同花生品种单株开花数量和开花持续时间差异较大。在供试品种中山花9号单株开花数量最多, 总开花数为124, 其次是日花1号, 单株总开花数99。花育36号和中花24单株开花数量最少, 分别为70和75, 较山花9号分别减少了43.5%和39.5%。不同类型品种开花持续时间也存在差异, 山花9号和日花1号开花持续时间最长, 花期平均持续44 d; 其次是潍花8号和冀花5号, 花期平均持续33 d; 花育36和中花24开花持续时间最短, 花期平均持续分别为29 d和31 d。
同列不同字母表示不同品种间差异显著(< 0.05)。
Different letters in the same column indicate significant differences at< 0.05 among peanut cultivars.
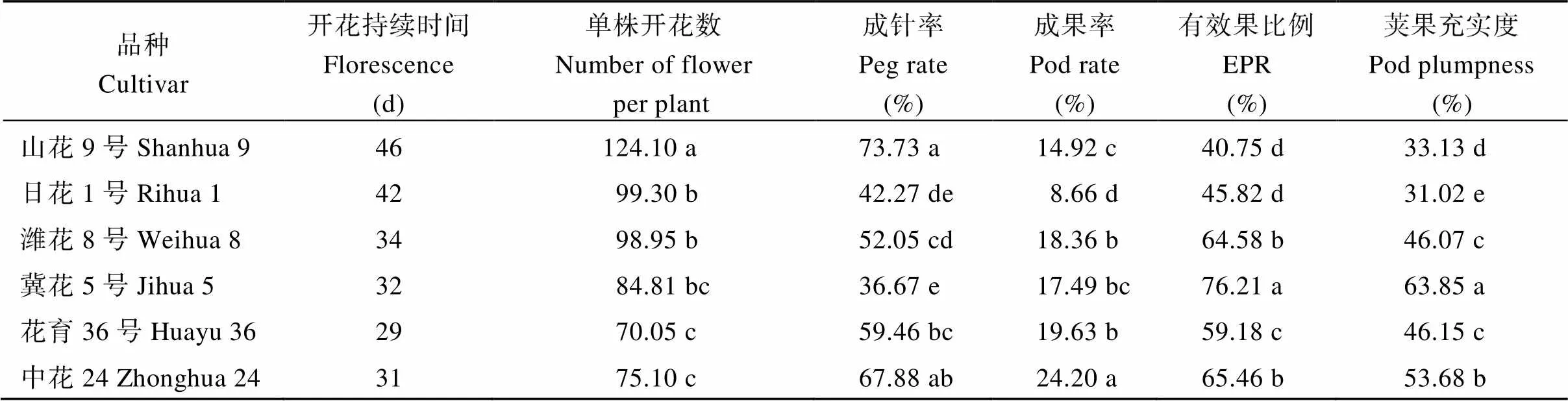
表8 不同花生品种库性状差异(2019年)
同列不同字母表示不同品种间差异显著(< 0.05)。
Different letters in the same column indicate significant difference at< 0.05 among peanut cultivars. EPR: economic pod ratio.
2.6.2 结荚特性差异 由表8可以看出, 山花9号和中花24成针率较高, 日花1号和冀花5号成针率较低。中花24和花育36号成果率较高, 分别为24.2%和19.6%; 潍花8号和冀花5号平均成果率为18.0%; 山花9号和日花1号成果率较低, 平均成果率为11.8%。说明成针率无类型间显著差异, 而成果率存在明显的类型间差异, 山花9号和日花1号的成果率显著低于其他品种。
花生收获时的产量是由秕果和饱果重量构成的,因此秕果和饱果称为有效果, 幼果没有经济价值, 其重量不能计入产量中, 称为无效果。由表8可以看出, 冀花5号有效果比例最高, 其次是中花24和潍花8号, 山花9号和日花1号有效果比例较低, 分别比冀花5号显著减少了46.5%和39.9%。荚果的充实度能反映源同化产物分配到荚果的份额, 也能反应品种荚果的饱满程度, 预估品种的增产潜力。供试品种中, 冀花5号荚果充实度最高, 其次是中花24。山花9号和日花1号的充实度最低, 平均充实度仅为32.1%, 显著低于其他品种。
2.7 花生品种间产量性状差异
以供试品种平均产量为对照, 对比13个品种的产量情况。由表9可以看出, 2018年, 冀花5号产量较平均产量显著增加了20.8%, 位居供试品种第1位; 冀花18155、山花9号、潍花8号、豫花9326、潍花16号、青花7号产量均高于平均产量; 花育36号产量显著低于平均产量。2019年, 冀花5号产量较平均产量显著增加了13.4%, 冀花18155产量较平均产量显著增加了7.7%, 其他高于平均产量的品种分别为: 中花24、潍花8号、豫花9326、山花9号、花育33号; 花育36号和日花1号产量显著低于平均产量。

表9 不同类型花生品种产量差异
同列不同字母表示不同品种间差异显著(< 0.05)。
Different letters in the same column indicate significant differences at< 0.05 among peanut cultivars .
2.8 花生源库类型划分
根据源库性状特征和产量差异, 对供试花生品种按聚类分组结果进行源库类型划分。第I组中2个品种的第1主成分得分、第2主成分得分及综合主成分得分均高于其他品种, 综合性状优良。其源库特征为单株叶面积较小, 净同化率高, 单株结果数多, 荚果充实度高, 产量表现突出, 属于源库协调型品种; 第II组中4个品种的第1主成分和第2主成分得分仅次于第I组, 综合性状较好。源库特征为叶面积大小适中, 净同化率较高, 但开花持续时间短, 单株开花数量少, 属于源足库少型品种; 第III组中3个品种的第3主成分得分最高, 第2主成分得分最低。源库特征为开花持续时间和单株开花数高于其他品种, 但无效果过多, 有效果比例和荚果充实度低, 属于源足库多型品种: 第IV组中4个品种的第1主成分得分和综合主成分得分最低, 第2主成分得分较低, 综合性状较差。源库特征为叶面积最大, 但净同化率低, 荚果充实度低, 属于源大库小型品种。
3 讨论
3.1 花生源库性状衡量指标
“源库理论”概念是Mason和Maskell在1928年首次提出的。这一理论的提出初步分析了作物产量的形成, 也在一定程度上弥补了光合理论的不足, 同时为作物获得高产提供了新思路和新方法。建立源库关系评价体系, 运用源库理论分析作物产量形成过程, 有助于客观评价每个品种的产量潜力, 从而指导育种工作与栽培实践[14]。前人已经研究了玉米、小麦、水稻、棉花等作物的源库特征特性, 也取得了不少学术成果[15-20], 提出了一系列关于衡量源库关系的指标。董桂春等[21]综合前人的研究结果将单位叶面积籽粒产量、单位叶面积库容量、单位干物质库容量、单位氮素库容量均定义为水稻源库比指标; 徐富贤等[22]提出叶容比(叶重/库容量)可以反映籽粒库的充实度; 凌启鸿[23]用实粒/叶, 粒重/叶反映水稻群体协调指标。本研究选用了13个北方主栽大花生品种为研究对象, 调查了18个表型性状并进行主成分分析, 明确了评价花生源库关系的5个主要因子, 以各因子中特征向量值大的表型特征描述花生的源库特征。5个主要因子分别为前期源因子, 以结荚期叶面积的特征向量值最大; 其次是产量形成因子, 以荚果充实度的特征向量值最大; 其余分别为前期库因子(开花数)、后期源因子(饱果期叶面积)和后期库因子(成针率、成果率)。5个主因子对源库关系特征的累积贡献率为88.7%, 能较全面的评价花生的源库特征。
3.2 源性状对产量形成的影响
植物叶片的光合产物是生长发育和产量形成主要物质基础。石庆华等[26]认为, 库大源不足是籼粳杂交稻结实率低的主要原因, 源是影响其产量进一步提高的主要因素; Venkatraman等[30]认为, 在现在大气环境下, 要增加作物产量应该减少叶面积而不是增加叶面积, 较低叶面积就能满足作物生长中对同化物积累的需要; 王夫玉等[24]认为, 水稻抽穗后要在适宜的绿叶面积基础上, 尽可能延长光合叶片的寿命, 提高籽粒充实度。大量试验和生产实践证明, 在适宜的叶面积范围内, 降低生育后期叶面积下降速率, 延长叶面积盛期持续的时间, 是增加花生荚果产量的一条有效途径[31]。本研究中源库协调型品种结荚期单株叶面积最小, 但其单位叶面积对荚果产量的贡献显著大于其他品种, 荚果产量也显著高于平均产量, 而源大型品种单位叶面积对荚果产量的贡献小, 产量低于平均产量。说明本试验供试品种中不存在因为源不足而导致产量降低的现象,相反过大的叶面积容易造成叶片间的相互遮蔽, 增加叶面积日消亡率, 导致后期叶面积下降, 不利于产量的形成。Yoshida[25]通过在开花前和开花后提高CO2浓度以提高水稻产量这一试验推断水稻产量受源的光合能力限制; 崔菁菁等[27]认为, 进一步提高叶片净光合速率并改善叶片的光合能力应作为高产水稻品种选育的新目标。本研究也认为, 源库协调型品种产量高的原因是结荚期净同化率显著高于大源型和源中间型品种。
3.3 库性状对产量形成的影响
Ricardo等[34]研究表明, 花生单株总花数与开花持续时间呈负相关, 与成针率呈正相关; 王晓光等[35-36]等研究发现, 始花期较早, 花期较长的品种单株总开花量和有效花量较多, 始花期较晚, 花期较短的品种单株总开花量较少。本研究结果显示, 花期长的品种, 单株开花数量多, 花期短的品种单株开花数量少。不同类型品种间成针率无显著差异, 而成果率存在明显的类型间差异, 库多型品种成果率显著低于其他品种。在水稻生产上, 提高水稻产量的有效途径之一是增加单位叶干重的小穗数和籽粒大小[28-29]。陈琛等[32]研究表明, 水稻产量与库容量呈线性正相关, 随着库容量的增大产量亦上升, 大库容量水稻产量高。曹显祖等[33]研究表明, 水稻产量随着库(颖花量)的增加呈抛物线关系, 即产量先是随着库的增加而上升, 当库增加到一定程度时, 产量随着库的增强而下降。本研究中, 多库型品种虽然开花数量多, 但成果率低, 无效荚果和果针消耗过多的营养物质, 导致有效果比例和荚果的充实度均显著低于其他品种, 限制产量的提高; 库少型品种由于花期短, 单株开花数少, 虽然成果率最高, 但产量仍较低; 源库协调型品种尤其冀花5号, 其荚果产量最高的主要原因是有效果比例和荚果充实度显著高于其他品种。因此在育种和生产过程中, 应注意控制花生开花持续时间和开花数量, 花数过多或过少均影响产量的提高, 本研究中以花期32~34 d时产量表现最佳。控制花数的同时提高成果率, 减少无效荚果数量, 增加有效荚果比例和荚果充实度, 从而提高产量。
3.4 花生源库类型划分
在一定生态及栽培条件下, 按照作物的源库特征划分类型, 不仅可以丰富作物源库理论, 还可以为作物的因种栽培和新品种选育提供理论指导。徐富贤等[22]通过库源特征分析, 将长江上游杂交中稻高产栽培条件下, 按群体穗粒数进行分类。群体穗粒数小于160粒属增库增产型, 160~220粒属于库源互作型, 大于220粒属于增源增产型。江龙等[37]将水稻品种划分为增源增产型、增库增产型、源库互作增产型。姜文洙等[38]采用动态聚类分析方法将水稻划分为源库互作型、增库增产型、增源增产型和源库饱和型4种库源类型。在本研究中选择了不同源、库类型的代表性品种, 将源库性状与产量性状结合, 分析各品种的源库类型, 将花生品种分为源库协调型、源大库小型、源足库少型和源足库多4个类型。
4 结论
本试验供试花生品种源库性状差异较大。结荚期和饱果期叶面积、成针率、有效果比例、荚果饱满度可以用来描述花生品种的源库特性。根据源库特征可把花生品种分为源库协调型、源大库小型、源足库少型和源足库多型4类。源库协调型品种产量高的原因是叶片净同化率高, 后期叶面积消亡慢; 花期持续时间和开花数量适中, 有效果比例和荚果充实度高。源大库小型品种产量限制因素为叶面积过大, 叶片净同化率低。源足库少型品种产量限制因素为开花持续时间短, 花量少。源足库多型品种产量限制因素为花期长, 花数多, 成果率低, 无效果针和荚果消耗营养, 有效荚果饱满度不足。在花生生产中, 应该针对不同源库类型品种, 采取相应措施控制叶源大小和开花量, 防止叶源冗余、花多不实和果多不饱, 提高有效果比例和荚果饱满度, 增加荚果产量。
[1] 李向东, 张永丽. 农学概论. 北京: 中国农业出版社, 2017. pp 49–51.
Li X D, Zhang Y L. General Introduction Agronomy. Beijing: China Agriculture Press, 2017. pp 49–51 (in Chinese).
[2] Wang Y, Pang Y L, Chen K, Zhai L Y, Shen C C, Wang S, Xu J L. Genetic bases of source-, sink-, and yield-related traits revealed by genome-wide association study in Xian rice., 2020, 8: 119–131.
[3] Senthold A, Belay T K, Marcelo H L, Amador C, Daniel F C. Simulating the impact of source-sink manipulations in wheat., 2017, 202: 47–56.
[4] 董桂春, 李进前, 董燕萍, 周娟, 田昊, 于小凤, 张传胜, 张岳芳, 余龙. 产量构成因素及穗部性状对籼稻品种库容的影响. 中国水稻科学, 2009, 23: 523–528.
Dong G C, Li J Q, Dong Y P, Zhou J, Tian H, Yu X F, Zhang C S, Zhang Y F, Yu L. Effects of yield components and panicle traits on sink potential in conventional indica rice., 2009, 23: 523–528 (in Chinese with English abstract).
[5] 田艺心, 高凤菊, 曹鹏鹏, 王乐政, 华方静, 王士龄, 田文铎. 大豆叶面积指数、干物质积累分配与产量的关系. 山东农业大学学报(自然科学版), 2018, 49: 750–754.
Tian Y X, Gao F J, Cao P P, Wang L Z, Hua F J, Wang S L, Tian W D. Relationship between leaf area index, dry matter accumulation distribution and yield of soybean.(Nat Sci Edn), 2018, 49: 750–754 (in Chinese with English abstract).
[6] Lyu X M, Zhang Y, Zhang Y X, Fan S J, Kong L G. Source-sink modifications affect leaf senescence and grain mass in wheat as revealed by proteomic analysis., 2020, 20: 257.
[7] Sonnewald U, Fernie A R. Next-generation strategies for understanding and influencing source-sink relations in crop plants., 2018, 43: 63–70.
[8] Lucas B, Gustavo A S, Marıa E O. Seed dry weight response to source-sink manipulations in wheat, maize and soybean: a quantitative reappraisal., 2004, 86: 131–146.
[9] 姜慧芳, 任小平. 我国栽培种花生资源农艺和品质性状的遗传多样性. 中国油料作物学报, 2006, 28: 421–426.
Jiang H F, Ren X P. Genetic diversity of peanut resource on morphological characters and seed chemical components in China., 2006, 28: 421–426 (in Chinese with English abstract).
[10] 王余龙, 蔡建中, 徐永林, 华鹤良. 水稻籽粒受容活性及其控制途径: I. 籽粒含水率与受容活性的关系. 江苏农学院学报, 1990, 11(3): 25–29.
Wang Y L, Cai J Z, Xu Y L, Hua H L. Activity of grain capacity and its regulation in rice: I. The relationship between grain water content and capacity activity., 1990, 11(3): 25–29 (in Chinese with English abstract).
[11] 曹显祖, 朱庆森. 水稻品种的源库特征及其类型划分的研究. 作物学报, 1987, 13: 265–272.
Cao X Z, Zhu Q S. Study on the characteristics of source and sink of rice varieties and their types., 1987, 13: 265–272 (in Chinese with English abstract).
[12] 嵇军德. 花生品种抗旱性与开花结实特性的关系. 山东农业大学硕士学位论文, 山东泰安, 2015.
Ji J D. The Relationship between Drought Resistance and the Flowering and Podding Characteristics of Peanut Varieties. MS Thesis of Shandong Agricultural University, Tai’an, Shandong, China, 2015 (in Chinese with English abstract).
[13] 王丽丽. 改变源-库比对花生籽仁品质和叶片衰老的影响. 山东农业大学硕士学位论文, 山东泰安, 2005.
Wang L L. The Effect of Altering Source-sink Ratio on Kernel Quality and Leaf Senescence in Peanut. MS Thesis of Shandong Agricultural University, Tai’an, Shandong, China, 2005 (in Chinese with English abstract).
[14] 王丰, 张国平, 白朴. 水稻源库关系评价体系研究进展与展望. 中国水稻科学, 2005, 19: 556–560.
Wang F, Zhang G P, Bai P. Achievement and prospects of research on evaluation of the relationship between source and sink in rice., 2005, 19: 556–560 (in Chinese with English abstract).
[15] 陈琛, 张家星, 李万元, 唐东南. 氮高效水稻主要源库性状的基本特点及其调控. 中国水稻科学, 2017, 31: 185–194.
Chen C, Zhang J X, Li W Y, Tang D N. Fundamental features of source-sink characters and their regulation in high nitrogen efficiency rice lines., 2017, 31: 185–194 (in Chinese with English abstract).
[16] 葛勇. 基于库源关系的中棉所63杂种优势机理研究. 中国农业科学院硕士学位论文, 北京, 2016.
Ge Y. Study on Heterosis Mechanism of CCRI63 Based on Sink-source Relationship. MS Thesis of Chinese Academy of Agricultural Sciences, Beijing, China, 2016 (in Chinese with English abstract).
[17] 刘星, 邱慧珍, 张文明, 张春红, 王亚飞, 马兴. 连作马铃薯植株库源关系及其对块茎产量的调节机理. 应用生态学报, 2017, 28: 1571–1582.
Liu X, Qiu H Z, Zhang W M, Zhang C H, Wang Y F, Ma X. Sink-source relationship of potato plants and its role involved in the reduction of tuber yield in continuous cropping system., 2017, 28: 1571–1582 (in Chinese with English abstract).
[18] 宁运旺, 马洪波, 张辉, 汪吉东, 许仙菊, 张永春. 甘薯源库关系建立、发展和平衡对氮肥用量的响应. 作物学报, 2015, 41: 432–439.
Ning Y W, Ma H B, Zhang H, Wang J D, Xu X J, Zhang Y C. Response of sweetpotato in source-sink relationship establishment, expanding, and balance to nitrogen application rates, 2015, 41: 432–439 (in Chinese with English abstract).
[19] Wei H H, Meng T Y, Li X Y, Dai Q G, Zhang H C, Yin X Y. Sink-source relationship during rice grain filling is associated with grain nitrogen concentration., 2018, 215: 23–38.
[20] Fernie A R, Bachem C W B, Helariutta Y, Neuhaus H E, Prat S, Ruan Y L, Stitt M, Sweetlove L J, Tegeder M, Wahl V, Sonnewald S, Sonnewald U. Synchronization of developmental, molecular and metabolic aspects of source-sink interactions., 2020, 6: 55–66.
[21] 董桂春, 田昊, 张彪, 李进前, 于小凤, 王余龙. 不同库容量类型籼稻品种源库相关参数的基本特点. 作物学报, 2009, 35: 1900–1908.
Dong G C, Tian H, Zhang B, Li J Q, Yu X F, Wang Y L. Characteristics of source-sink related parameters in conventionalrice cultivars with different types of sink potential., 2009, 35: 1900–1908 (in Chinese with English abstract).
[22] 徐富贤, 熊洪, 张林, 朱永川, 刘茂, 蒋鹏, 郭晓艺. 南方稻区杂交中籼稻高产品种的库源结构及其优化调控规律研究进展. 中国生态农业学报, 2016, 24: 1285–1299.
Xu F X, Xiong H, Zhang L, Zhu Y C, Liu M, Jiang P, Guo X Y. Research progresses and prospects of sink-source structures and optimal regulation of high-yield varieties of hybrid rice in China., 2016, 24: 1285–1299 (in Chinese with English abstract).
[23] 凌启鸿. 作物群体质量. 上海: 上海科学技术出版社, 2000. pp 230–257.
Ling Q H. The Quality of Crop Population. Shanghai: Shanghai Scientific and Technical Publishers, 2000. pp 230–257 (in Chinese).
[24] 王夫玉, 黄丕生. 水稻群体源库特征及高产栽培策略研究. 中国农业科学, 1997, 30(5): 26–33.
Wang F Y, Huang P S. Study on source-sink characteristics and high-yield cultivation strategies of rice population., 1997, 30(5): 26–33 (in Chinese with English abstract).
[25] Yoshida S. Effects of CO2enrichment at different stages of panicle development on yield components and yield of rice (L.)., 1973, 19: 311–316.
[26] 石庆华, 徐益群, 张佩莲, 曾宪江, 潘晓华, 钟旭华. 籼粳杂交稻的氮素吸收特性及其对库源特征的影响. 江西农业大学学报, 1994, 16: 333–339.
Shi Q H, Xu Y Q, Zhang P L, Zeng X J, Pan X H, Zhong X H. Effects of N uptake on the sink and source ofF1hybrids rice., 1994, 16: 333–339 (in Chinese with English abstract).
[27] 崔菁菁, 徐克章, 石晶皎, 武志海, 陈展宇, 张治安, 吴春胜. 不同水稻品种库源的特征变化. 植物学报, 2015, 50: 699–705.
Cui Q Q, Xu K Z, Shi J J, Wu Z H, Chen Z Y, Zhang Z A, Wu C S. Changes in sink-source variation characteristics of different rice varieties., 2015, 50: 699–705 (in Chinese with English abstract).
[28] Huang L Y, Yang D S, Lia X X, Peng S B, Wang F. Coordination of high grain yield and high nitrogen use efficiency through large sink size and high post-heading source capacity in rice., 2019, 233: 49–58.
[29] Zhang H, Chen T, Liu L, Wang Z, Yang J, Zhang J. Performance in grain yield and physiological traits of rice in the Yangtze river basin of china during the last 60 yr., 2013, 12, 57–66.
[30] Venkatraman S, Praveen K, Stephen P L. Decreasing, not increasing, leaf area will raise crop yields under global atmospheric change., 2017, 23: 1626–1635.
[31] 万书波. 中国花生栽培学. 上海: 上海科学技术出版社, 2003. pp 286–288.
Wan S B. Peanut Cultivation in China. Shanghai: Shanghai Scientific and Technical Publishers, 2003. pp 286–288 (in Chinese).
[32] 陈琛, 于小凤, 赵步洪, 高英博, 舒小伟, 徐杰皎, 唐东南, 黄建晔, 王余龙, 姚友礼, 董桂春. 水稻大库容遗传群体源库性状、物质生产与分配的基本特征. 江苏农业学报, 2019, 35: 1009–1014.
Chen C, Yu X F, Zhao B H, Gao Y B, Shu X W, Xu J J, Tang D N, Huang J Y, Wang Y L, Yao Y L, Dong G C. Fundamental features of source-sink properties, biomass production and distribution in rice genetic populations with high sink capacity type., 2019, 35: 1009–1014 (in Chinese with English abstract).
[33] 曹显祖, 缪兴伯. 杂交中籼稻源库特征与高产栽培对策. 杂交水稻, 1989, 6(2): 4–6.
Cao X Z, Miu X B. Source-sink characteristics of hybridrice and high-yielding cultivation strategies., 1989, 6(2): 4–6 (in Chinese).
[34] Ricardo J H, Jorge B, María E O. Genetic improvement of peanut in Argentina between 1948 and 2004: links between phenology and grain yield determinants., 2015, 174: 12–19.
[35] 王晓光, 蒋春姬, 赵新华, 于海秋, 曹敏建. 辽宁省引进的花生高产品种开花特性. 中国油料作物学报, 2013, 35: 54–57.
Wang X G, Jiang C J, Zhao X H, Yu H Q, Cao M J. Flowering characteristics of high yield peanuts varieties introduced into Liaoning Province., 2013, 35: 54–57 (in Chinese with English abstract).
[36] 王晓光, 赵新华, 蒋春姬, 于海秋, 曹敏建. 辽宁省引进的花生高产品种结荚特性研究. 中国油料作物学报, 2013, 35: 400–404.
Wang X G, Zhao X H, Jiang C J, Yu H Q, Cao M J. Podding characteristics of high yielding peanut varieties introduced into Liaoning Province., 2013, 35: 400–404 (in Chinese with English abstract).
[37] 江龙, 韦宏恩, 杨昌达, 陈天华. 不同水稻品种的源库特性与产量形成的关系. 山地农业生物学报, 1998, 17: 318–322.
Jiang L, Wei H E, Yang C D, Chen T H. The research of the relationship between the source and sink characteristics of different kinds of rice and the formation of their output., 1998, 17: 318–322 (in Chinese with English abstract).
[38] 姜文洙, 吴涛, 唐曹甲子, 王彩芬, 马静, 强爱玲, 都兴林. 高产粳稻品种源库特征及评价体系的研究. 东北农业科学, 2019, 44(5): 1–4.
Jiang W Z, Wu T, Tang C J Z, Wang C F, Ma J, Qiang A L, Du X L. Studies on characteristics and evaluation of source-sink in high-yieldrice., 2019, 44(5): 1–4 (in Chinese with English abstract).
Source-sink characteristics and classification of peanut major cultivars in North China
GAO Fang, LIU Zhao-Xin, ZHAO Ji-Hao, WANG Ying, PAN Xiao-Yi, LAI Hua-Jiang, LI Xiang-Dong*, and YANG Dong-Qing*
College of Agronomy / State Key Laboratory of Crop Biology, Shandong Agricultural University, Tai’an 271018, Shandong, China
To explore whether and how source-sink theory was involved in mediating peanut high yield formation, a field experiment was conducted using 13 major peanut cultivars in North China. Principal component analysis and cluster analysis were used to evaluate natural indicators for the characterization and classification of source-sink types such as leaf area (LA) per plant, number of flowers, and pod rate. The results showed that LA per plant, number of flowers, pod rate, and pod plumpness at the pod-setting and pod-filling stages could be the major characteristics to evaluate source-sink relationship. Based on the source-sink indicators and yield components, these peanut cultivars could be classified into four categories, including source-sink coordination type, large source-few sink type, sufficient source-few sink type, and sufficient source-large sink type. The source-sink coordination type varieties could achieve high yield mainly due to the higher net assimilation rate at the pod-setting stage, the moderate florescence and number of flowers, and the higher economic pods ratio and pod fullness. Large source-few sink type cultivars had higher LA, but lower net assimilation rate and smaller contribution of the leaf area to the yield formation, thus decreased the pod yield. Sufficient source-large sink type cultivars had longer florescence and more flowers compared to others types, leading to reduce the pod rate and pod plumpness, which was the main limiting factor to increase the pod yield. On the contrary, both florescence and the number of flowers per plant of sufficient source-few sink type cultivars were less than others types. In summary, to prevent the redundancy of leaf source, more flowers but less pegs, and more pods but less kernels and to promote the effective proportions and pod fullness resulting in higher pod yield in peanut production, corresponding measures should be taken to control leaf area and amount of flowering according to different source-sink types peanut cultivars.
peanut; source; sink; evaluation indicator; yield
10.3724/SP.J.1006.2021.04164
本研究由国家重点研发计划项目“大田经济作物优质丰产的生理基础与调控”(2018YFD1000900), 山东省重大科技创新工程项目(2018YFJH0601-3)和山东省现代农业产业技术体系创新团队项目(SDAIT-04-01)资助。
This study was supported by the National Key Research and Development Program of China “Physiological Basis and Agronomic Management for High-quality and High-yield of Field Cash Crops” (2018YFD1000900), the Shandong Key Research and Development Program (2018YFJH0601-3), and the Innovation Team Project for Modern Agricultural Industrious Technology System of Shandong Province (SDAIT-04-01).
李向东, E-mail: lixdong@sdau.edu.cn; 杨东清, E-mail: chengyang2364@126.com
E-mail: 15563177918@163.com
2020-07-22;
2021-01-21;
2021-02-20.
URL: https://kns.cnki.net/kcms/detail/11.1809.S.20210220.1148.004.html
猜你喜欢
农学学报(2022年8期)2022-11-16
花生学报(2022年1期)2022-08-16
安徽农学通报(2022年6期)2022-04-07
江苏农业科学(2022年3期)2022-03-03
热带农业工程(2021年5期)2021-12-06
农业科技与装备(2021年4期)2021-08-30
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02
农民致富之友(2020年27期)2020-09-12
山东农业科学(2019年9期)2019-12-09
广西农学报(2019年1期)2019-07-18
