
甘草酸单铵盐对体外瘤胃发酵参数及甲烷产量的影响
2021-07-12 03:22:28蒋辰宇崔浩然白天天普宣宣刘俊峰郭雪峰
动物营养学报 2021年6期
蒋辰宇 崔浩然 白天天 普宣宣 刘俊峰,2 郭雪峰,2*
(1.塔里木大学动物科学学院,阿拉尔843300;2.畜牧科技兵团重点实验室,阿拉尔843300)
在过去25年中,随着人们对畜产品需求量的增加,加速了畜牧业的发展,导致反刍动物数量增多和温室气体排放量增加。甲烷排放量仅次于二氧化碳,但它在地球上的升温潜能约为二氧化碳的25倍[1]。其他研究表明,反刍动物的甲烷排放不仅造成2%~12%的能量损失,而且降低了饲料利用率并导致动物生产性能的降低和饲养成本增加[2]。因此,降低反刍动物的甲烷排放量不仅减少对环境的污染,并且还提高了反刍动物生产性能,减少饲料能量损失。有研究表明,皂苷作为植物次生代谢产物可以提高瘤胃发酵效率[3],通过直接或间接抑制产甲烷菌的活性,降低甲烷产生,此外,它还能提高菌体蛋白的合成效率,提高流入十二指肠的菌体蛋白产量[4]。皂苷是苷元为三萜或螺旋甾烷类化合物的一类糖苷,源于不同植物提取的皂苷在改善瘤胃发酵方式及效果上也不尽相同[5-8]。甘草酸单铵盐是甘草皂苷(又称甘草酸)的一种盐形式。甘草酸单铵盐的分子式为C42H65NO16,甘草皂苷的分子式为C42H62O16。由于甘草皂苷在常温中水溶性较低,为获得稳定、安全、方便的使用途径,通常使用水溶性较好的甘草酸盐,如甘草酸单铵盐、甘草酸二铵盐、甘草酸一钾盐等[9]。新疆是中国甘草的主要产区,具有非常丰富的甘草资源,若甘草酸单铵盐能够用于调控反刍动物瘤胃发酵,降低瘤胃甲烷产量,可为新疆地区丰富的甘草资源开发利用提供新的思路。因此,本试验研究不同浓度的甘草酸单铵盐对绵羊体外瘤胃发酵参数及甲烷产量的影响,旨在为其作为植物提取物添加剂的应用提供参考数据。
1 材料与方法
1.1 试验材料
甘草酸单铵盐选购自新疆阿拉尔某甘草产业有限责任公司,为白色粉末,高效液相色谱法(HPLC)测定其纯度≥98%。
1.2 试验动物及饲粮
选取3只平均体重为(35±3) kg、安装永久性瘘管的卡拉库尔羊作为瘤胃液采集的试验动物。每日09:00和20:00分2次定量饲喂,自由饮水。根据《肉羊饲养标准》(NY/T 816—2004)[10]进行饲粮配制,饲粮以玉米和苜蓿颗粒为主要原料,精粗比为60:40,其组成及营养水平见表1。
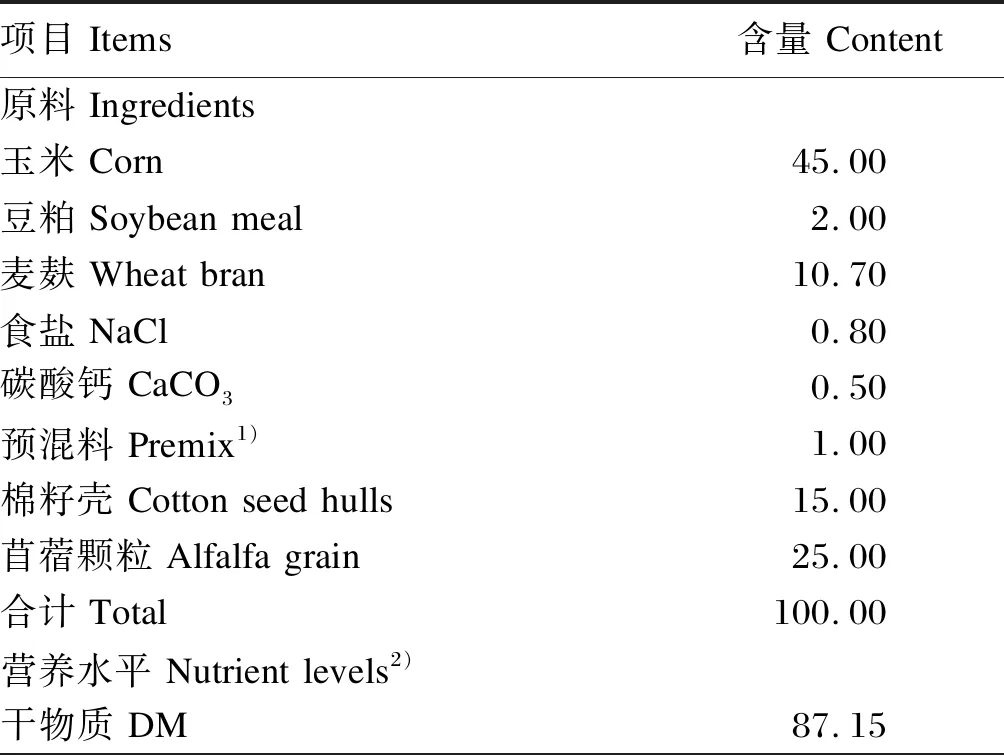
表1 饲粮组成及营养水平(风干基础)

续表1项目 Items含量 Content消化能 DE/(MJ/kg)13.61粗蛋白质 CP11.35钙 Ca1.14磷 P0.32非纤维性碳水化合物 NFC3)56.11中性洗涤纤维 NDF34.85非纤维性碳水化合物/中性洗涤纤维 NFC/NDF1.61
1.3 试验设计
体外培养以粉碎过40目后的基础饲粮为发酵底物,分别设置对照组、Ⅰ组、Ⅱ组、Ⅲ组、Ⅳ组和Ⅴ组,甘草酸单铵盐添加浓度分别为0、50、150、250、350、450 mg/L的,分别进行3、9、12、21、24 h的分批次体外培养,每个浓度的每个时间设置3个重复,测定其pH、总产气量、干物质降解率、氨态氮(NH3-N)浓度、挥发性脂肪酸(VFA)浓度及甲烷产量。
1.4 体外发酵培养方法
1.4.1 体外发酵装置
体外发酵装置为恒温水浴摇床(温度和摇晃频率均可调节),摇床上安装自制框架,用于固定玻璃注射器(100 mL),并使玻璃注射器的2/3浸入水浴中。玻璃注射器底端连接有自制可控制开关的阀门,用于抽取人工瘤胃液和收集发酵气体。
1.4.2 人工瘤胃液配置
瘤胃液采集:晨饲前2 h,采集3只绵羊瘤胃中不同位点的瘤胃液,并用40~60目4层尼龙布过滤,随后倒入预热达39 ℃的保温瓶中,盖严瓶口,迅速带回实验室并保温备用。
人工瘤胃液制备:人工唾液根据Menke等[11]的方法制备。备齐所需培养液,并按1∶2的比例向瘤胃液中加入提前通入二氧化碳并预热的人工唾液均匀混合,此过程持续通入二氧化碳。
1.4.3 体外发酵培养及样品的处理
准确称取发酵底物0.2 g及对应浓度的甘草酸单铵盐,将其放入注射器顶端,每个样品称取3个平行样。注射芯在插入注射器前应在其表面均匀涂抹凡士林,并小心插入注射器,防止发酵底物被吹出。将配制完的人工瘤胃液吸至注射器内,此过程持续通入二氧化碳,每支30 mL,关闭注射器阀门,置于39 ℃恒温数显水浴摇床中振荡发酵。记录不同发酵时间点的产气量,气体通过采血针及真空采血管进行收集,收集的气体用于测定甲烷产量,收集气体后的发酵液立即进行pH测定,随后将发酵液3 500 r/min离心10 min。取发酵液上清液分装入瓶,-20 ℃保存,用于测定NH3-N和VFA浓度。
1.5 指标测定
总产气量:通过读取注射器上的刻度测定。
pH:通过希玛牌便携式pH测试笔进行测定。
干物质降解率:按照《动物营养与饲料科学实践教程》[12]测定饲粮干物质含量并按公式计算降解率。
欧阳橘红调离滨湖先到南京,偶然的机会认识了部里一个司长,半年不到就结了婚,婚后就没回南京上班了。她先在化工部当工程师,那年国务院组建石油化工集团公司,从化工部分了一批人到集团公司,她也分到了集团公司,最后在集团副总工程师的位置退休,现在还兼了两所大学客座教授。
干物质降解率=100×(B-C)/B。
式中:B为待测样品中干物质的含量;C为待测样品尼龙袋残渣中干物质含量。
NH3-N浓度:参照冯宗慈等[13]方法进行测定。
VFA浓度:参照丁学智等[14]的方法,采用GC-2014气相色谱仪进行测定。测定条件为进样口温度130 ℃,检测器温度200 ℃,柱温220 ℃,柱流量0.6 mL/min。
甲烷产量:采用GC-2014气相色谱仪测定。测定条件为进样口温度120 ℃,检测器温度180 ℃,柱温80 ℃,柱流量1 mL/min。
1.6 数据处理
数据用SPSS 21.0的单因素方差分析进行显著性分析,用Duncan氏法和LSD法进行多重比较,以平均值±标准差表示结果。P<0.05为差异显著。
2 结果与分析
2.1 不同浓度甘草酸单铵盐对体外累积产气量的影响
由表2可知,随着发酵时间的增长,产气量逐渐增加,其中发酵初期产气量增长速度较快,且添加甘草酸单铵盐后的产气量增长速度高于对照组。但在发酵24 h后,Ⅲ组、Ⅳ组和Ⅴ组的产气量均显著低于对照组(P<0.05)。Ⅰ组和Ⅱ组在发酵24 h后的产气量略大于对照组,但差异不显著(P>0.05)。发酵24 h后产气量最低组为Ⅴ组,产气量为47.93 mL;产气量最高组为Ⅱ组,产气量为64.83 mL。

表2 不同浓度甘草酸单铵盐对体外产气量的影响
2.2 不同浓度甘草酸单铵盐对发酵液pH的影响
由表3可知,随着发酵时间的增长,各组pH呈逐渐下降的趋势。不同浓度的甘草酸单铵盐处理后,各试验组pH均有所下降。在发酵3 h后,各试验组pH均显著低于对照组(P<0.05);发酵9 h后,Ⅲ组、Ⅳ组、Ⅴ组的pH显著低于对照组(P<0.05),其他试验组与对照组相比差异不显著(P>0.05);发酵12 h后,各试验组pH均低于对照组,其中Ⅰ组、Ⅱ组、Ⅲ组、Ⅳ组与对照组相比差异显著(P<0.05);在发酵21 h后,各试验组pH均显著低于对照组(P<0.05);但发酵24 h后,各试验组与对照组相比,差异均不显著(P>0.05)。
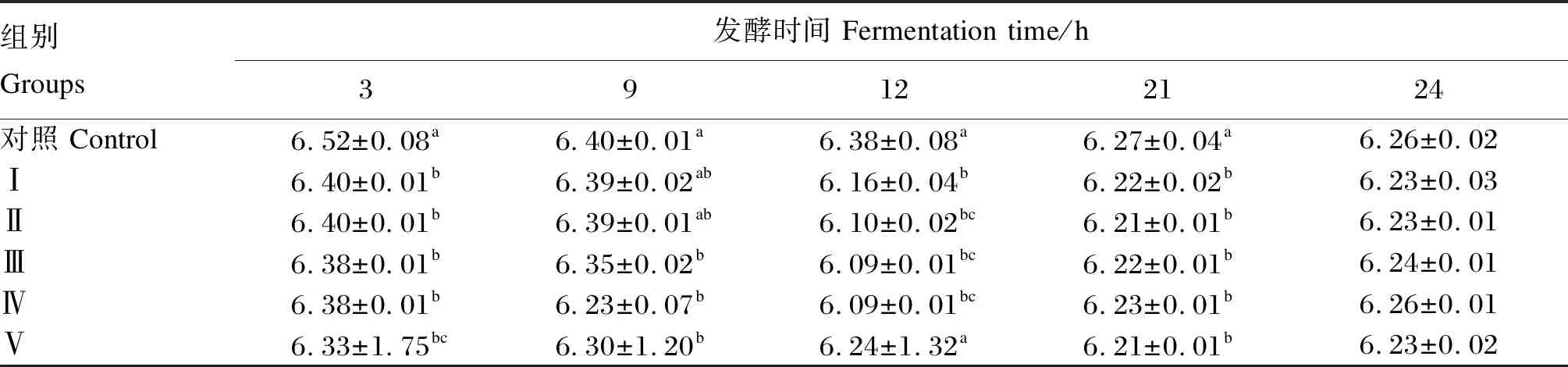
表3 不同浓度甘草酸单铵盐对发酵液pH的影响
2.3 不同浓度甘草酸单铵盐对发酵底物干物质降解率的影响
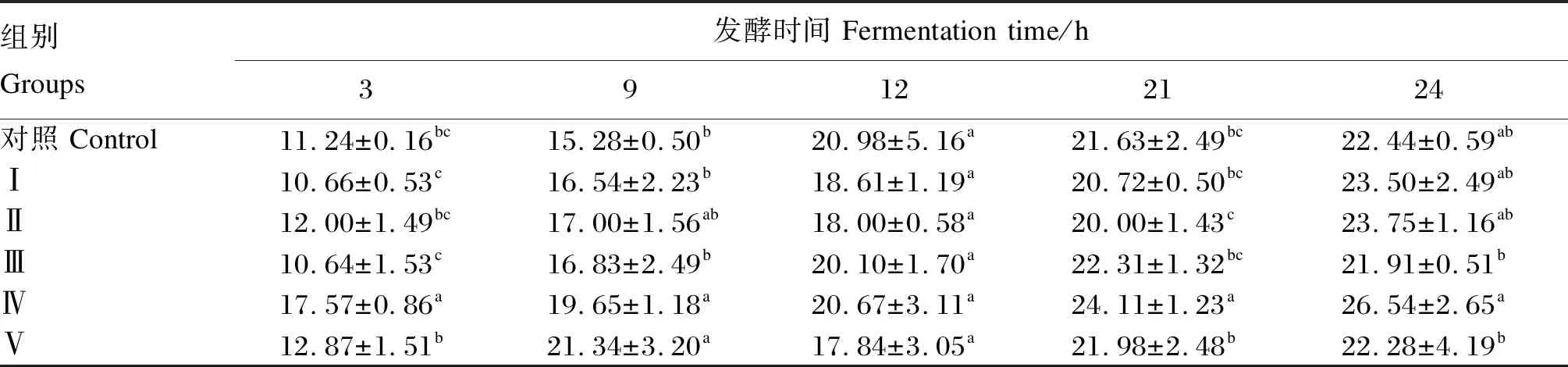
表4 不同浓度甘草酸单铵盐对发酵底物干物质降解率的影响
2.4 不同浓度甘草酸单铵盐对发酵液NH3-N浓度的影响
由表5可知,与对照组相比,各试验组的NH3-N浓度有下降的趋势。在发酵3 h后,Ⅰ组、Ⅳ组、Ⅴ组的NH3-N浓度均显著低于对照组(P<0.05);在发酵9 h后,Ⅲ组、Ⅳ组的NH3-N浓度显著低于对照组(P<0.05);在发酵12 h后,Ⅲ组、Ⅳ组、Ⅴ组的NH3-N浓度均显著低于对照组(P<0.05);在发酵21 h后,Ⅳ组的NH3-N浓度显著低于对照组(P<0.05);在发酵24 h后,Ⅱ组和Ⅴ组的NH3-N浓度显著低于对照组(P<0.05)。
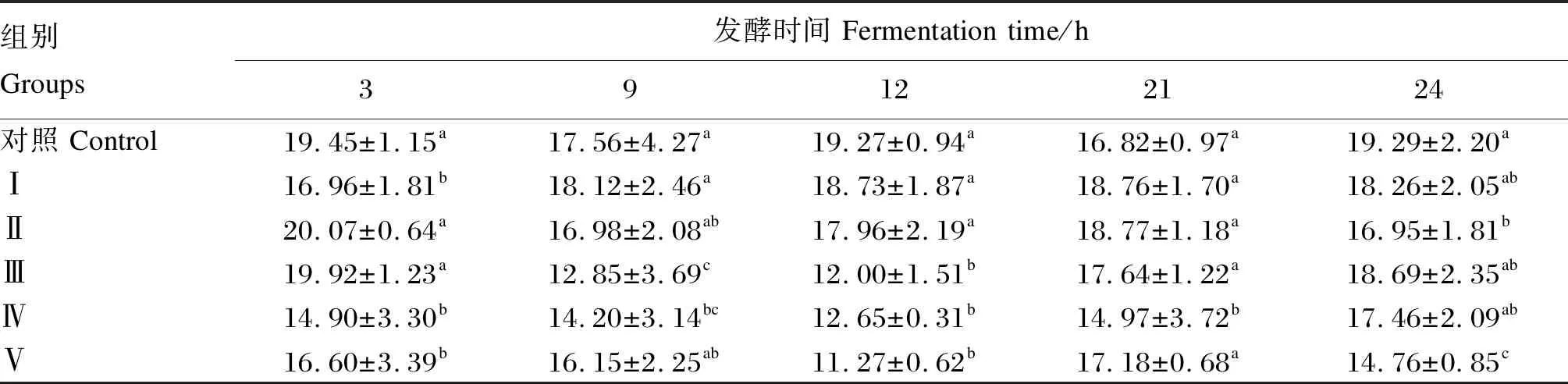
表5 不同浓度甘草酸单铵盐对瘤胃液NH3-N浓度的影响
2.5 不同浓度甘草酸单铵盐对发酵液VFA浓度的影响
由表6可知,随着发酵时间的增长,添加甘草酸单铵盐发酵液的VFA浓度均呈逐渐增加后稍有降低的趋势。与对照组相比,试验组乙酸浓度均呈降低的趋势,在发酵12 h后,Ⅲ组乙酸浓度显著低于对照组(P<0.05);在发酵3、9和12 h后,Ⅳ组乙酸浓度显著低于对照组(P<0.05);在发酵3和12 h后,Ⅴ组乙酸浓度显著低于对照组(P<0.05)。Ⅰ组再发酵12 h后,Ⅲ组在发酵12 h后,Ⅳ组在发酵3、9和12 h后,Ⅴ组在发酵3和12 h后丙酸浓度均显著低于对照组(P<0.05),但在发酵24 h后,各试验组丙酸浓度与对照组相比,差异不显著(P>0.05)。Ⅳ组在发酵3和9 h后,及Ⅴ组在发酵3、12及21 h后,丁酸浓度均显著低于对照组(P<0.05),在发酵24 h后,Ⅳ组的丁酸浓度最低,但与对照组相比差异不显著(P>0.05)。乙酸/丙酸随着甘草酸单铵盐浓度的增加逐渐降低。
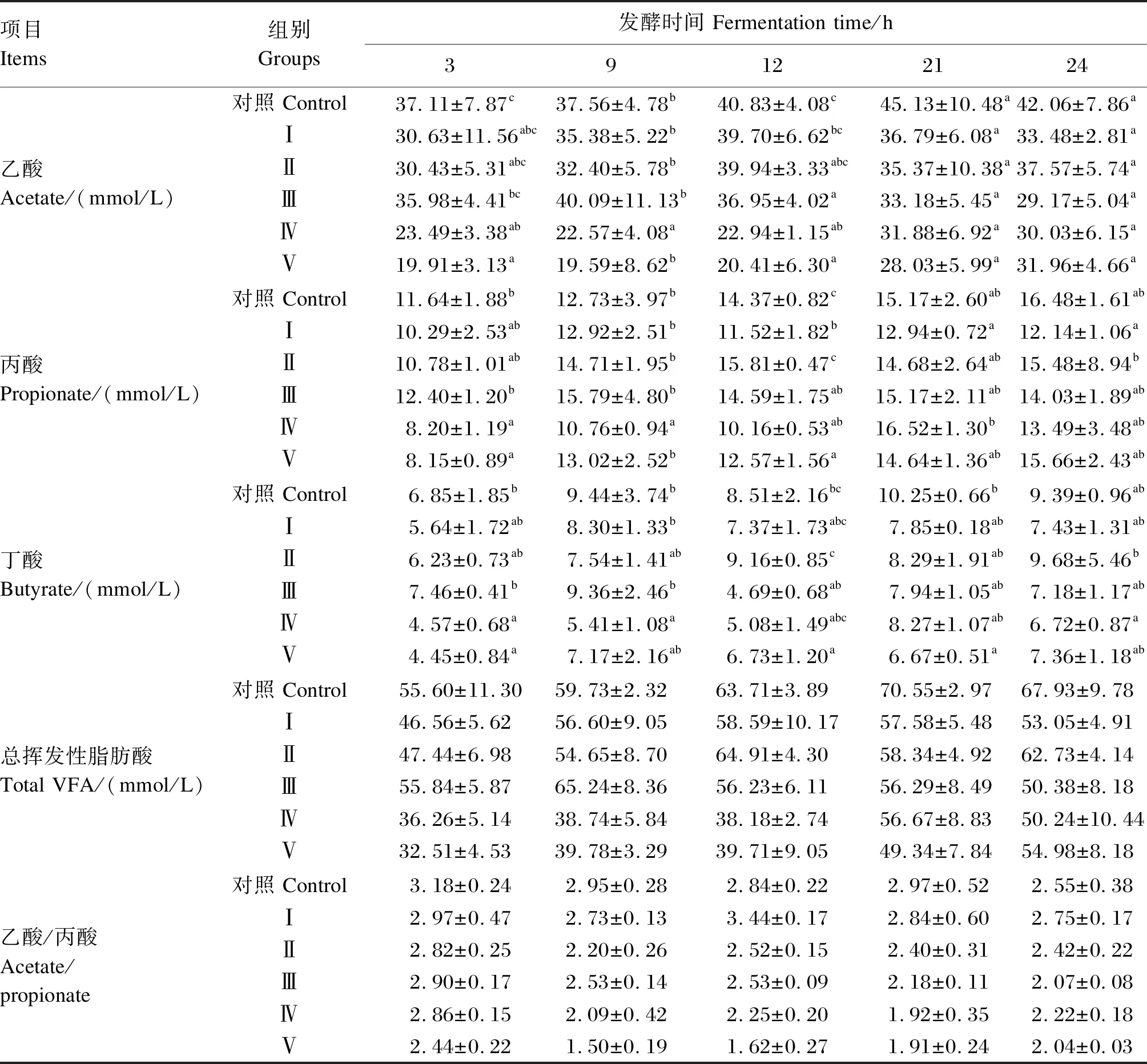
表6 不同浓度甘草酸单铵盐对瘤胃液VFA浓度的影响
2.6 不同浓度甘草酸单铵盐对发酵液甲烷产量的影响
由表7可知,随着各组发酵时间的增加,甲烷产量也随之增加,但甲烷产量随着甘草酸单铵盐浓度的增加而逐渐降低。在发酵9 h后,Ⅳ组的甲烷产量显著低于对照组(P<0.05),其他试验组均与对照组相比差异不显著(P>0.05);在发酵3、12、21、24 h后,各试验组与对照组相比差异均不显著(P>0.05)。
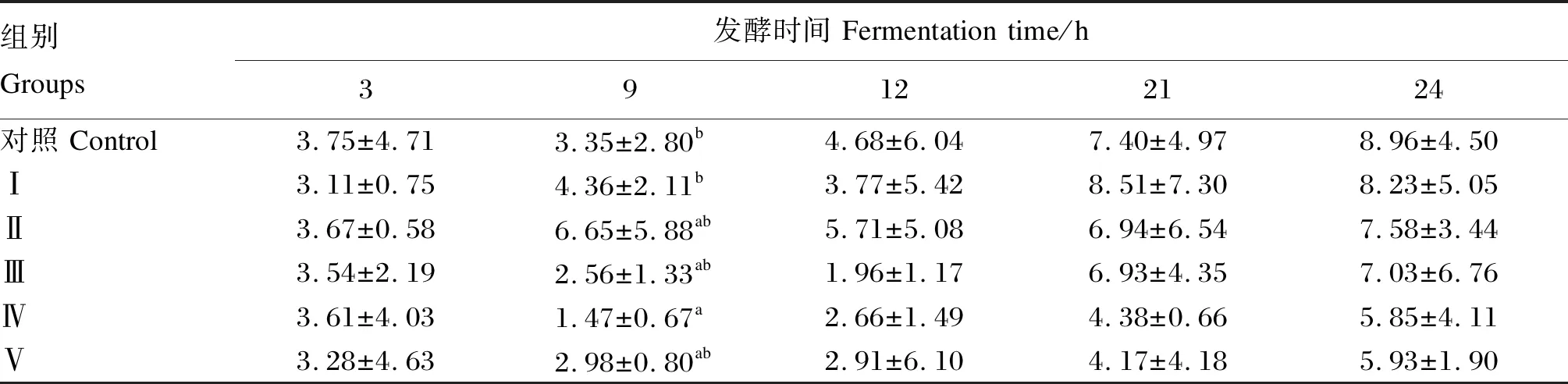
表7 不同浓度甘草酸单铵盐对发酵液甲烷产量的影响
3 讨 论
瘤胃产气量在一定程度上可以反映出发酵底物被瘤胃微生物利用的程度[15],同时产气量也能够表现出饲粮中可以利用营养成分的多少,当可利用的营养成分越高时产气量越高[16]。瘤胃气体主要来源于瘤胃中碳水化合物发酵所产生的气体,主要成分是二氧化碳(50%~65%)、甲烷(30%~40%),其中还有少量的氮气、氧气、氢气和硫化氢[17]。Hu等[18]利用不同浓度的茶皂素体外发酵24 h,结果表明添加茶皂素减少了瘤胃的产气量;郭雪峰等[5]利用甘草提取物进行体外发酵24 h,结果表明产气量随着甘草提取物浓度的增加而降低,与本试验结果一致。但王晓霞等[6]的研究发现,通过体外发酵试验研究无患子皂苷及茶皂素对瘤胃发酵及甲烷产量的影响,表明添加无患子皂苷及茶皂素均提高了瘤胃产气量并且降低了甲烷产量。牛俊丽等[7]的研究也表明丝兰皂苷提高了瘤胃产气量同时降低了甲烷产量。王新峰[8]在体外发酵试验中添加绞股蓝皂苷减少了产气量同时也减少了甲烷产量。不同植物皂苷均对瘤胃甲烷的产量起到抑制的作用,但产气量的高低有所不同,这可能是由于不同植物皂苷的分子结构不同,导致对改善瘤胃发酵途径有所不同,其原理有待进一步研究。
pH是反映瘤胃稳定性的重要指标。饲粮结构是影响瘤胃pH的主要因素,pH过高或过低对瘤胃微生物的生长和繁殖都会产生影响[19]。瘤胃发酵的pH正常范围一般在6.0~7.0。同时pH的变化会影响VFA浓度以及VFA组成[20]。本试验参考史高峰等[21]的方法测定甘草酸单铵盐呈弱酸性,可能影响到本试验各组的pH,使其有降低的趋势,但各组pH(6.09~6.40)始终处于6.0~7.0的正常范围之内,对瘤胃微生物的正常活动没有产生负面影响。
饲粮底物降解率在一定程度上反映瘤胃微生物对其利用的情况,同时降解率的高低与饲粮营养组成有着密不可分的关系[22]。潘龙等[23]的研究表明,添加柴胡皂苷提高了饲粮干物质降解率,这与本试验添加350 mg/L的甘草酸单铵盐提高了干物质降解率结果一致。也有研究表明,添加低浓度的丝兰皂苷可刺激某些瘤胃细菌的生长,调节微生物群落结构,从而提高了饲粮消化率,但高浓度的皂苷在改善瘤胃发酵上起到驱除原虫类似的特性,会导致饲粮消化率下降[24]。但徐晨晨等[25]的研究表明,添加苜蓿皂苷对饲粮干物质降解率无显著影响。因此,关于皂苷对饲粮降解率的影响结果报道不一致,这可能与皂苷的种类和添加剂量有关,具体作用机制有待进一步研究。
NH3-N是饲粮中蛋白质在瘤胃内发酵分解的产物,也是瘤胃微生物合成菌体蛋白的主要成分,其浓度反映瘤胃微生物对饲粮中蛋白质分解能力及吸收速率的平衡状态[26]。在瘤胃中的大部分细菌都是以NH3-N为生长的唯一氮源,而瘤胃原虫不能利用NH3-N合成所需的蛋白质,但可以产生大量NH3-N[27]。严淑红[28]报道奶牛灌服茶皂素可降低NH3-N浓度,其原因可能是茶皂素抑制了原虫的活性,本试验结果与其相似,随着甘草酸单铵盐浓度的增加,发酵液的NH3-N的浓度逐渐降低。但有其他研究表明添加绞股蓝皂苷使NH3-N浓度增加,其原因可能是绞股蓝皂苷导致原虫裂解,产生大量微生物蛋白并被其他微生物所利用,从而导致NH3-N浓度的增加[8]。本试验各试验组的NH3-N浓度均在6~30 mg/dL的正常范围之内,因此添加甘草酸单铵盐未对发酵液NH3-N的平衡性产生不利影响。
VFA可为反刍动物提供能量,是动物生长、繁殖与泌乳等基本活动的主要能量来源之一,其浓度变化直接影响反刍动物对营养物质的吸收、利用和生产能力等[29]。其中瘤胃微生物是影响VFA比例和利用率的主要因素。纤维降解菌利用饲粮中碳水化合物生成乙酸、甲酸、H2、乙醇和乳酸,同时瘤胃甲烷菌与其存在共生关系可以利用种间氢转移跳过降解过程中生成丙酸和琥珀酸等步骤,提高能量转化效率以及饲料利用率[30]。有研究表明,甲烷产量与纤维的降解率呈正相关,且反刍动物瘤胃中产甲烷菌的数量和纤维降解菌的数量也呈正相关关系[31]。本试验在添加甘草酸单铵盐后乙酸在发酵后期浓度有所降低,致使乙酸/丙酸下降,导致甲烷产量降低,与上述报道一致。张婷婷[32]研究报道,茶皂甙主要通过改变瘤胃VFA组成结构来调控瘤胃发酵模式,且改善瘤胃发酵的主要原因是降低了乙酸/丙酸,这与本试验的结果一致。但王勇等[33]报道称,在饲粮中添加苜蓿皂苷提取物提高了乙酸和丁酸的浓度,降低了丙酸浓度,改善了瘤胃发酵模式并提高了采食量及日增重。关于皂苷对瘤胃VFA组成影响的报道不一致,这可能与皂苷的种类及理化性质有关。也有研究报道认为瘤胃pH对VFA的结构也有影响,当pH在5.5时添加丝兰提取物改变了VFA结构,提高了丙酸浓度,降低了乙酸浓度,但pH在7.0时则对VFA结构无影响[34]。本试验添加甘草酸单铵盐发酵液的pH为6.09~6.40,可能会引起VFA结构的改变,但机理需要进一步研究。
影响甲烷产量的因素有很多,如饲粮营养结构、pH、乙酸/丙酸、瘤胃甲烷菌数量等[35]。反刍动物甲烷排放量其中90%是由瘤胃微生物甲烷菌产生[36]。本试验结果表明,随着甘草酸单铵盐浓度的提高,甲烷产量逐渐降低,说明甘草酸单铵盐对瘤胃甲烷的产生存在抑制作用,但同时瘤胃pH、NH3-N、VFA浓度等指标均在正常的范围内,说明甘草酸单铵盐在抑制甲烷产生的同时并未对瘤胃发酵产生负面影响。Rodríguez等[37]以狼尾草作为发酵底物,添加象耳豆皂苷体外发酵24 h并测定甲烷产量,结果表明甲烷产量显著降低。有研究认为丝兰皂苷、绞股蓝皂苷、丝兰皂苷等均可降低瘤胃甲烷的产生量[5-8],但降低瘤胃甲烷产生量的途径并不相同。有研究表明,皂苷可导致瘤胃原虫的数量减少,从而导致生成甲烷需要的H2减少,与原虫共生的甲烷菌数量降低[38]。但由于不同种类的皂苷在改善瘤胃发酵途径上有所不同,影响甲烷产量的机理可能也有所不同,因此甘草酸单铵盐抑制甲烷产生的微生物机理有待进一步通过体内试验进行研究。
4 结 论
① 甘草酸单铵盐可降低瘤胃液体外发酵的产气量、pH、NH3-N浓度、乙酸/丙酸、甲烷产量,但均未超过正常的生理范围。
② 在本试验条件下,通过各项指标分析比较,综合认为添加浓度为350 mg/L的甘草酸单铵盐适宜绵羊体外发酵。
猜你喜欢
西南石油大学学报(自然科学版)(2019年5期)2019-12-20 07:00:56
意林·全彩Color(2019年8期)2019-11-13 09:23:32
录井工程(2017年3期)2018-01-22 08:39:56
领导文萃(2017年10期)2017-06-05 22:27:01
中国卫生标准管理(2015年4期)2016-01-14 05:16:45
西南医科大学学报(2016年4期)2016-01-03 01:26:29
广州大学学报(自然科学版)(2015年4期)2015-12-23 11:50:10
中国当代医药(2015年33期)2015-03-01 02:09:17
丝绸(2015年11期)2015-02-28 14:56:49
中国药业(2014年20期)2014-05-17 03:13:44
