
林分密度对杉木人工林林下植被和土壤性质的影响
2021-07-09 05:07舒韦维卢立华何日明
生态学报 2021年11期
舒韦维,卢立华,*,李 华,农 友,何日明,陈 海,黄 彪
1 中国林业科学研究院热带林业实验中心, 凭祥 532600 2 广西友谊关森林生态系统国家定位观测研究站, 凭祥 532600 3 广西壮族自治区中医药研究院, 南宁 530022 4 国际竹藤中心园林花卉与景观研究所, 北京 100102
林下植被是森林生态系统的重要组成部分,在维持生态系统功能稳定性、增加生物多样性、改善立地条件,促进养分循环与能量流动发挥着不可取代的作用[1-3]。植被和土壤作为一个密不可分的整体[4],其相互协同变化的作用机理,维持着生态系统的动态稳定。而林分密度影响着植物群落对环境资源的利用与分配[5],是人工林群落结构的数量指标[6];同时,控制林分密度也是人工林经营的手段之一[7]。国内外学者对林分密度与林下植被和土壤理化性质的关系开展了大量的研究,主要认为合理调整林分密度能显著影响林分结构的空间异质性,改善林下微环境,进而使得林下植被物种多样性的改变[8- 10];另外,林下植被的发育不仅能有效提高土壤有机质,改善土壤理化性质,还能增加生物多样性[11]。
杉木(Cunninghamialanceolata)作为我国南方重要的速生用材树种,由于长期采取单一种植的经营模式,导致杉木人工林出现水土流失严重、病虫害加剧、土壤肥力衰退、生产力下降、生物多样性丧失等问题,严重制约着森林可持续经营[12- 14]。研究表明,初植密度过低(1667株/ hm2)或过高(10000株/ hm2)皆不利于杉木林下土壤理化性质和植物多样性的发展[15];随着间伐强度的增大,可增加杉木林下植被物种丰富度、多样性指数、盖度和生物量[9],显著改善土壤理化性质[16]。也有研究发现,间伐能明显改变灌木层和草本层物种组成,但对物种多样性的影响并不明显[17]。以往关于杉木的研究更多集中在人为干扰、林龄、经营方式等对物种多样性或土壤理化性质的影响,有关不同密度对物种多样性、土壤理化性质和生物量的综合研究较少。因此,本研究以南亚热带地区杉木人工林为研究对象,探究密度尺度下杉木人工林林下植被多样性、土壤理化性质和生物量之间的联系,旨在为该区域杉木人工林物种多样性保护、林地肥力维持及可持续经营提供科学依据。
1 研究区域概况
研究地设置在中国林业科学研究院热林中心青山实验场(106°39′—106°59′E,21°57′—22°19′N),地处广西凭祥市,地形起伏较大,最高海拔为1045.9 m,最低海拔为130.0 m,河谷海拔在130.0—150.0 m之间,属于典型的低山丘陵地貌。该地区年平均气温为20.5—21.7℃,7—8月极端高温40.3℃,1月份极端低温-1.5℃,雨量充沛,降雨时空不均匀,相对湿度大,年降雨量1200—1500 mm,集中在4—9月,为南亚热带半湿润-湿润气候。土壤主要由花岗岩发育而成的砖红土,其土层厚度大于80 cm[18],自然条件和地理位置决定了该地区地带性植被为南亚热带季雨林。马尾松(Pinusmassoniana)和杉木(Cunninghamialanceolata)为南亚热带主要优势树种,其次是格木(Erythrophleumfordii)、米老排(Mytilarialaosensis)、西南桦(Betulaalnoides)、红椎(Castanopsishpstrix)等。林下灌木主要为盐肤木(Rhuschinensis)、余甘子(Phyllanthusemblica)、桃金娘(Rhodomyrtustomentosa)、野牡丹(Melastomacandidum)等,草本主要有乌毛蕨(Blechnumorientale)、五节芒(Miscanthusfloridulus)、蔓生莠竹(Microstegiumvagans)等。不同密度样地林分概况如表1所示。本文林分和土壤数据均为2018年调查。

表1 样地基本概况
2 研究方法
2.1 样地设置及调查方法
该研究林地为1992年种植的杉木纯林,初植密度为2500株/hm2,经造林初期轻度间伐后再无人为干扰,基本处于自然恢复状态。根据研究需要,2005年选取地形因子、土壤类型等立地条件基本相似,林相整齐的林分作为样地林,对该杉木纯林进行4种不同密度处理,密度A:保留杉木株数35—40株 (密度为650株/hm2);B:保留杉木株数60—65株 (密度为1100株/hm2);C:保留杉木株数70—75株 (密度为1250株/hm2);CK:保留杉木株数95—100株 (密度为1650株/hm2),每种密度处理设置3个20 m×30 m的标准地,共12个样地。在每个样地内,用相邻网格法设置6个10 m×10 m的小样方,在样地四个角及中心设置5个5 m×5 m的灌木样方,5个1 m×1 m 的草本样方,分别对每个样方内物种基本概况进行调查,收集草本样方中凋落物的量,并做好记录。调查过程中将藤本植物及乔木幼苗记录到灌木层中。
2.2 样品采集与测定方法
土壤样品采集:分别在4种密度处理的试验地内,分别按上、中、下布设3个土壤剖面作为采样点,按0—20 cm,20—40 cm两层采集土壤样品1 kg左右,做好标记,带回实验室,去除植物根系、杂物和石砾,至于阴凉处自然风干,用于测定土壤理化性质。
土壤理化测定:采用干燥法测定土壤含水量,电位法测定土壤pH;土壤容重与总孔隙度等物理性质采用环刀法取样测定;有机质含量采用重铬酸钾水合加热法;全氮采用硒粉-硫酸钾-硫酸消化蒸馏滴定法,水解氮采用碱解扩散法;全磷采用硫酸-高氯酸消煮-钼锑抗比色法,有效磷采用盐酸-硫酸浸提法;全钾采用氢氧化钠碱熔,火焰光度计测定,速效钾采用乙酸铵浸提,火焰光度计法测定。
生物量测定:通过“全收获法”将灌木和草本分为地上部分和地下部分,以测其鲜重,并提取样品进行养分含量分析。凋落物层也进行全收获法测定其鲜重并分别采取分析样品,将样品带回实验室,置于80℃下烘干至恒重后称干重,并换算成单位面积干物质的量。
2.3 数据处理
根据调查内容,统计样方内物种组成,并对表征林下植被物种多样性基本特征的Shannon-Wiener指数H′、Simpson指数D、Pielou均匀度指数Jsw、Patrick丰富度指数S′进行计算,各指数计算公式如下:
Patrick 丰富度指数:
S′=S
式中:S为物种数,Pi为第i种的个体数ni占所有种个体总数n的比例,即Pi=ni/n(i=1,2,3,……,S)。
所有数据分析及作图利用R 3.6.1完成。采用单因素方差分析(one-way ANOVA)和最小显著差异法(LSD)检验不同密度处理下生物多样性、生物量、土壤物理性质的差异显著性,对杉木林下植被物种多样性指数与土壤理化性质、生物量做Pearson相关分析。
3 结果分析
3.1 不同密度下物种多样性指数
图1显示,杉木林下植被各多样性指数在不同密度间均无显著差异;灌木层中,随林分密度升高,Simpson指数、Pielou均匀度指数表现出逐渐递减的趋势,变化范围分别为0.798—0.760,0.855—0.828;而Shannon-Wiener指数、Patrick丰富度指数随林分密度的增加均呈现出双峰型变化趋势,峰值出现在密度A、C处,且4种多样性指数最小值均出现在密度CK处。草本层中Pielou均匀度指数随林分密度的增加,表现出先增加后减少的单峰趋势,最大值出现在密度C处;其余3种多样性指数变化趋势与灌木层Shannon-Wiener指数、Patrick丰富度指数变化趋势一致。
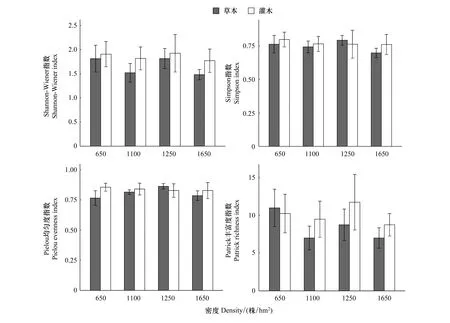
图1 不同密度杉木人工林林下物种多样性指数Fig.1 Species diversity index of understory vegetation in Cunninghamia lanceolata plantation with different densities
3.2 不同密度下植被生物量
如图2所示,不同密度杉木林下灌木层总生物量(地上+地下)随林分密度的增大呈先减后增的“U”字型变化趋势,在密度A最大,C最小;草本层总生物量(地上+地下)随密度的增加而减少。凋落物的量与林分密度呈正相关,随密度的增大而增大。林下生物量在各组分的分配表现为:凋落物>草本地上>灌木地上>草本地下>灌木地下,且地上部分占比较地下部分大。不同密度间,草本地上、灌木地下生物量在数值上波动较大,灌木地上、草本地下生物量则波动较弱,但差异均未达到显著水平。由此可知,草本地上生物量对不同密度的响应较灌木更为敏感。
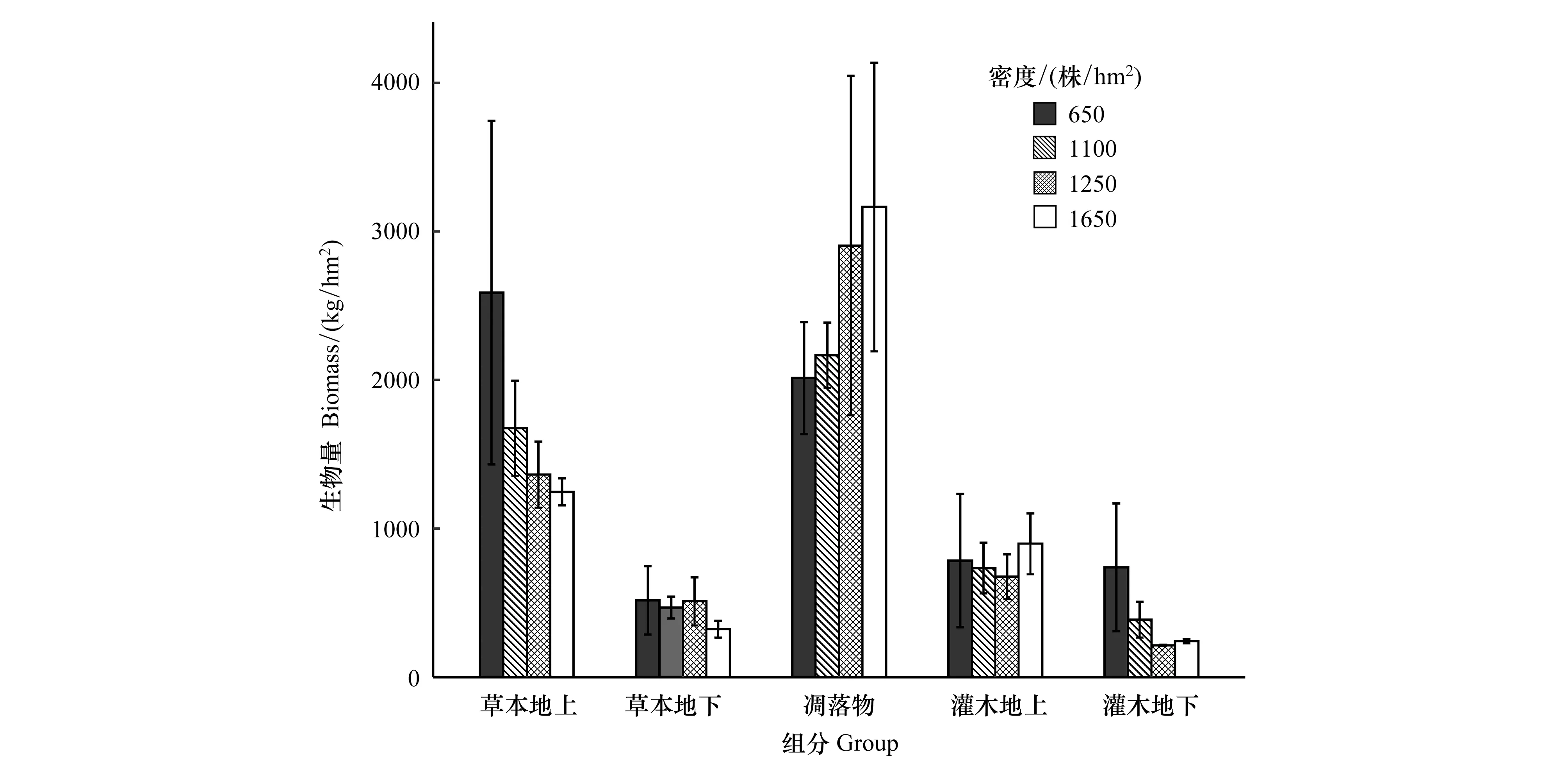
图2 不同密度杉木人工林林下植被生物量Fig.2 Biomass of Cunninghamia lanceolata plantations with different densities
3.3 不同密度下土壤理化性质
从表2可以得出,林分密度对土壤理化性质无显著影响(P>0.05),土壤养分含量和含水量随土层深度的增加而降低,土壤容重、pH随土层深度增加而增加; 土壤pH 值范围为4.23—4.44之间,为强酸性,是强烈淋溶作用所致,与杉木生长特性相符。0—20 cm土层,全磷、有效磷的含量随密度的增加呈现出先减少后增大再减少的双峰趋势,在密度A和C处出现峰值,全钾、速效钾的含量则在密度C处为最大值的单峰趋势;20—40 cm土层,全磷、有效磷、全钾、速效钾的含量均随密度的增大表现出先增大后减少的趋势;有机质含量随林分密度增加而降低。土壤养分与杉木林下灌、草层多样性指数变化趋势基本一致,说明林下植被与土壤密切相关。
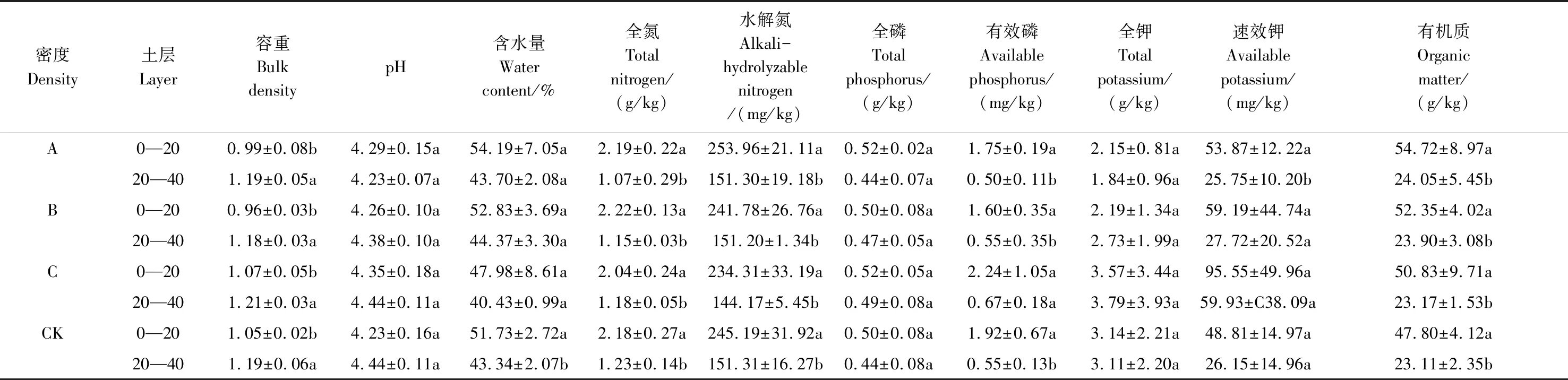
表2 不同密度杉木人工林土壤理化性质
3.4 不同密度下物种多样性指数与土壤理化性质的关系
多样性指数与土壤理化性质的相关分析(表3)表明:灌木层中,Shannon-Wiener指数与水解氮、有机质呈显著正相关(P<0.05),Patrick丰富度指数与全氮、水解氮、有效磷呈显著正相关;草本层中,4种多样性指数与土壤理化指标均不存在显著相关关系。

表3 不同密度杉木人工林林下物种多样性指数与土壤理化性质的相关系数
3.5 不同密度下物种多样性指数与生物量的关系
表4结果显示灌木层Shannon-Wiener指数与灌木地上生物量呈极显著正相关(P<0.01),与灌木总生物量呈显著正相关(P<0.05);Simpson指数与灌木地上生物量、总生物量呈显著正相关(P<0.05);Patrick丰富度指数与灌木地上、总生物量呈极显著正相关(P<0.01),与草本地上、总生物量呈显著正相关(P<0.05),其余指数和生物量无相关性。草本层Shannon-Wiener指数、Patrick丰富度指数与灌木地下生物量呈极显著正相关(P<0.01);Simpson指数灌木地下生物量呈显著正相关(P<0.05)。

表4 不同密度杉木人工林林下物种多样性指数与生物量的相关系数
4 结论与讨论
4.1 林分密度对杉木林下植被物种多样性的影响
物种多样性能清楚的反映出群落物种组成、林分结构、动态演替等方面的重要指标,在维持生态系统的稳定中发挥着举足轻重的作用。调查发现,因实验林地长期无人为干扰,杉木林乔木层已有一定数量的常绿阔叶树种,如米老排、大叶栎、红椎、粗叶榕、杜茎山等,但杉木依旧为其群落的优势种,在群落中处于绝对优势地位。林分密度变化导致杉木种群数量发生变化, 使其对林下物种的抑制效应与林下物种所占有的空间、水、热等资源比率也随之发生变化[19]。本研究中,杉木林下灌木层和草本层物种多样性对密度差异的响应机制不同,灌木层中Simpson指数、Pielou均匀度指数随林分密度升高而降低,这与以往研究结果一致,间伐能够促进林下植被的物种丰富度和多样性,且随间伐强度的增大而增大[20-21]。但Shannon-Wiener 指数、Patrick丰富度指数随林分密度的增加均呈现出双峰型变化趋势,在密度A和C处均出现峰值;在密度C处出现峰值可能与杉木立木结构有关,杉木杆材修值挺拔,顶端优势明显,自然整枝强烈,叶片细小空间排列较为稀疏,层间透光率高,高密度种植增加了林下生境的复杂性,有利于林下植被生长,使灌木长势较好种类增多[15]。随着林分密度的增大,杉木林下草本层Shannon-Wiener指数、Simpson指数、Patrick丰富度指数均随着密度的增大表现出先减少后增大再减少的趋势,Pielou均匀度指数则随林分密度的增加,呈现出先增加后减少的单峰趋势,最大值出现在密度C处;原因可能是随着森林的生长,林分郁闭度增加,各密度处理的林分抑制林下灌木和草本生长发育的作用变弱,在密度C下,由于生境优势,相较于其他密度林分更有利于草本植物的拓植。
Alem 等[22]在研究桉树与柏树纯林及混交林相互作用中发现,乔木层密度与林下植被多样性之间无显著关系;尤业明等[17]对间伐8 a后的杉木林研究表明,不同间伐强度并没有明显改变林下灌木层的物种丰富度和多样性;此外,同区域采取间伐处理的马尾松林也并未显著提高林下物种多样性[23],与本研究结果一致。虽然林下植被多样性指数在数值上各有高低,但灌木层、草本层4种指数对于密度变化的响应均无显著差异(P>0.05)(图1),其原因可能是因为本研究地的林分已接近过熟林(27 a),且为间伐抚育后的第15 a,随林龄的增长,郁闭度增加,人为干扰少,林下环境越是接近,因而导致不同密度的林下植被物种多样性差异不显著。
4.2 林分密度对杉木林下植被生物量的影响
林下植被生物量是评价森林群落物质生产及能量平衡的基础,是植物资源利用、生态位占有、种内种间竞争能力的体现[24]。另外,生物量作为衡量林下植被的重要指标之一,在一定程度上能反映物种的丰富程度[25]。森林群落中乔木层的郁闭或者稀疏,都会直接影响林下植被的生长发育,从而引起灌木层、草本层生物量的差异[26]。本研究发现,灌、草层总生物量在密度A值最大,且草本层生物量总体占优,这是因为林分发育过程中,郁闭度增加,灌木层也逐渐从阳生树种向中性或阴生树种发展,导致灌木层生物量下降;另外,密度越低,草本层长势越好,草本层与灌木层之间的竞争使得灌木层生物量下降。随着林分密度的变化,草本地上、灌木地下生物量较其他组分波动幅度大,草本地下、灌木地上生物量较为稳定,原因可能是随着林分逐渐郁闭,林下植被通过光合作用积累物质失去了有力的光照条件,草本植物选择将更多的碳投入到地下,减少地上部分的分配,从而增加草本根系生物量,以便在不良环境中依靠根系吸收水分和养分资源,增强自身的竞争能力[27],而灌木应对不良环境的生存策略则与草本相反。相关研究认为,林分密度和郁闭度的改变会导致林内水热条件和生物量的变化,进而影响凋落物的积累量[26]。本研究中,凋落物的量随林分密度的增加而增加,也就是说,凋落物产量受密度大小的影响,与Roig等[28]研究结果一致。林分密度降低,凋落物分解加速,其积累量会逐渐减少,回归土壤的养分含量增加,使土壤肥力的时效性延长[29-30];此外,Kunhamu等[31]研究认为,间伐降低了凋落物中氮、磷、钾的含量,导致落叶量减少。
本研究发现4种多样性指数与灌木层生物量的相关性较为密切,却与草本、凋落物层相关性均不明显,可能是由于生物与非生物等各种因素综合作用的结果。林下植被多样性与生物量动态平衡体现了其相互作用的过程,一方面,物种多样性特征影响叶面积指数,从而影响林内小气候和种间关系, 改变光合速率,间接调控林下植被长势;另一方面,生物量影响林分地上地下能量分配,改变凋落物数量影响养分循环和土壤理化性质,反作用于生物多样性的发展。可见物种多样性与生物量关系密切,协调好两者关系有利于人工林可持续经营,因此林分密度对杉木人工林下植被多样性与生物量之间的影响机理还需深入探究。
4.3 林分密度对杉木林土壤理化性质的影响
林分密度的改变使得林下植被种类和组成发生改变,势必造成该地生境的变化,进而导致土壤理化性质的不同[32]。本研究中,0—20 cm土层,在A处理下全磷含量最高,B处理全氮含量最高A处次之,且均高于CK 样地,氮、磷作为植物生长最重要的限制因子,其含量的增加与植物多样性密不可分。首先,低密度下,乔木及灌草层植物根系有足够的分布空间,根系发达,根系间相互作用导致微生物活性增强、数量增多[33],加强了土壤对氮的循环[34];再者,林分密度会影响凋落物数量,并通过光照、温湿度等环境因素影响凋落物分解速率;其次,植物根呼吸及养分元素的吸收存在种间差异,促使土壤动物群落发生改变,进而增强了凋落物分解,大量的磷元素归还于土壤[35]。有研究表明,间伐后光照强度的增加会促使植物对有效磷和速效钾的需求[36],土壤内有效磷和速效钾含量会随间伐强度的增大而减少,但由于本研究林地处于成熟林阶段,具有稳定性,土壤养分也趋于稳定,虽然A处理下有效磷和速效钾含量相对较低,可各处理间差异并不显著。而在0—40 cm土层中全磷、有效磷、全钾、速效钾含量在密度C值最大,是因为土壤养分存在垂直分布差异,深层土受林分密度的影响较小。当物种多样性较高时,地被植物生物量、盖度高,凋落物不断积累,从而丰富了土壤的有机质[37];本研究中草本层生物量随林分密度增大而减少,与有机质含量变化趋势一致。灌木层Shannon-Wiener指数、Patrick丰富度指数与全氮、水解氮、有效磷、有机质之间呈显著正相关关系,表明全氮、水解氮、有效磷、有机质是影响该地区杉木林下植被生长分布的主要因子,因此,在成熟林中适当施加氮磷肥,减弱其淋溶作用,可促进灌木层物种多样性的提高。另外研究发现,草本层多样性指数与理化性质间不存在明显关系,这与Andrew等[38]研究结果一致,可能与杉木林立地条件以及观测时间有关。
4.4 结论
灌木层与草本层多样性指数均在密度A(650株/hm2)出现峰值,其地上生物量大于地下生物量,且总生物量都在密度A达最大,表明在此密度下光照、水分、养分等环境因子有利于林下植被的生长,杉木林下植被丰富、群落稳定性水平较高。对土壤理化性质分析可知,有机质含量随密度的增大而减少,全氮、水解氮、有效磷、有机质为影响该区域杉木林林下物种多样性的主要因子。本研究通过对不同林分密度杉木人工林林下植被、生物量、土壤理化性质研究发现,对于成熟林分,密度A(650株/hm2)可为植物生长提供良好的生长空间并改善林地土壤肥力,有利于促进林下植被的发育,增加林下物种多样性和生物量,因而可作为最适密度,为该地杉木林可持续经营和稳定发展提供参考。
猜你喜欢
乡村科技(2021年1期)2021-12-03
青年文学家(2021年16期)2021-08-05
少儿美术(2020年8期)2020-11-06
农民致富之友(2020年4期)2020-04-03
诗潮(2019年10期)2019-11-19
中国水土保持科学(2019年6期)2019-04-26
中央民族大学学报(自然科学版)(2018年4期)2018-03-19
农业与技术(2017年9期)2017-05-28
诗潮(2017年2期)2017-03-16
绿色科技(2014年4期)2015-01-06
