
青藏高原达则错近1000年来生态系统变化及可能机制*
2021-07-07 03:31李秀美侯居峙王明达
湖泊科学 2021年4期
李秀美,侯居峙,王明达,徐 磊
(1:信阳师范学院地理科学学院/河南省水土环境污染协同防治重点实验室/豫南岩矿宝玉石鉴定及加工中心,信阳 464000)(2:中国科学院青藏高原研究所,北京 100101)(3:辽宁师范大学地理科学学院,大连 116029)(4:中国水产科学研究院南海水产研究所,广州 510300)
全球变化背景下,对人类赖以生存的地球环境的过去、现在和未来的研究成为国际学术界广泛关注的热点问题.湖泊作为生态系统的重要组成部分,是地球表层各圈层(大气圈、水圈、岩石圈和生物圈)相互作用的重要连接枢纽,通过地表水的汇聚和蒸发成为地表水循环过程的重要环节,对气候变化极为敏感[1].湖泊水体的物理化学参数是影响湖泊生态系统的重要因素, 其在气候变化下的改变深刻地影响着湖泊生态系统的组成与变化趋势[1].研究表明,全球变化已导致地球上的湖泊生态系统状态发生变化,对其生态系统结构和功能、生物多样性构成严重威胁[2-3].
青藏高原平均海拔在4000 m以上,被称为亚洲水塔”、“地球第三极”,是我国最大的生态脆弱区,具有生态系统结构简单、抗干扰能力弱和易受全球环境变化影响的特点[4-5].在青藏高原上分布着全球海拔最高、面积最大、数量最多的高原湖泊群,其中面积大于1 km2的湖泊数量有1171个[6].青藏高原地区海拔高,气候寒冷,环境恶劣,绝大多数湖泊受人类活动的直接干扰较小, 湖泊特征变化忠实而敏感地记录了该区气候与环境的变化过程[1].第二次青藏高原综合科学考察研究将获取青藏高原高分辨率的湖泊生态系统及其影响因子数据, 认识和评判湖泊变化对气候变化和人类活动的响应过程、机制和程度作为重要研究内容[1].当前,青藏高原气候变暖的速度是全球平均速度的3倍,并通过改变湖泊水温和盐度来影响湖泊生态系统[3].但长时间尺度青藏高原湖泊生态系统变化及其对气候环境变化的响应还不甚清楚.
目前的研究手段如现场观测、围隔实验和模型模拟等可以很好地揭示湖泊生态系统在有观测记录以来的演替和变化过程,但是不能提供历史时期湖泊生态系统的变化及其对气候环境变化和人类活动的响应.而湖泊沉积物具有信息丰富、连续性好、沉积速率快和地理覆盖面广等特点,记录了流域人类活动的信息以及湖泊生态环境的演变过程,在过去全球变化和区域响应方面具有不可替代的优势[7],成为开展较长时间尺度生态系统及环境演变研究的有效载体.保存在沉积物中的生物、地球化学元素等指标可以指示湖泊内部的环境变化,以及水生生态系统的演变信息.例如长链烯酮(long-chain alkenones, LCA)不饱和度可以定量重建古温度[8-11];长链烯酮化合物的总浓度可以重建浮游植物藻类的生产力[12];总有机碳(TOC)是反映湖泊沉积物有机质含量的有效指标[13],可指示湖泊及其流域初级生产力总量的变化特征[14];湖泊沉积物中的总氮和总磷含量能够反映不同时期水体的营养状况;保存在湖泊沉积物中的溞类残体(壳瓣及休眠卵)能够再现溞属种类的历史演变过程[15-17].总之,古湖沼学可以为探讨湖泊生态系统的长期变化及其对气候环境变化的响应提供重要信息.
以西藏拟溞(Daphniatibetana)为优势浮游动物物种的咸(盐)水湖是西藏湖泊的一个重要类型,该类湖泊占到西藏湖泊数量的一半以上[18].相对于淡水环境,咸(盐)湖泊往往具有高离子浓度、寡营养等诸多特点.位于藏北腹地的达则错是该类型湖泊的典型代表,湖区人烟稀少,且水深大,其深水区沉积过程基本排除了人为干扰及风浪扰动影响,因此可代表低矿化度深水盐湖的自然沉积过程[18],是开展较长时间尺度湖泊生态系统演变及其对气候环境变化响应研究的理想区域.目前,国土资源部盐湖资源与环境重点实验室已经对该湖的水化学及生物动态进行了长期监测[18],但气候变化影响下的湖泊生态系统变化过程和机制目前还不清楚,关于该区历史时期湖泊生态系统变化及其对气候环境演变的信息还极为有限.为此,本文通过达则错2根沉积岩芯的总氮、总磷和TOC含量进行分析,重建历史时期达则错营养盐以及有机质变化情况,结合沉积物中西藏拟溞残体丰度和总烯酮含量记录的生物信息,以及前期由长链烯酮不饱和度重建的气候变化信息,揭示青藏高原中部达则错历史时期湖泊生态系统变化及其对气候环境演变的响应机制.本研究可以为青藏高原脆弱生态环境的保护和修复提供决策支持.
1 资料与方法
1.1 研究区概况
达则错(31.82°~31.98°N, 87.42°~87.65°E)(图1)地处藏北高原腹地,位于西藏自治区那曲地区尼玛县境内的一个断陷盆地中,海拔4450 m,属于碳酸盐型咸水湖[19].湖泊长21.1 km,最大宽16.9 km,最大水深38 m,湖泊面积和流域面积分别为245和10885 km2,补给系数44.5[19],湖水主要依赖波仓藏布补给,结冰期为11 月至次年4 月.湖区严寒干燥,距离达则错150 km的申扎气象站(30.57°N, 88.38°E,海拔4672 m)数据显示该地区1981-2012年期间年均降水量为316 mm,90%降水量在7-9月;年均气温0.55℃,年平均夏季温度(7-9月)8℃.

图1 达则错地理位置(A)、湖泊水系(B)和水深示意图(C)
达则错属于半对流湖泊,可能与湖泊深部湖水盐度较高有关[20].根据2012年8月实地观测(图2),达则错温跃层在水深16~23 m处,湖水pH为9.80~10.04,表层湖水盐度为14.69 g/L,随着深度增加盐度也随之增大,湖水底层盐度为21.41 g/L,在25~29 m处出现盐跃层;表层湖水溶解氧浓度为5.42 mg/L,23~24 m处溶解氧浓度最大为7.26 mg/L, 底层湖水溶解氧浓度为0.湖水透明度较小,塞氏盘深度为6 m.湖区东部存在很多古湖岸阶地,最高的阶地比现代湖面高约57 m,表明达则错对过去气候变化响应敏感[21].达则错为无鱼湖泊,经鉴定,达则错浮游植物以蓝藻、硅藻、裸藻、绿藻为主;优势浮游动物物种为西藏拟溞(Daphniatibetana)[22],占该湖浮游动物生物量的82.30%[18].
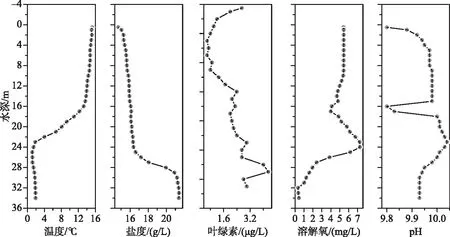
图2 达则错水质参数垂向变化(测于2012年8月)
1.2 样品采集与处理分析
用于研究的达则错柱状沉积物DZC2011(长291 cm)和DZC2012(长76 cm)分别采集于2011年夏季和2012年夏季.利用 Uwitec 采样平台进行达则错湖芯样品的钻取,采样点位于同一点位(31.89°N,87.55°E,4420 m),采样点水深37 m(图1C).利用手持测深仪选取水深较深且地形相对平缓地区进行取样.样品采集后用Uwitec分样器按0.5 cm为间隔进行分样,分完后装入Nasco©whirl pak 无菌采样袋密封冷冻保存.
沉积柱年代学框架基于放射性年代学方法(210Pb、137Cs和14C),包括11个210Pb、137Cs定年以及7个湖泊沉积物全样有机质14C定年.对获得的7个14C年代用INTCAL13树轮校正曲线进行日历年龄校正,并剔除碳库效应(2552 a),最后运用Bacon软件计算得到整个沉积柱的年龄为3100 a.详细定年方法及结果参照文献[23].本研究利用DZC2012沉积柱进行总氮(TN)、总磷(TP)、总有机碳(TOC)含量和西藏拟溞丰度等指标测试的分析,结合前期利用DZC2011沉积柱测试的烯酮有机指标[23],分析达则错历史时期湖泊生态系统变化及其对气候环境演变的响应机制.
达则错2011-1钻孔200 cm深度以下烯酮含量太低,接近检出限,因而我们只对上部200 cm(约近2000年)烯酮含量变化进行讨论.烯酮化合物的前处理及测试分析步骤详见文献[23].沉积物总氮和总磷含量在暨南大学水生生物研究中心测定,沉积物样品冷冻干燥后,去除杂质,经玛瑙研钵研磨后过100目尼龙筛保存备用,总氮含量采用碱性过硫酸钾消解法测定[24], 总磷含量测定采用SMT法[25]测定;总有机碳分析测试在中国科学院青藏高原研究所环境变化与地表过程实验室进行,样品冷冻干燥后,称取2份样品(100.0 mg),分别放置在两小舟(材料为耐高温陶瓷,TOC测试仪专用)内,用日本岛津公司生产的配有SSM-5000A固体样品进样附件的TOC-Vcph型总有机碳分析仪(重复测量的相对标准偏差(RSD)<1%)对样品进行分析测试,得到总碳(TC)和无机碳(IC)含量,通过差减法计算得到样品的总有机碳含量;在显微镜下对沉积物中西藏拟溞残体(壳瓣及休眠卵)进行鉴定、计数和拍照,详细的步骤参考文献[26],达则错湖泊沉积物中西藏拟溞残体的鉴定在暨南大学水生生物研究中心进行.
1.3 统计分析
去趋势对应分析(DCA)结果显示前4个排序轴的梯度长度最大值低于3,因此本文选择线性模型冗余分析(RDA)用于测试环境因子与生物因子之间的关系,采用蒙特卡罗置换检验每个有效环境变量的显著性.RDA分析中,以西藏拟溞丰度和长链烯酮化合物总含量反映的湖泊浮游植物生产力作为响应变量,以环境指标(TN、TP、TOC和温度数据)作为解释变量.所有排序分析均在CANOCO 5软件中运行[27].
2 结果
2.1 达则错沉积物中西藏拟溞残体的电镜图片特征
经鉴定,达则错沉积物中浮游动物残体主要为西藏拟溞壳瓣和休眠卵.西藏拟溞壳瓣和休眠卵电镜图片见图3.图3a、b为西藏拟溞壳瓣侧面观,壳瓣背侧无脊棱背缘微凸,腹缘凸出较甚,后背角钝,无壳刺;图3c、d为西藏溞休眠卵卵鞍背面观,卵鞍狭长,豆荚状,但前缘稍弯曲,背缘边布满栉刺,内储冬卵2枚,冬卵的长轴与卵鞍背侧相互平行.

图3 达则错沉积物西藏拟溞壳瓣(a、b)和休眠卵(c、d)显微照片
2.2 达则错温度记录
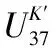

图4 过去1000年达则错西藏拟溞丰度(a)、总烯酮含量(b)、总磷含量(c)、总氮含量(d)、TOC含量(e)、温度(f)记录以及达索普冰芯浓度(g)变化(紫色阴影指示湖区低温时段,红色阴影指示湖区受人类活动显著影响阶段,红色虚线指示达则错生态系统开始发生显著变化的时间)
2.3 达则错沉积岩芯中各指标变化特征
过去1000年,达则错湖泊沉积物中西藏拟溞残体丰度、TN、TP及TOC含量总体上呈现较为一致的变化趋势(图4).西藏拟溞丰度变化范围为1.1×104~9.2×105ind./m2,长链烯酮含量变化范围为22.5 ~1.26×104mg/g,TN含量的变化范围为0.70~2.50 mg/g,TP含量的变化范围为0.14~0.65 mg/g,TOC含量的变化范围为0.70%~6.29%.根据各指标在过去1000年的变化特征,将其分为6个阶段:
第一阶段(950-1050 AD),西藏拟溞丰度、TN、TP及TOC含量较低,长链烯酮含量较高;第二阶段(1050-1250 AD),西藏拟溞丰度、TN及TOC含量明显上升,TP含量略有升高,长链烯酮含量显著降低并达到全岩芯最低值;第三阶段(1250-1450 AD),西藏拟溞丰度、TN及TOC含量明显降低,TP含量变化微弱,长链烯酮含量在1250 AD开始显著上升并维持在高值阶段;第四阶段(1450-1600 AD),西藏拟溞丰度、TN及TOC含量缓慢上升,TP含量变化微弱,长链烯酮含量明显降低;第五阶段(1600-1850 AD),西藏拟溞丰度、TN及TOC含量逐渐降低并维持在相对低值阶段,TP含量缓慢降低,长链烯酮含量逐渐升高;第六阶段(1850-2000 AD),西藏拟溞丰度和TOC含量逐渐升高,长链烯酮含量呈波动上升趋势,TN和TP含量快速升高并达到过去1000年最高值.
3 讨论
3.1 达则错沉积物中各生物化学指标的指示意义
西藏拟溞是达则错的优势浮游动物物种,广泛分布在海拔4000 m以上的青藏高原咸水湖泊中[26],适合在高寒、贫营养性盐水水体中生活,为一冷水性广盐种,在冰下水温为-1~-2℃范围生长繁殖[29].在不良环境条件下,水体中的现生西藏拟溞会产生休眠卵保存在沉积物中,在条件适宜时这些休眠卵通过孵化补充到现生种群中.研究表明,沉积物中保存的西藏拟溞残体(壳瓣残体及休眠卵)可以揭示过去长时间尺度西藏拟溞丰度的变动,进而反映气候环境变化(温度、湖泊水位及营养盐浓度等)[26].因此,达则错沉积物中西藏拟溞壳瓣及休眠卵含量能够指示历史时期湖泊中浮游动物优势种西藏拟溞丰度的动态变化.过去1000年达则错西藏拟溞丰度呈波浪形变化,在1050-1250、1450-1600、1850-2000 AD含量较高,在950-1050、1250-1450和1600-1850 AD含量较低,最大值出现在约1165 AD.
长链烯酮是指由藻类合成的碳数范围为C35~C42且具有2~4个不饱和键的酯类化合物.目前湖泊中长链烯酮的具体母源藻种类还不清楚.培养实验表明单鞭金藻(Chromulinasp.)、等鞭金藻(C.lamellosa)、颗石藻(E.huxleyi)可能是湖泊长链烯酮的母源[30].近年来,随着分子生物学的研究发现湖泊环境中烯酮母源藻可能还存在其他物种,具有多样性[31].研究发现湖泊沉积物中长链烯酮化合物的总浓度可以反映湖泊浮游植物的生产力[12].在湖泊深水区域,水草和底生藻类数量甚微,浮游植物几乎是湖泊唯一的初级生产者[32].因此,达则错沉积物中长链烯酮化合物的总浓度可以指示过去湖泊中初级生产力的变化.过去1000年达则错初级生产力整体呈波动上升的趋势,其中在1050-1250、1450-1600以及1850-1900 AD湖泊初级生产力显著降低.
湖泊沉积物中TOC是描述湖泊沉积物总有机质含量的效指标[13].在以内生作用为主的湖泊中TOC含量有效反映湖泊生产力状况[33].刘莎莎等已经对达则错表层沉积物有机碳构成及其来源进行研究[18],结果表明,达则错沉积物中有机质主要来源于湖泊自生的水生生物碎屑混合物,而陆源高等植物输入的贡献率较低.且沉积颗粒物的镜下鉴别及统计数据显示达则错深水区有机质基本全为内源的西藏拟溞残体[7].因此达则错湖泊沉积基本上是自然过程的内生沉积,湖泊沉积物TOC含量可以反映过去的生产力变化[18].过去1000年,达则错湖泊沉积物TOC含量在1050-1250、1450-1600以及1850-2000 AD相对较高,表明湖泊生产力较大.但在1250-1450以及1600-1850 AD达则错沉积物TOC含量较低,表明湖泊生产力较低.
氮、磷是构成水体初级生产力和食物链最重要的生源要素,也是湖泊富营养化的关键限制性因子,对湖泊生态系统有重要影响[34].达则错湖泊沉积物中TN和TP含量可以反映不同时期湖泊水体的营养状况.过去1000年达则错湖泊沉积物中TN含量的变化幅度大于TP.沉积物中TP含量在1850 AD之前变化较小,自1850 AD之后,沉积物TP含量锐增.沉积物中TN含量在950-1050以及1450-1600 AD较高,自1850 AD之后TN含量显著增加并达到过去1000年最高值.TN、TP含量变化表明过去1000年中达则错在1050-1250以及1450-1600 AD湖泊养养水平较高,而在950-1050、1250-1450以及1600-1850 AD湖泊营养水平较低,在近150年水体富营养化趋势加剧.
3.2 过去1000年达则错生态系统变化及其对气候环境变化的响应机制
RDA分析结果显示, RDA 1轴(λ=0.19)和RDA 2轴(λ=0.04)对响应变量(西藏拟溞和浮游植物生产力)数据变率的解释量为23%(图5).蒙特卡罗置换检验结果显示,P=0.002, 说明气候环境因子对生物因子的影响达到显著水平.环境因子中TN含量对西藏拟溞和浮游植物初级生产力变率的影响最为显著.由图5可以看出,RDA 1轴(横轴)主要反映TN和TP梯度,第1轴从左到右TN和TP含量呈现增加的趋势;RDA 2轴(纵轴)主要反映温度梯度,从上至下温度呈增加的趋势.前人对达则错表层沉积物有机碳含量及构成的研究表明,西藏拟溞是该湖沉积物的有机组分的主要来源,对湖泊TOC含量变化的贡献最大[18].根据RDA排序图(图5)的结果,西藏拟溞丰度变化与TOC含量变化呈正相关关系.相关分析也表明二者之间存在显著的正相关关系(r=0.55,P<0.0001,图6a),表明西藏拟溞对湖泊TOC含量变化影响较大.尽管浮游植物对湖泊沉积物TOC含量变化也有一定的影响,但RDA排序图(图5)以及线性相关分析结果(r=0.07,P=0.5725, 图6d)均表明二者相关性不大,这可能是由于西藏拟溞在繁盛期会摄食大量的藻类,从而削弱了浮游植物对TOC含量的贡献.在1050-1250以及1450-1600 AD西藏拟溞丰度较高的时期,浮游植物藻类含量显著下降(图4 a,b),表明浮游动物对浮游植物的摄食压力较大.
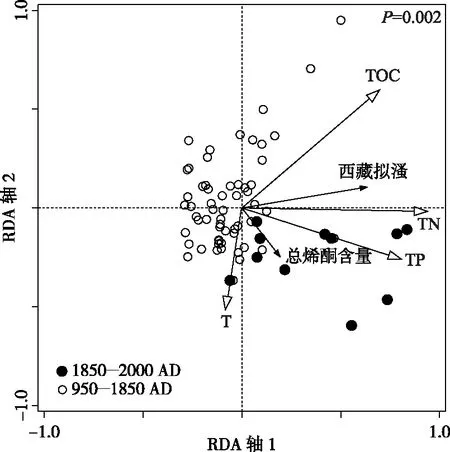
图5 冗余分析三序图
达则错浮游植物生产力变化与TN、TP含量变化呈显著相关性,说明营养盐浓度的高低会直接影响到浮游植物藻类的生长和繁殖(图5,图6e,r=0.3,P=0.0145; 图6f,r=0.47,P<0.0001).其中总烯酮含量与TP含量的相关性高于其与TN含量的相关性,说明磷是该湖藻类种群密度的主要限制营养元素.先前的研究表明, 湖泊沉积物枝角类残余物可以重建湖泊过去的营养状态[35].RDA分析结果以及相关分析均表明达则错沉积物中西藏拟溞丰度与TN、TP等营养盐含量之间存在极显著性相关(图5,图6b,r=0.62,P<0.0001;图6c,r=0.54,P<0.0001),表明该湖泊中西藏拟溞丰度随着湖泊水体营养化程度的加强而升高.达则错为无鱼湖泊,西藏拟溞没有被捕食的压力.随着水体营养元素含量的升高,可食性浮游植物的生物量增加,西藏拟溞丰度明显增加.

图6 达则错西藏拟溞丰度以及总烯酮含量与TOC、TN、TP含量的相关关系
气候变化能够调节湖泊水温、分层时间、冰层覆盖时间和养分浓度,是影响水生生态系统结构特性的重要因素[36-37].气温降低会导致湖水温度下降,且下降幅度在上层湖水(变温层)比深层湖水(均温层)要大得多,还会导致湖泊热稳定性的长期下降、湖水分层期的缩短以及结冰期的延长.达则错烯酮不饱和度指标重建的温度记录显示在过去1000年该区存在2个明显的低温期:1050-1250及1450-1600 AD(图4f).在低温期内,湖水的热稳定性降低,湖底沉积物的氮、磷等营养元素上涌,进而导致湖泊中的TN、TP含量升高(图4c, d).作为浮游植物必需的营养物质,氮、磷含量的增高引起湖泊浮游植物产量增长,进一步提高了湖泊的养分,为西藏拟溞的生长和繁殖创造了有利条件.达则错属于无鱼湖泊,西藏拟溞没有被捕食的压力,湖泊营养浓度的升高会导致西藏拟溞的繁盛.西藏拟溞在繁盛期由于摄食量增加又会消耗掉大量的藻类,导致湖泊浮游植物生产力降低.因此,在1050-1250及1450-1600 AD低温期,达则错湖泊营养盐含量升高,浮游动物优势种西藏拟溞丰度明显增加,而浮游植物藻类因被摄食而数量降低,湖泊总有机质含量升高.反之,在950-1050、1250-1450以及1600-1850 AD等时段,湖区气温相对较高,湖泊热稳定性较好,温跃层深度以下湖水与上部湖水基本不发生混合,湖底营养盐不上涌,湖泊氮磷等营养盐含量低,因食物缺乏、养分浓度低导致西藏拟溞丰度降低.本研究表明自然状况下,气候变化通过改变湖泊理化性质来影响湖泊初级和次级生产力,是影响湖泊生态系统的重要因素.
然而过去150年,达则错湖泊生态系统发生了显著改变.RDA排序结果显示,1850 AD以后的样点主要排列在第四象限,与西藏拟溞丰度、TN、TP和浮游植物生产力的增加以及温度的升高对应,说明1850 AD以来西藏拟溞丰度及浮游植物生产力的增加与环境因子(TN、TP以及温度)呈正相关(图5).过去150年达则错温度与中世纪暖期相当(两时段内的平均温度均为7℃),但是湖泊生产力却显著升高,沉积物TN、TP含量突增并达到过去1000年峰值(图4),这表明此时段达则错湖泊生态系统与之前的时段明显不同,气候变化不再是该时段湖泊生态系统变化的主要驱动力.古湖沼学研究表明,近百年来氮沉降的增加可能是影响偏远高山湖泊养分来源变化并导致湖泊生态系统变化的最重要因素[38-42].加剧的大气污染如氮沉降、酸沉降已经影响到到偏远高山、北极等远离人类活动的区域[43-44].Hu等对青藏高原东南部沙德措和月亮湖两个湖泊沉积岩芯的多指标分析表明,19世纪末期以来大气氮沉降增加是这2个高寒湖泊的生态系统变化的主要驱动因素,而气候因素对这一时期内湖泊生态系统的变化影响较小[38],表明近百年来人类通过改变大气沉降等自然过程对高山湖泊生态系统的间接影响程度增强[44].达则错自1850s以来TN含量的增长趋势与达索普冰芯记录的硝酸盐浓度的上升趋势(图4g)[45]相一致,可以指示全球氮沉降的影响.此外,近期对达则错附近的羌塘1号冰芯的研究表明,近百年来高原中部硝酸盐浓度的上升主要受人类活动导致的氮沉降影响[46],进一步证实人类活动导致的大气沉降可能是该时段达则错湖泊养分含量剧增的主要原因之一.
人类活动导致的污水排放也是影响湖泊TN和TP含量变化的重要原因[47].青藏高原中部的尼玛县城坐落于达则错的入湖河流波仓藏布河畔,位于达则错以西约25 km处.2012年课题组对达则错实地考察,发现当地居民在湖边洗衣,表明人类活动会对湖泊产生直接影响.县城人类活动排放的生活污水以及畜牧业的粪便污水随波仓藏布流入达则错,可能导致该湖近期营养盐含量增加.据地方志记载,今日的尼玛县原属申扎县,直到20世纪后半叶尼玛县城才北迁到现在的位置,在其北迁之前,达则错地区为羌塘无人区,受人类活动直接影响较小[48].因此,人类活动污水排放的直接影响也可能是达则错近期TN、TP含量升高的主要原因.过去150年,氮、磷等营养元素的升高引起湖泊浮游植物产量增长,为西藏拟溞的生长和繁殖创造了有利条件.然而,西藏拟溞的增长也可能会受到同期高温作用的抑制.西藏拟溞为冷水性物种,具有低温适应性,较高温度会使湖泊中触发西藏拟溞季节性繁殖的刺激因素丢失.而且,据研究报道,当湖泊中TP浓度过高时,浮游甲壳动物的生物量并不再随着TP浓度的增加而增加[49].因此,在过去150年,尽管达则错湖泊中营养盐浓度显著升高,但是在1970-1990 AD期间,当营养盐浓度达到一定耐受限度后,湖泊中西藏拟溞丰度不再随着营养盐浓度的增长而增长.综上,在过去150年,达则错湖泊生态系统发生了显著改变,人类活动的直接和间接影响是引起该湖泊生态系统变化的主要原因.此外,气候变化也对湖泊生态系统的变化起了一定的调节作用.
青藏高原属于我国典型的生态环境脆弱地区, 即便少量的污染物输入也有可能对高原生态系统产生较大影响[43].本文结果表明,全球氮沉降以及人类活动的直接影响对青藏高原湖泊生态系统的影响不容忽视.未来在追求地区经济发展的同时要加强对水质的保护,控制人为因素对流域生态系统的干扰,加强区域生态修复和环境保护.
4 结论
青藏高原中部无鱼湖泊达则错沉积岩芯钻孔生源要素记录了过去1000年以来区域的生态系统变化及气候环境演变历史.湖泊生态系统尤其是生产力在自然状况和人类活动影响下存在显著变化.在自然状况下,气候变化通过改变湖泊理化性质来影响湖泊初级和次级生产力,是影响湖泊生态系统的重要因素,较高的湖泊生产力发生在温度较低和营养盐含量较高时期(1050-1250及1450-1600 AD);而较低的湖泊生产力发生于温度较高且营养盐含量较低时段(950-1050、1250-1450以及1600-1850 AD).在过去150年,达则错气温与中世纪暖期相当,但是湖泊生产力却逐渐升高,沉积物TN、TP含量突增并达到过去1000年峰值,这表明此时段达则错湖泊生态系统与之前的时段明显不同,气候变化不再是该时段湖泊生态系统变化的主要驱动力.分析发现大气沉降以及人类活动污水排放的直接影响是过去150年达则错生态系统变化的主要原因.
综上所述,达则错湖泊生态系统在人类活动影响下发生了显著的改变.在千年尺度上驱动高原中部达则错湖泊生态系统变化的主控因素是气候变化,而过去150年来人类活动的直接和间接影响取代气候变化成为影响该湖泊生态系统变化的主导因素.
致谢:感谢暨南大学生态学系林秋奇研究员在野外调查以及湖泊浮游动物鉴定分析工作中给与的帮助,感谢暨南大学生态学系王妙、张华俊博士在总氮、总磷测定工作中给与的帮助.
猜你喜欢
海洋通报(2022年2期)2022-06-30
环境工程技术学报(2022年3期)2022-06-05
中国饲料(2022年5期)2022-04-26
食品安全导刊·中旬刊(2022年3期)2022-04-15
海洋石油(2021年3期)2021-11-05
现代装饰(2020年10期)2020-10-28
空间科学学报(2020年6期)2020-07-21
中国海洋大学学报(自然科学版)(2019年10期)2019-10-14
中国饲料(2019年5期)2019-03-20
小学阅读指南·低年级版(2017年2期)2017-03-23
