
金线莲UGPase基因克隆与表达及在多糖合成中的作用
2021-06-28 01:10:34嵇元烨吴秋丽江晴儿谢晓鸿孔向军王忠华
中草药 2021年12期
嵇元烨,吴 梅,吴秋丽,江晴儿,谢晓鸿*,孔向军*,王忠华*
1.浙江万里学院生物技术研究所,浙江 宁波 315100
2.金华市匠康金草生态农业科技科技有限公司,浙江 金华 321007
金线莲Anoectochilus roxburghii(Wall.)Lindl.是兰科开唇兰属植物,是名贵的中草药,其全草入药,具有清热凉血、除湿解毒的功效,可治疗肺结核咯血、糖尿病、肾炎、膀胱炎、重症肌无力、遗精、风湿性及类风湿性关节炎、小儿惊风、妇女白带以及毒蛇咬伤等症[1]。研究表明,多糖是金线莲主要的药用活性成分之一,金线莲多糖具有降血糖[2]、抗氧化[3]、抗肿瘤[4]、保肝[5]、免疫调节[6]等生物活性。
在植物糖代谢途径中,UGPase基因主要催化尿苷三磷酸和葡萄糖-1-磷酸,形成尿苷二磷酸葡萄糖和PPi(Glucose 1-phosphate+UTP⇌PPi+UDPG),这是一个依赖Mg2+的可逆反应,其反应产物UDPG可以参与植物中蔗糖合成,纤维素合成以及淀粉合成等,并为植物细胞壁合成提供前体物质[7],同时UGPase 是合成植物多糖的关键酶[8]。目前,已从马铃薯Solanum tuberosumL.[9]、大麦Hordeum vulgareL.[10]、小果野蕉Musa acuminataColla[11]、水稻Oryza sativaL.[12]、香瓜Cucumis meloL.[13]、棉花Gossypium hirsutumL.[14]、小麦Triticum aestivumL.[15]等植物中分离出UGPase基因。目前国内外关于金线莲多糖合成途径中的关键基因的报道较少,而UGPase基因在金线莲多糖合成代谢途径中的调控作用尚未明确。本研究采用实时荧光定量PCR(Quantitative Real-time PCR,qRT-PCR)技术和RACE(rapid-amplification of cDNA ends)技术,首次从金线莲中克隆出UGPase基因全长,并用生物信息学手段对该序列进行分析验证。同时分析了其在7 个金线莲品种中组培期与种植期茎段中的表达量、酶活与多糖含量的相关性。为深入研究UGPase基因在金线莲多糖合成途径中的分子调控作用和通过基因工程手段特异性调控关键酶基因从而促进代谢产物大量合成积累提供基础。
1 材料与仪器
1.1 材料收集与处理
本研究以由2019年下半年同批的7 个不同品种金线莲(尖叶、小叶、圆叶、健君、无纹、匠康1 号、匠康2 号)的组培苗以及种植苗的茎段为材料,具体差异见图1。其中组培苗采自2019年11月,组培时间3 个月;种植苗采自2020年5月,经炼苗后种植时间6 个月,材料收集自金华市农业科学研究院,经浙江万里学院生物与环境学院王忠华教授鉴定为兰科开唇兰属植物金线莲A.roxburghii(Wall.)Lindl.的茎段部位。新鲜材料经过清水冲洗后,用酒精擦拭及DEPC 处理水洗涤后,放入液氮中速冻,然后置于-80 ℃冰箱中保存备用。

图1 不同品种金线莲样品差异Fig.1 Samples differences in different varieties of Anoectochilus roxburghii
1.2 仪器
MastercyclerproS 梯度PCR 仪、5430R 低温高速离心机、微量移液器(Eppendorf 公司);GelDocXR+凝胶成像仪、CFX96实时荧光定量PCR(Bio-RAD公司);NanoDropTM2000 分光光度计(Thermo Fisher Scientific公司);蒸汽高压灭菌锅(SANYO 公司);超低温冰箱(New Brunswick);电子分析天平(Mettler Toledo 公司);电泳仪(北京六一生物科技公司);紫外分光光度计(上海天能科技有限公司);恒温金属浴(上海生工有限公司);冷冻恒温摇床(上海智城有限公司);智能生化培养箱(宁波江南仪器厂)。
2 方法
2.1 总RNA 提取、cDNA 第1 链的合成及RACE cDNA 的合成
以尖叶金线莲组培苗茎段为材料,按北京金线莲茎段的总RNA。采用NanoDropTM 2000 分光光度计(Thermo Fisher,美国)测定总RNA 的质量和纯度,1.5%琼脂糖凝胶电泳鉴定其完整性。使用北京全式金公司的TransScript One-Step gDNA Removal and cDNA Synthesis SuperMix 第1 链合成试剂盒反转录合成cDNA 第1 链。使用TaKaRa SMARTer® RACE 5’/3’ Kit 试剂盒合成 5’及3’RACE cDNA,-20 ℃保存。
2.2 简并引物设计与合成、PCR 扩增UGPase 基因序列及信息生物学分析
根据GenBank 中已注册的植物UGPase基因部分或全长序列(基因编号 JX294909.1、XM_020741275.1、FJ536261.1、XM_021452444.1等),采用MEGA7.0 软件的多序比对功能检索不同植物UGPase基因序列中重合度较高的部分,设计出简并引物:UGPase-F:5’-GAGTGGAGYAAGATYCAGAC-3’,UGPase-R:5’-AACCAKACATCACCAGARACCTTCA-3’。目的产物约1170 bp。引物由北京擎科新业生物技术有限公司杭州合成部合成。
以“2.1”项获得的cDNA 为模板,UGPase-F、UGPase-R 为上下游引物进行PCR 扩增。PCR 扩增体系为:50 μL 反应体系,含2×Flash Hot Start MasterMix 25 μL、上下游引物各2 μL、cDNA 模板2 μL、ddH2O 19 μL。PCR 反应条件为:94 ℃预变性2 min;94 ℃变性30 s,51.7 ℃退火30 s,72 ℃延伸1 min,35 个循环;72 ℃延伸 2 min。PCR 产物经1%琼脂糖凝胶电泳后,切下目的片段,经北京全式金公司的EasyPure Quick Gel Extraction Kit琼脂糖凝胶DNA 回收试剂盒纯化后与PMD 18-T载体进行连接,将重组载体转入到DH5α 大肠杆菌感受态细胞中。过夜培养后筛选8 个阳性菌落进行测序分析,将测序结果进行BLAST 比对分析,验证测序结果的正确性。
根据上述测序得到的UGPase基因保守区序列,利用Primer5 设计特异性引物,3’RACE 引物为3-UGPase 5’-GGTGGCACGCTAATCTCATATGAAGGAAG-3’,5’ 的 RACE 引物为 5-UGPase 5’-ATGGGAAGACATCACCGTGCCCTGGAG-3’。2次RACE-PCR 反应体系均为50 μL,包括SeqAmp DNA Polymerase 1 μL、2×SeqAmp Buffer 25 μL、PCR-Grade H2O 15.5 μL、5’or3’RACE cDNA 2.5 μL、3-UGPaseor5-UGPase 1 μL、10×UPM 5 μL。PCR程序为5’RACE:94 ℃、30 s,72 ℃、3 min,5个循环;94 ℃、30 s,70 ℃、30 s,72 ℃、3 min,5 个循环;94 ℃、30 s,68 ℃、30 s,72 ℃、3 min,30 个循环;4 ℃保温。3’RACE:94 ℃、30 s,68 ℃、30 s,72 ℃、3 min,30 个循环;4 ℃保温。后续实验步骤同保守区克隆过程。
得到UGPase基因完整序列后,使用ExPASy中的Translate 工具将基因序列翻译成氨基酸序列,使用ProtParam 软件分析蛋白质的基本性质。使用Net Phos 3.1 Server 软件分析蛋白质的磷酸化位点,使用NetOGlyc 4.0 软件分析蛋白质的糖基化位点。使用TMHMM-2.0 软件分析蛋白质的跨膜情况,使用SignalP-5.0 软件分析蛋白质的信号肽,使用SoftBerry 网站分析蛋白质的亚细胞定位。使用SOPMA 软件分析蛋白质的二级结构,使用SWISS-MODEL 软件分析蛋白质的三级结构。在NCBI 中的Conserved Domain Database(CDD)在线软件中分析蛋白质结构功能域,最后在MEGA7.0软件中使用邻位相连法(neighbor-joining)构建UGPase 蛋白的系统进化树。
2.3 UGPase 基因表达量、相关酶活性与不同金线莲多糖含量测定及分析
按照“2.1”项的方法提取7 个不同品种金线莲组培期及种植期茎段qRT-PCR cDNA,按照“2.2”项得到的测序结果,设计 qRT-PCR 引物,UGPase-YGF:5’-GATTATCCCAAACCCAAAGG-3’,UGPase-YGR:5’-AGAGGGGTTAGCAGGATTTG-3’;预计扩增长度232 bp。选择金线莲Actin-2[16]基因作为内参基因,Actin-2-F:5’-CGGGCATTCACGAGACCAC-3’,Actin-2-R:5’-AATAGACCCTCCAATCCAGACACT-3’。分别以不同品种的茎段cDNA 为材料,UGPase-YGF、UGPase-YGR 为上下游引物进行qRT-PCR 反应,反应体系共20 μL:Tip Green qPCR SuperMix 10 μL,上、下游引物以及DNA 模板各0.4 μL,ddH2O 8.8 μL。扩增程序:94 ℃预变性30 s,94 ℃变性5 s,60 ℃退火30 s,共40 个循环;qRT-PCR 结果按照公式2-ΔΔCt[17]计算。
以7 个不同品种金线莲茎段液氮研磨后离心取上清液为材料,采用上海科兴公司的酶联免疫分析(ELISA)试剂盒数据检测不同品种金线莲组培期及种植期茎段中的UGPase 酶酶活性。另外以个不同品种金线莲茎段固体粉末为材料,采用苯酚-硫酸法[18]测定不同品种金线莲茎段中多糖含量。分析采用SPSS24.0 软件进行单因素方差分析并采用Pearson 双变量相关性法[19]结合多糖含量、基因表达量及酶活进行修改下分析。
3 结果与分析
3.1 总RNA 的提取及金线莲UGPase 基因的克隆
金线莲总RNA 的琼脂糖凝胶电泳结果见图2-D,可清晰地看到2 条RNA 条带(28 S 和18 S)其中28 S 条带亮度大约是18 S 条带的2 倍,A260/A280值在2.08,可进行下一步的cDNA 第一链合成实验[20]及RACE cDNA 合成实验。测序得到中间序列1160 bp(图2-A)、5’端序列657 bp(图2-B)和3’端序列924 bp(图2-C)。拼接各片段,得到UGPase基因cDNA 序列全长1786 bp。开放阅读框为1425 bp,编码475 个氨基酸,cDNA 全长中还包含5’非编码区53 bp,3’非编码区308 bp,以及起始密码子ATG、终止密码子TGA 和多聚A 尾。将全长序列上传至GENBANK,获得NCBI 登录号MT978185。

图2 UGPase基因全长序列的克隆及总RNA琼脂糖凝胶电泳Fig.2 The cloning of full nucleotide sequences of UGPase gene
3.2 金线莲UGPase 基因信息生物学分析
通过Protparam 软件预测得到:UGPase 蛋白相对分子质量为64 996.47,理论pI 为6.40,氨基酸中亮氨酸(Leu)占比最高,达到10.8%,分子式为C2969H4711N757O846S14。在体外哺乳动物网织红细胞中预测半衰期为30 h,不稳定指数为34.49,属于稳定蛋白,脂肪系数为108.41,亲水性平均值为0.096,属于疏水蛋白。通过Net Phos 3.1 Server及NetOGlyc4.0 软件预测得到:UGPase 蛋白有磷酸化位点47个,其中丝氨酸24 个,苏氨酸17 个,酪氨酸6 个。有糖基化位点4 个,分别在第67、71、73 及82 个氨基酸。通过SignalP-5.0、TMHMM-2.0 及SoftBerry网站预测得到:UGPase 蛋白存在信号肽,位置可能位于第22~23 个氨基酸,表明该蛋白为分泌型蛋白,同时该蛋白存在跨膜位点,跨膜区可能位于第494~516 个氨基酸,表明该蛋白为跨膜蛋白,亚细胞定位系数表明,UGPase 蛋白定位于细胞质的可能性最高。通过SOPMA及SWISS-MODEL软件预测得到:UGPase 蛋白具有203 个α-螺旋(alpha helix),占比34.18%;137 个延伸链(extended strand),占比23.06%;49 个β-转角(beta turn),占比8.25%;205 个无规卷曲(random coil),占比34.51%,三级结构预测结果见图3。通过CDD 软件分析得到:UGPase 蛋白属于糖基转移酶家族A(Glyco_tranf_ GTA_type)成员之一,具有PLN02474(位于第22~486 位氨基酸)、UDPGP(位于第48~458 位氨基酸)、UGPase_euk(位于第97~396 位氨基酸)及QRI1(位于49~486 位氨基酸)4 个结构域。

图3 金线莲UGPase 蛋白三级结构Fig.3 Tertiary structure of UGPase protein from Anoectochilus roxburghii
通过 BLAST 氨基酸序列比对发现金线莲UGPase 氨基酸序列与铁皮石斛Dendrobium officinaleKimura et Migo(AGA17038.1)、蝴蝶兰Phalaenopsis equestris(Schauer)Rchb.f.(XP_ 020596934.1)、深圳拟兰Apostasia shenzhenicaZ.J.Liu&L.J.Chen(PKA66203.1)、芦笋Asparagus officinalisL.(XP_020273555.1)、油棕Elaeis guineensisJacq.(XP_010935335.1)、糜子Panicum miliaceumL.(RLN32892.1)、海枣Phoenix dactyliferaL.(XP_008804801.1)、菠萝Ananas comosus(Linn.)Merr.(OAY68359.1)、狗尾草Setaria viridis(L.)Beauv.(XP_034583618.1)、高粱Sorghum bicolor(L.)Moench(XP_ 002453185.1)和甘蔗Saccharum officinarumL.(ACL80329.1)的相似性分别达到88.60%、87.96%、86.51%、85.31%、85.07%、84.55%、84.01%、83.80%、82.86%、82.43%和85.34%。其中金线莲UGPase 氨基酸序列与同属兰科的铁皮石斛、同源性最高(图4)。选取了一部分典型的单子叶植物使用MEGA7.0 软件构建系统进化树(图5),进行分子系统学分析,系统进化树结果显示,金线莲与铁皮石斛、蝴蝶兰、深圳拟兰、油棕、海枣聚为一大支,其中进行与铁皮石斛、蝴蝶兰具有最近的亲缘关系,这可能由于3 种植物都属于兰科,所以亲缘较近。

图4 金线莲UGPase 氨基酸序列与其他植物比对结果Fig.4 Results of comparison of UGPase amino acid sequence with other plants

图5 12 种植物UGPase 蛋白进化树Fig.5 Evolutionary tree of UGPase protein in 12 plant species
3.3 不同品种金线莲多糖含量测定
不同品种金线莲茎段多糖含量测定结果见图6所示。结果显示,在这7 个品种中,金线莲茎段多糖变化趋势均为明显上升,且每个品种在2 个时期中茎段多糖含量均不相同,在组培期茎段中多糖含量最高的是无纹品种,最低的是匠康2 号品种,所有品种之间均存在显著差异,而在种植期茎段中多糖含量最高的是匠康1 号,最低的是匠康2 号,所有品种之间均存在显著差异。从组培期至种植期多糖增加最多的是小叶品种,最少的是匠康2 号品种,多糖含量最少增加了1 倍以上,最多达到了9 倍。
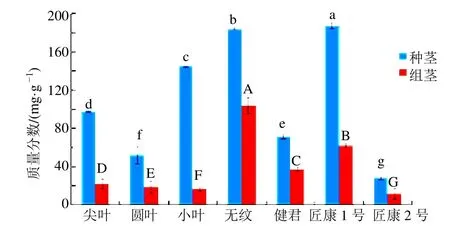
图6 不同品种金线莲茎段多糖含量测定Fig.6 Determination of polysaccharides in different varieties of Anoectochilus roxburghii
3.4 不同品种金线莲UGPase 基因的表达分析
不同品种金线莲UGPase基因荧光定量PCR 实验结果结果如图7所示,结果显示,UGPase基因在7个不同品种金线莲2个时期的茎段中均有表达,但在不同时期具有有不同的表达水平。在组培期,茎段中表达最高为无纹品种,最低为圆叶品种,其中无纹品种与匠康1 号、尖叶品种表达量有显著差异,而这2 个品种与健君、小叶和匠康2 号3 个品种茎段表达量有显著差异,最后表达量最小的圆叶品种和前面3 个品种有显著差异。种植期茎段表达量最高同样是无纹品种,最低为匠康2 号品种,无纹品种与匠康1 号品种与其他5 个品种茎段表达量有显著性差异。另外在组培期到种植期之间,7 个品种金线莲茎段部位的表达量均呈现上升趋势,且增加趋势较为明显。
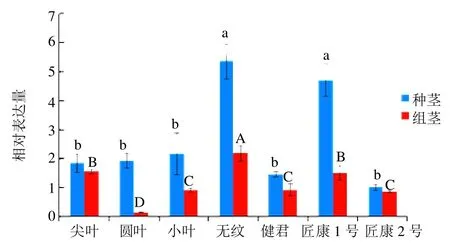
图7 不同品种金线莲UGPase 基因茎段表达量Fig.7 Relative expression levels of UGPase of stem in different varieties of Anoectochilus roxburghii
3.5 不同品种金线莲UGPase 酶的酶活性测定
不同品种金线莲茎段UGPase 酶活性测定结果如图8所示。结果显示,UGPase 酶在7 个不同品种金线莲2 个时期的茎段中均有活性,但在不同时期酶活变化不大。在组培期,茎段中酶活最高为圆叶品种,最低为匠康2 号品种,7 个品种茎段部位酶活均无显著差异。种植期茎段酶活最高匠康1 号品种,最低为匠康2 号品种,匠康2 号品种较其他6 个品种茎段酶活略低。另外在组培期到种植期之间,7 个品种的金线莲茎段部位酶活均呈现上升趋势,但没有出现大幅度增加的情况。
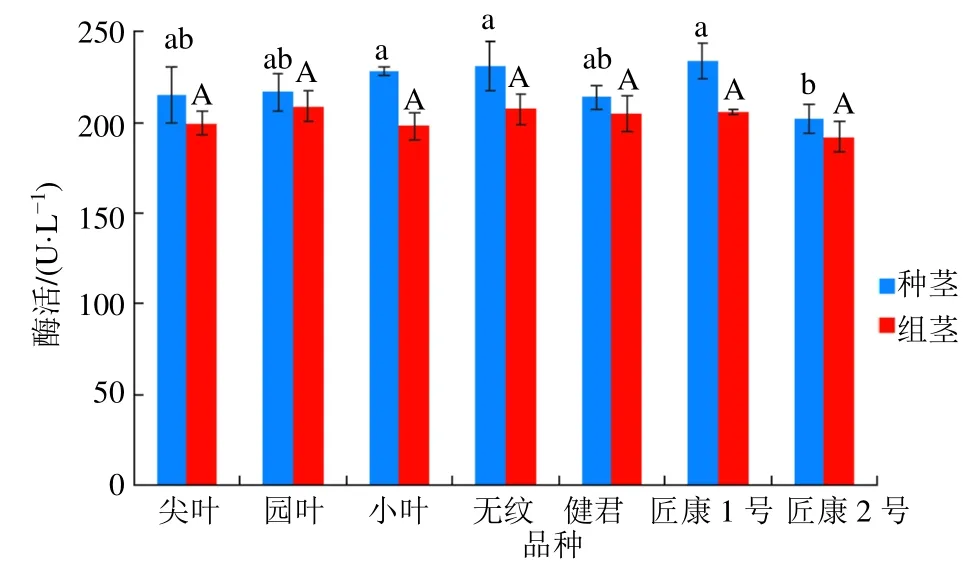
图8 不同品种金线莲UGPase 酶茎段酶活Fig.8 Enzymatic activity of UGPase of stem and leaf in different varieties of Anoectochilus roxburghii
3.6 不同品种金线莲多糖与表达、酶活性相关性分析
通过SPSS 软件对实验结果进行相关性分析,结果如表1所示。不同品种不同时期金线莲茎段部位多糖含量变化均与UGPase基因表达量以及酶活呈极显著正相关性,而表达量与多糖含量的相关系数要高于酶活,另外,UGPase基因表达量与单独组培时期以及单独种植时期不同品种金线莲茎段部位多糖含量均呈极显著正相关,相关系数较与不同品种不同时期金线莲茎段部位多糖含量略小。但酶活只与单独种植时期不同品种金线莲茎段部位多糖含量呈极显著正相关,与单独组培时期多糖含量并未有显著相关性,而且与种植期多糖含量相关系数同样小于与不同品种不同时期金线莲茎段部位多糖含量的相关系数。

表1 金线莲多糖含量与UGPase 基因表达量及酶活相关性分析Table 1 Analysis of correlation between polysaccharide content and expression of UGPase gene or enzyme activity in Anoectochilus roxburghii
4 讨论
植物多糖具有调节免疫功能、抑制肿瘤、延缓衰老、抗疲劳、降血糖、抗辐射、抗菌抗病毒及保肝等作用。为了利用植物基因工程、细胞工程工业化大量生产该重要活性成分,多糖合成途径及调控的关键酶基因成为研究热点,植物多糖合成途径是植物糖代谢相关途径的一部分,植物的糖代谢相关途径是整个植物生物代谢的中心,它勾通了蛋白质代谢、脂类代谢、核酸代谢及次生物质代谢。尿苷二磷酸葡萄糖焦磷酸化酶(UGPase)是植物糖代谢途径中的一个关键酶,其首先由Munch-Petersen、Kalckar 和Cutolo 于1953年在酵母细胞中发现[21],近年来,国内外学者已在党参[22]、白花泡桐[23]、龙须菜[24]、黄芪[25]、铁皮石斛[26-27]、落叶松[28]、海带[29]和紫穗槐[30]等植物中克隆分离出UGPase基因。它催化反应UTP+葡萄糖-1-P⇌UDP-葡萄糖+PPi,在不同的植物组织中,其催化方向有所不同。在源组织中,二氧化碳经卡尔文循环固定生成磷酸丙糖后,将其转化为果糖-1-6-二磷酸,然后在磷酸酶、己糖磷酸变位酶以及葡萄糖磷酸变位酶的催化下生成葡萄糖-1-磷酸随后经UGPase 催化生成UDP-葡萄糖,为蔗糖的生物合成提供底物其产物[31],在库组织中,催化蔗糖的降解,即催化UDP-葡萄糖生成葡萄糖-1-磷酸[32]。UGPase 还与胞液中的腺苷二磷酸葡萄糖焦磷酸化酶(ADP-glucose pyrophosphorylase,AGPase)相偶联,形成腺苷二磷酸葡萄糖(ADPG),参与造粉体淀粉的合成[10,33]。生物体合成多糖的前提是单糖以活化的形式存在,使其可以被充分利用,UDP-Glc 作为活化糖,它在被UGPase 经葡萄糖催化生成后充分参与到植物中多种多糖类物质的合成中,是植物多糖合成的关键酶。
本研究利用已报到的其他植物UGPase基因序列,首次从兰科植物金线莲中克隆出UGPase基因全长序列,该基因全长1786 bp。开放阅读框为1425 bp,编码475 个氨基酸,在NCBI 上进行在线BLAST 序列比对,UGPase 氨基酸序列同源性比对及系统进化树分析显示,金线莲UGPase 氨基酸片段与其他植物有较高的同源性(88.60%~82.43%),其中与铁皮石斛等兰科植物同源性最高,表明UGPase基因编码区高度保守,与前人报道的结果非常相似[34]。另外李帅玲[35]研究得出金线莲是多糖积累型植物,其主要积累部位为根茎,其他部位多糖几乎可忽略不计,因此本研究选择7 个不同品种金线莲的茎段来作为实验材料。在本研究中发现,不同品种金线莲之间多糖含量具有显著性差异,这与魏翠华等[36]、施满容等[37]研究结果类似。UGPase基因表达量在不同品种金线莲茎段中与多糖含量呈极显著正相关,这与在铁皮石斛[38]的类似研究中结果一致。此外,UGPase 酶活性也同样与多糖含量存在极显著正相关,不过相关系数较表达量小。由此可以初步判断UGPase基因确实参与金线莲多糖合成代谢途径,在主要多糖积累部位参与调控多糖的合成积累。
本研究以尖叶金线莲茎段为材料,克隆出金线莲UGPase基因全长。通过氨基酸序列同源性分析及系统进化树等的方法,可推断克隆得到的序列片段就是金线莲UGPase基因的cDNA 片段。另外通过测定7 个不同品种金线莲茎段UGPase基因表达量及相关酶活,并与多糖含量进行相关性分析,初步证明UGPase基因是参与金线莲多糖合成代谢的关键基因之一。这为金线莲多糖的合成代谢途径研究提供了必要的前期准备,并为今后通过分子生物学手段增加其多糖含量奠定基础。
利益冲突所有作者均声明不存在利益冲突
猜你喜欢
中国果业信息(2023年12期)2024-01-09 06:59:14
小学生作文(低年级适用)(2022年11期)2022-12-02 09:01:38
湖北农业科学(2021年13期)2021-08-04 03:56:54
World Journal of Clinical Cases(2020年9期)2020-05-13 07:34:08
种子(2019年4期)2019-05-28 02:04:10
福建农业科技(2018年11期)2019-01-17 08:16:36
中成药(2018年10期)2018-10-26 03:41:08
天然产物研究与开发(2018年6期)2018-07-09 06:01:28
中成药(2018年3期)2018-05-07 13:34:45
中成药(2017年7期)2017-11-22 07:33:56
