
香椿优树半同胞家系在秦巴山区生长变异与早期选择
2021-06-28 07:09代锐焦怀明何志翔杨勇智郑海宏彭建肖兴翠
四川林业科技 2021年3期
代锐, 焦怀明, 何志翔, 杨勇智, 郑海宏, 彭建, 肖兴翠*
1.旺苍县林业科技推广服务中心,四川 广元 628200;
2.旺苍县林产有限公司,四川 广元 628200;
3.四川省林业科学研究院,四川 成都 610081;
4.九寨沟县林业和草原局,四川 阿坝 623400
香椿(Toona sinensis)是楝科香椿属落叶乔木,是我国特有的速生珍贵用材树种,生长迅速,树干通直,木材色泽红润、花纹美丽,且具有香味,有“中国桃花心木”之称[1]。由于香椿比桢楠、柏木等慢生珍贵树种速生,因此四川省从2000年开始,就把香椿作为优先发展的6个珍贵用材树种之一,在川中丘陵区、川南地区优先发展,并在退耕还林中大量使用,现在又将香椿作为世行造林、储备林建设的主要造林树种之一。经过近20年的发展,四川香椿人工林面积已有近百万亩。然而前期香椿造林因良种缺乏,种苗质量差,造林效果不好。尽快选育出适合四川各区域的香椿良种,成为香椿研究的重点之一。
在香椿良种选育方面,国内外学者在优良家系早期选择方面开展了一些研究,刘正周等[2]在河南南阳对来自4个省9个种源的香椿家系10年生时进行了优良种源选择,宋志姣[3]在广西南宁对来自9个种源32个香椿家系3年生时开展了种源早期选择,张海燕[4]在福建邵武对来自 14 个省区 29 个香椿种源4年生时进行了早期生长评价,并对70 个香椿优树半同胞子代进行了早期优良家系选择[5],左继林等在江西瑞昌对来自11个不同种源的香椿造林1年时进行了早期种源选择[6,7],高楠等[8]在福建永春和华安对来自12个种源的 70 个香椿优树子代造林2年时进行了早期优良家系选择,胡继文等在和河南对49个香椿无性系造林3年时进行了早期选择[9]。
四川省香椿良种选育相对起步较晚,主要是从“十三五”开始进行种质资源收集,对收集的省内外半同胞家系进行了分家系育苗,先后进行了苗期生长比较与优良家系选择[10,11],2017年开始先后在蓬溪、旺苍和合江建立了区域化试验林,旨在选育出适合不同区域的香椿良种,为香椿造林提供良种。本文以栽植在旺苍县的34个香椿半同胞家系为研究对象,对造林3年的生长和变异进行了分析,进行优良家系早期选择,以为后期的良种选育提供参考。
1 材料与方法
1.1 试验地概况
试验地位于广元市旺苍县的国营苗圃 (106°5'E,32°4'N),海拔530 m~550 m。旺苍县属亚热带湿润季风气候。因特殊的地理地貌影响,四季分明,雨量充沛,光热资源丰富,无霜期较长,山地气候明显。全年平均气温16.1 ℃,全年无霜期260 d,年平均降雨量1 200 mm,年平均日照时数为1 355.3 h。苗圃地的土壤为黄壤土,pH值5.15,表层土有机质23.50 g·kg-1,水解氮 170.2 mg·kg-1,有效磷 10.34 mg·kg-1,速效钾 146.9 mg·kg-1,土壤肥力中等。
1.2 试验材料及来源
参试的34个香椿半同胞家系主要来源于四川朝天、万源、蓬溪、洪雅、筠连、沐川、马边、陕西、广西灌阳及福建霞浦10个种源,分别是四川省林科院和亚林所、福建省林科院在这些地方选择的香椿优树的半同胞家系,优树所在地理位置及生长环境基本情况见表1。
1.3 试验设计
2016年3 月,对34个香椿半同胞家系进行分家系育苗,培养1年生轻基质容器苗,平均苗高25 cm,地径3 mm。2017年3月进行造林,随机区组设计,株行距为2 m×3 m,5株小区,4次重复。造林前进行穴状整地,穴的规格为50 cm×50 cm×40 cm。造林1~3年每年5—9月进行除草2~3次。造林第2年追施有机无机复混肥250 g·株-1,造林第3年分两次追施高氮复合肥500 g·株-1。
1.4 数据测量及分析
2020年11 月,测量胸径和树高。采用Excel 2010进行数据统计,采用SPSS 17.0进行方差分析及LSD多重检验。其中单株材积采用四川省和滇西北阔叶树二元立木材积表,V=5.2750716×10-5D1.9450324H0.9388533[12]。
遗传变异系数:C=S/X×100%,式中:S为标准差;X为某一性状的群体平均值;
家系遗传力h2=(MSv-MSt)/MSv×100,式中:MSv为处理间均方;MSt为处理内均方[13,14];
遗传增益△G=h2Si/X,现实增益G/%=(Xi-X)/X×100,式中,h2为性状的遗传力,Si为家系选择差,Xi为个体平均数[15]。
2 结果与分析
2.1 不同家系生长与种源环境相关性分析
由表2 可见,在家系生长方面,胸径与树高和单株材积都有极显著(P<0.01)的相关性,树高和单株材积之间也有极显著的相关性。在与优树地理位置和生长环境方面,总体而言,胸径、树高和单株材积与经度、纬度和海拔呈负相关性,与年均气温、降雨量和无霜期呈正相关性。即随着优树种源所在地经纬度的增大、海拔的增高,优树半同胞家系的生长越差,而随着优树种源所在年均气温的增高、降雨量的增大、无霜期变长,优树半同胞家系的生长越好。其中地理位置对香椿半同胞家系胸径、树高生长的影响大小为纬度>海拔>经度,气候对香椿半同胞家系胸径、树高、单株材积生长的影响大小为无霜期>年均气温>降雨量。其中胸径与经度、纬度、海拔均有极显著的负相关性,与年均气温、无霜期均有极显著的正相关性;树高与纬度、海拔均有极显著的负相关性,与年均气温、无霜期均有极显著的正相关性;单株材积与经度和海拔均呈显著(P<0.05)的负相关性,与年均气温、无霜期均有极显著的正相关性。
2.2 不同家系生长变异
由表3可见,34个香椿半同胞家系间的胸径、树高、单株材积和林分蓄积均有极显著差异(P<0.01),表明香椿不同家系间变异很大,有很好的早期选择潜力,可开展优良家系早期选择。胸径、树高、单株材积的遗传力分别为0.830、0.850和0.777,遗传力均较大,树高的遗传力略大于胸径和单株材积。
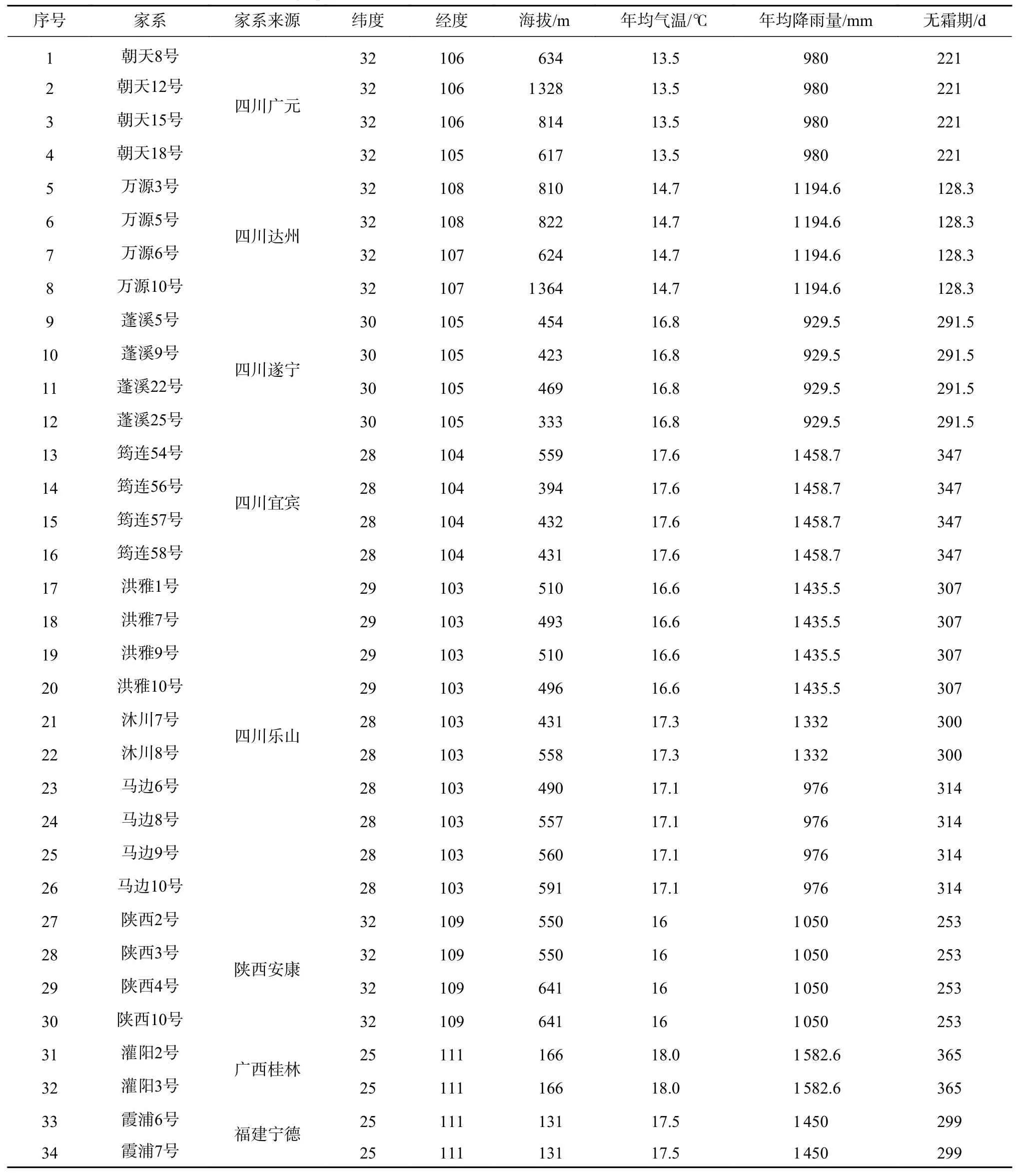
表1 香椿不同家系所在地理位置及气候条件Tab.1 Geographical location and climatic conditions of different Toona sinensis families

表2 香椿家系生长与环境因子相关性分析Tab.2 Correlation analysis between growth and environmental factors of Toona sinensis families
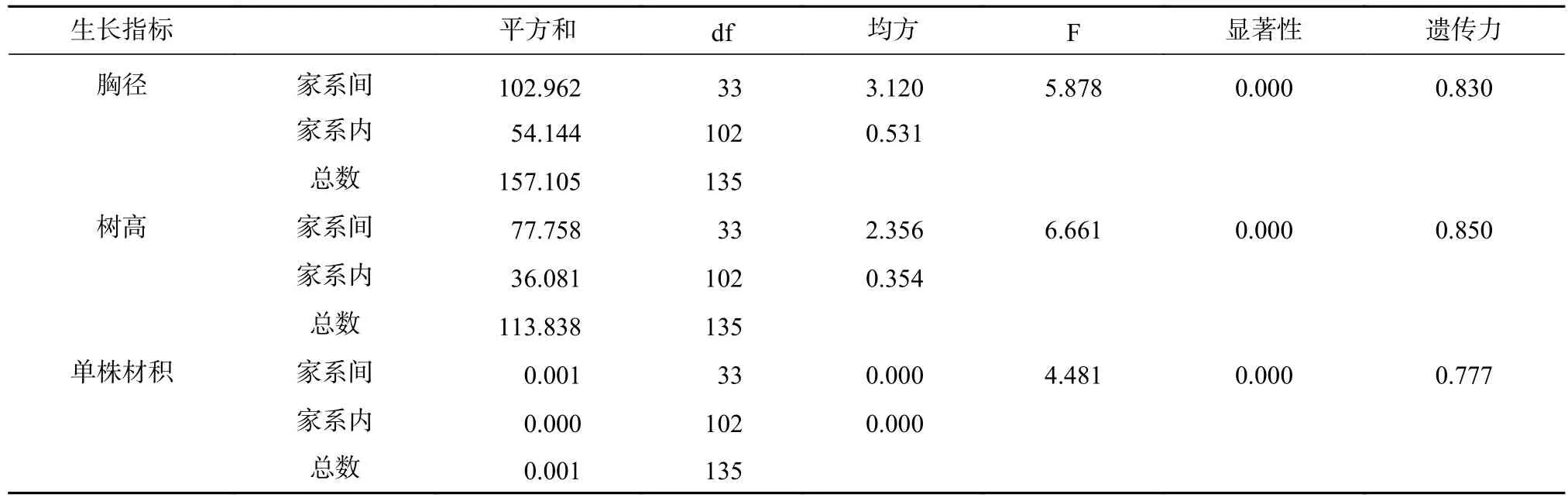
表3 香椿不同家系 3 年生长量方差分析Tab.3 Variance analysis of 3-year growth data of different Toona sinensis families
2.2.1 不同家系胸径生长变异
由表4可见,34个香椿半同胞家系的平均胸径为4.81 cm,超过平均值的家系有20个。其中以蓬溪22号的胸径最大,在5.70 cm~6.94 cm之间,平均为6.35 cm,年均胸径2.12 cm,遗传增益为26.47%,显著(P<0.05)高于胸径≤5.08 cm的18个家系,与胸径在5.15 cm~6.06 cm之间的15个家系间有差异但差异不显著。其次是洪雅9号,在5.40 cm~6.68 cm之间,平均胸径为6.06 cm,年均胸径2.02 cm,遗传增益为21.51%,与胸径≤4.78 cm的14个家系间有显著差异,与蓬溪22号及胸径在4.83 cm~5.89 cm之间的19个家系间没有显著差异。排在第三的是蓬溪 25号和沐川 7号,分别在 4.90 cm~8.30 cm及5.64 cm~6.20 cm之间,平均胸径均为5.89 cm,年均胸径1.96 cm,遗传增益分别为18.59%和18.67%,显著高于胸径≤4.58 cm的13个家系,与蓬溪22号、洪雅9号和胸径在4.78 cm~5.69 cm之间的18个家系间均没有显著差异。34个香椿半同胞家系胸径的平均变异系数为22.43%,其中除了蓬溪25号、洪雅1号、筠连54号、马边6、9、10号、朝天8、12、15、18号、陕西3、10号这12个家系的变异系数大于15%外,其余22个家系的变异系数均小于15%,表明绝大部分家系生长较为均匀一致,有利于家系选择。
2.2.2 不同家系树高生长变异
由表5可见,34个香椿半同胞家系的平均树高为4.30 m,超过平均值的家系有20个,其中以洪雅9号的树高最大,在5.05 m~5.82 m之间,平均为5.33 m,年均树高生长量为1.78 m,遗传增益为20.26%,显著(P<0.05)高于树高≤4.23 m的14个家系,与树高在4.32 m~5.24 m之间的19个家系间没有显著差异。洪雅9号的变异系数为6.7%,表明树高生长较为均匀。其次是蓬溪22号、筠连56号、霞浦7号、灌阳2号、蓬溪25号,树高在4.20 m~6.40 m之间,年均树高生长量为1.70 m~1.75 m,遗传增益在16.06%~18.58%,显著高于树高≤4.09 m的13个家系,5个家系间及与洪雅9号和树高在4.23 m~5.03 m的15个家系之间均没有显著差异。5个家系中灌阳2号和蓬溪25号的树高变异系数大于15%,表明这两个家系的树高生长略微不整齐,有待继续观测。34个香椿半同胞家系树高的平均变异系数为21.36%,其中除了灌阳2号、蓬溪25号、洪雅 1号、万源 5号、筠连 54号、马边 6、9、10号、朝天12、15、18号、陕西3、10号这11家系的变异系数大于15%外,其余23个家系的变异系数均小于15%,树高生长较为均匀一致,有利于家系选择。
2.2.3 不同家系单株材积生长变异
由表6可见,34个香椿半同胞家系的平均单株材积为0.0050 m3,其中以蓬溪22号的单株材积最大,平均为0.0093 m3,大小在0.0070 m3~0.0120 m3之间,遗传增益高达67.29%,显著(P<0.05)高于单株材积≤0.0053 m3的18个家系,与单株材积在0.0058 m3~0.0085 m3之间的15个家系间均没有显著差异。其次是蓬溪25号,单株材积平均为0.0085 m3,大小在0.0045 m3~0.0179 m3之间,遗传增益为55.93%,显著高于单株材积≤0.0048 m3的15个家系,与蓬溪22号及单株材积在0.0050 m3~0.0083 m3之间的15个家系间均没有显著差异。排在第3位的是洪雅9号,单株材积平均为0.0083 m3,大小在0.0060 m3~0.0110 m3之间,遗传增益为51.62%,显著高于单株材积≤0.0043 m3的14家系,与蓬溪22号、洪雅9号及单株材积在0.0048 m3~0.0075 m3之间的18个家系间均没有显著差异。单株材积是胸径和树高累积效应的结果,34个半同胞家系的变异系数均大于15%,平均为57.28%,表明不同家系在3年时的材积生长存在一定的差异,个体存在一些分化,有待继续观测。
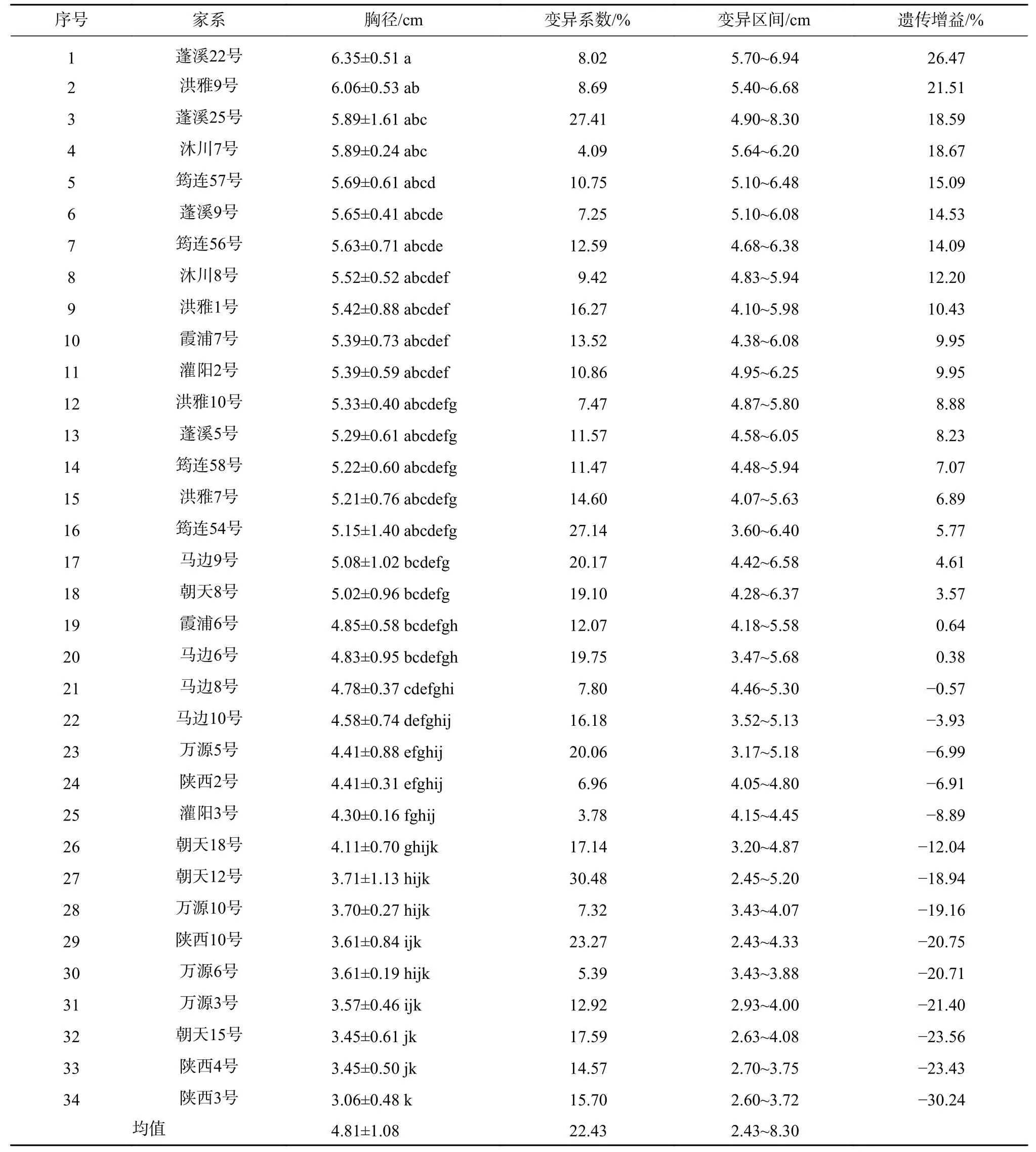
表4 香椿不同家系胸径生长变异Tab.4 Variation of DBH growth of different Toona sinensis families

表5 香椿不同家系树高生长变异Tab.5 Variation of tree height growth of different Toona sinensis families
2.3 不同家系生长比较及早期选择
由于单株材积是胸径和树高累积效应的结果,且单株材积与胸径的树高的相关性大于胸径和树高,因此以单株材积为主树高和胸径为辅进行优良家系早期选择。将34个香椿半同胞家系分为Ⅰ、Ⅱ、Ⅲ、Ⅳ 4类:遗传增益Ⅰ>Ⅱ>Ⅲ>Ⅳ,定为优、良、中、差4等,结果见表7,不同等级包含的香椿家系见表8。
由上表可见,Ⅰ等家系包含了蓬溪22、25号和洪雅9号3个家系,入选率8.82%,都是来自四川川中丘陵区的家系,胸径、树高、单株材积平均值分别为6.10 cm、5.23 m和0.0087 m3,胸径、树高、单株材积的选择差分别为1.29 cm、0.93 m和0.0037 m3;胸径、树高、单株材积的遗传增益分别为22.19%、18.30%、58.28%,为优等家系。Ⅱ等家系4个,入选率11.76%,除了灌阳2号来自广西,沐川7号、筠连56、57号均来自四川川南地区,平均胸径、树高、单株材积分别为5.65 cm、5.05 m和0.0072 m3,胸径、树高、单株材积的选择差分别为0.84 cm、0.75 m和0.0022 m3;胸径、树高、单株材积的遗传增益分别为14.45%、14.77%、34.96%,胸径和树高均有一定的增益,单株材积的遗传增益较大,为良等家系,有一定的生长潜力,有待继续观测。Ⅲ等家系包括12个家系,胸径、树高、单株材积的选择差均为正值,胸径和树高的遗传增益均小于10%,单株材积的遗传增益平均为15.05%,为中等家系。Ⅳ等家系15个,胸径、树高、单株材积的选择差和遗传增益均为负值,长势差,平均胸径、树高和单株材积均低于34个家系的总平均值,为差等家系。

表6 香椿不同家系单株材积生长变异Tab.6 Variation of individual volume growth of different Toona sinensis families
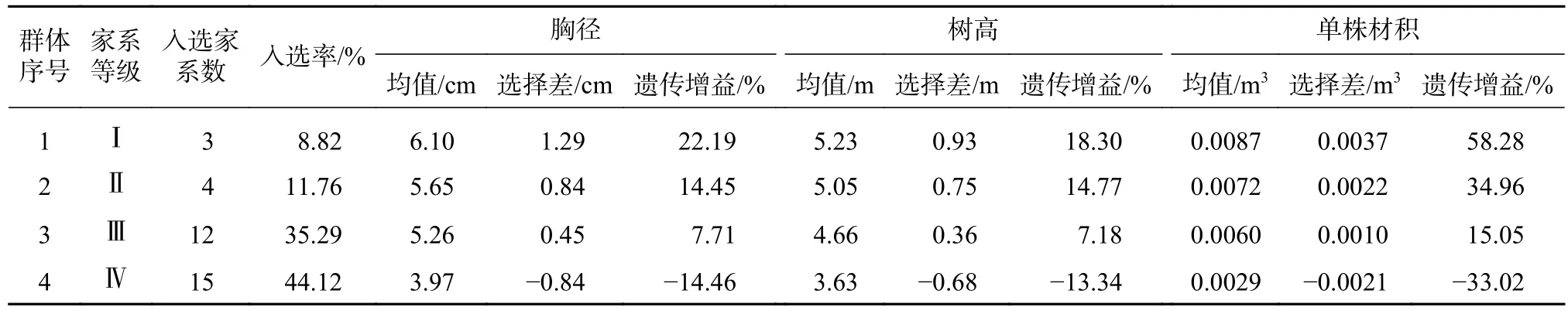
表7 香椿优良家系早期选择与遗传增益估算Tab.7 Early selection and genetic gain estimation of Toona sinensis superior families

表8 不同等级包含家系Tab.8 Toona sinensis families that belonging to different grades
3 结论与讨论
香椿半同胞家系的生长与优树本身的遗传因子有关,同时也与优树种源所在地的环境因子有关。胸径、树高和单株材积与纬度和海拔呈极显著的负相关性,与孙鸿有等[16]的研究结果一致,与年均气温、降雨量和无霜期呈正相关性,地理位置影响大小为纬度>海拔>经度,气候影响大小为无霜期>年均气温>降雨量。10个香椿种源中总体上四川的种源好于广西和福建的,陕西的种源生长最差,与相关性分析结果一致。因此在进行良种选育时,应该在比试验区域所在位置纬度低、海拔低、经度小以及无霜期长、年均气温高、降雨量大的种源中进行优树选择,收集其种子育苗开展良种选育,更有利于选育出适合该区域的香椿良种。
变异系数的大小决定了林木生长的均匀程度。34个香椿半同胞家系中,胸径22个家系的变异系数小于15%,树高有23家系的变异系数大于15%,单株材积的变异系数均大于15%,变异系数总体比苗期大[10-11],表明香椿半同胞家系随着树龄的增长,家系内逐渐出现一定的分化,但绝大部分家系的胸径和树高还是生长较为一致,有利于开展家系良种选育。
34个香椿半同胞家系3年生平均胸径、树高和单株材积分别为4.81 cm、4.30 m和0.0050 m3,变幅分别在2.43 cm~8.30 cm之间、1.80 m~6.40 m之间、0.0010 m3~0.0179 m3之间,不同家系间均有极显著差异,与大部分香椿半同胞家系早期选择的研究结果一致[2-8],具有很好的早期选择基础。34个香椿半同胞家系中,胸径生长以蓬溪22号最大,其次是洪雅9号,平均胸径分别为6.35 cm和6.06 cm,年均胸径>2.02 cm,遗传增益分别为26.47%和21.51%;树高以洪雅9号最大,平均树高5.33 m,年均树高生长量1.78 m,遗传增益为20.26%,其次是蓬溪22号、筠连56号、霞浦7号、灌阳2号、蓬溪25号5个家系,年均树高生长量在1.7 m以上,遗传增益在16.06%以上;单株材积排在前3位的依次是蓬溪22号、蓬溪25号和洪雅9号,遗传增益分别为67.29%、55.93%和51.62%。
遗传力反映亲代将把性状遗传给子代的能力[17],本文34个家系的胸径、树高和单株材的遗传力分别为0.830、0.850和0.777,树高的遗传力略大于胸径和单株材积,考虑到单株材积是胸径和树高累积效应的结果,因此以单株材积为主,胸径和树高为辅进行优良家系早期选择。为了提高选择效果,降低了入选率,提高了选择差[18],选择出的蓬溪22、25号和洪雅9号3个家系,入选率8.82%,胸径、树高、单株材积的选择差分别为1.29 cm、0.93 m和0.0037 m3,遗传增益分别为22.19%、18.30%、58.28%,入选率很低,选择差较大,胸径、树高、单株材积的增益均较大,选择效果较好。本次选择的3个香椿早期优良家系,有很大可能成为良种的潜力,对半个轮伐期后香椿良种选育提供了重要的参考价值。
猜你喜欢
中华实用诊断与治疗杂志(2022年1期)2022-08-31
中华实用诊断与治疗杂志(2022年1期)2022-08-31
广西林业科学(2022年3期)2022-07-08
东方少年·布老虎画刊(2020年3期)2020-06-09
中南林业科技大学学报(2019年4期)2019-04-08
家教世界·创新阅读(2018年9期)2018-12-17
森林工程(2018年1期)2018-05-14
绿色科技(2016年23期)2017-03-15
卷宗(2012年5期)2012-10-21
中国火炬(2012年9期)2012-07-25
