
镉胁迫对双腺藤幼苗生长及生理特性的影响*
2021-06-24 13:49肖雪李宗艳马长乐于达勇
西部林业科学 2021年3期
肖雪,李宗艳,马长乐,于达勇
(云南省功能性花卉资源及产业化技术工程研究中心,西南林业大学 园林园艺学院,云南 昆明 650224)
近年来,随着矿产开发、工业污染、交通排放以及农业生产等活动的持续进行,我国土壤重金属污染愈加严重[1-2]。据统计,我国土壤重金属污染区域主要分布在中国的中部、南部和西南部等地区,其中镉污染率最高[3]。重金属镉毒性强,易扩散,难降解,对人类和生态系统健康具有极大威胁[4]。近年来,媒体曝光了许多“镉麦”“镉米”事件,反映了我国重金属污染的严重性[5]。如何有效防止土壤环境重金属污染,如何修复土壤尤其是耕地土壤重金属污染以保障粮食作物安全生产,已成为当前土壤及环境领域的研究热点和难点。由于土壤中的重金属易被植物吸收与积累,因此,植物修复可为传统的物理或化学修复技术提供经济有效和可持续的替代方案[6],而将园林植物栽植于重金属污染地区,将会起到景观美化和土壤修复的双重作用。
双腺藤(Mandevillasanderi)为夹竹桃科(Apocynaceae)双腺藤属(MandevillaLindl.)多年生常绿藤本植物,原产美洲热带,有着“热带藤本皇后”的美称。该植物花大色艳、花叶繁盛、喜光耐热、对土壤的适应性强,在竖向立面景观及园林工程地栽中表现优秀,其蔓生性不强,亦适合室内盆栽,具有较高的观赏价值[7-9]。目前,对双腺藤的研究主要集中在栽培[10-12]、组培[13-15]、以及病害[16]等方面,重金属胁迫下双腺藤的适应性及耐受机制未见报道。本试验通过对双腺藤幼苗进行不同浓度(0、5、20、50、100、200 mg/kg)镉处理,研究双腺藤在镉胁迫下的一系列生长变化以及生理响应,为进一步了解植物对镉胁迫的生长生理响应机制提供理论依据,也为镉污染地区选择新型园林观花植物提供参考。
1 材料和方法
1.1 试验材料
供试材料为2020年7月中旬从上海怡源公司购买的长势一致、苗龄约1个月的双腺藤扦插苗,共100盆。换盆于带有托盘的圆形塑料花盆(15 cm×12 cm)中,每盆500 g风干基质,1盆栽植1株,于西南林业大学树木园大棚中进行统一的养分和水分管理,保持其正常的生长发育和水分代谢。供试基质由腐殖土、自然土与珍珠岩按照4∶1∶1(体积比)的比例配成,pH值为5.6,镉背景值是0.53 mg/kg。
1.2 试验设计
在统一的养分和水分管理下培养30 d后,选择长势良好且表型一致的植株,开展土壤镉胁迫试验。试验采用盆栽方式,设置5个不同浓度镉处理:Cd1(5 mg/kg)、Cd2(20 mg/kg)、Cd3(50 mg/kg)、Cd4(100 mg/kg )、Cd5(200 mg/kg)。将CdCl2·2.5H2O水溶液均匀浇灌在需要处理的盆土中,对照(CK)浇灌同体积的清水,每个处理12盆,共72盆。试验期间的浇水以见干见湿为准则,并进行正常的田间管理。镉胁迫处理2个月后进行各项指标的测定,每个处理重复3次。
1.3 测定方法
1.3.1 生长指标测定
采集双腺藤完整植株,用卷尺(精确到0.1 cm)和游标卡尺(精确到0.01 cm)分别测量株高、基径、根长,并计算单株总叶片数和分枝数;将双腺藤植株洗净擦干,用剪刀将其剪成根、茎、叶3部分,分别放入105 ℃烘箱杀青30 min,再将烘箱调至70 ℃烘干至质量不变,取出后准确称量植株的各部分生物量。
1.3.2 生理指标测定
采用浸泡法测定叶片相对电导率,将用蒸馏水洗净并擦干的叶片剪成大约1 cm长的长条,快速称取鲜样3份,每份0.1 g,分别置于10 mL蒸馏水的试管中,室温下浸泡12 h,用电导仪测定电导率[17];采用硫代巴比妥酸(TBA)法测定丙二醛(MDA)含量,用紫外分光光度计于450、532、600 nm处测定吸光值;可溶性蛋白含量用考马斯亮蓝染色法测定,5 mL考马斯亮蓝G-520加入酶提取液1 mL,混匀后放置5 min,在595 nm下比色;游离脯氨酸含量采用酸性茚三酮法测定,吸取上层脯氨酸红色甲苯溶液用紫外分光光度计于520 nm波长处测定吸光值[18]。
称取0.2 g待测叶片于5 mL的0.05 mol/L磷酸缓冲液(pH=7.8,内含1%的PVP和1 mmol/L的EDTA-Na2)和少量石英砂中冰浴研磨至匀浆,4 ℃下11 000 r/min离心30 min,最后定容至5 mL,上清液即为所需酶液。用氮蓝四唑法测定超氧化物歧化酶(SOD)活性,愈创木酚法测定过氧化物酶(POD)活性,紫外分光光度计法测过氧化氢酶(CAT)活性[19]。
1.4 数据处理
采用 Microsoft Excel 2010 统计处理数据、计算平均值和标准差;结合SPSS 22.0软件对各指标进行单因素方差分析(One-way ANOVA),并用Duncan法进行多重比较,显著性水平α=0.05。使用Origin 2018软件制图。
2 结果与分析
2.1 不同浓度镉处理对双腺藤幼苗生长特性的影响
如表1所示,随Cd2+浓度增加,双腺藤幼苗叶片数、分枝数、株高均呈现先增加后降低的趋势,其中:3个指标的值在Cd1处理中均最大,比CK分别增加了0.6%、5.6%、2.1%,但差异均不显著,而在其他处理中它们的值均低于CK,且在Cd4、Cd5处理中它们的值与CK差异均达显著水平(P<0.05);叶片数和分枝数在Cd4、Cd5处理中分别比CK降低了10.7% 、28.3%和27.8%、44.4%(P<0.05),而在Cd2、Cd3处理中则均与CK差异不显著;株高在Cd2处理中与CK差异不显著,而在Cd3、Cd4、Cd5处理中比CK分别降低了8.1%、10.5%、12%,差异显著(P<0.05)。随Cd2+浓度增加,双腺藤的基径逐渐降低(表1),Cd1、Cd2的基径与CK差异不显著,Cd3、Cd4、Cd5的基径分别比CK显著降低12.2%、12.5%、14.6%(P<0.05)。随Cd2+浓度增加,双腺藤的根长呈现出先增后降再增再降的变化规律(表1),Cd1比CK高 1.8%,差异不显著,而其他处理的根长均低于CK,其中:Cd3、Cd5的根长分别比CK显著降低了12%、15.1%(P<0.05),Cd2、Cd4的根长均与CK差异不显著。
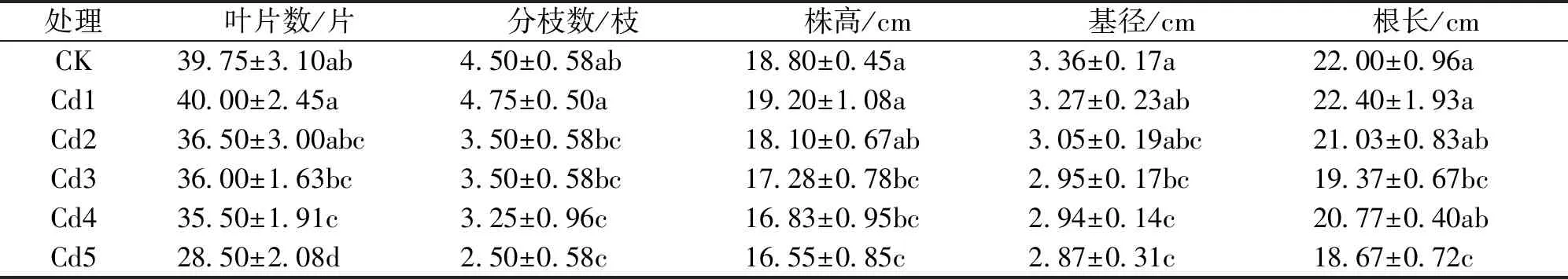
表1 不同浓度Cd2+处理对双腺藤幼苗生长指标的影响Tab.1 Effects of different concentrations of Cd2+ treatments on the growth indices of M.sanderi seedlings
2.2 不同浓度镉处理对双腺藤幼苗生物量的影响
由图1可知,随着土壤Cd2+浓度的递增,双腺藤根生物量呈现先增后降的变化规律,茎生物量逐渐递减,而叶、总生物量呈波浪形递减趋势。除茎生物量外,各生物量均在Cd1时达到峰值,但与CK相比均不显著,其他Cd2+浓度处理下的各生物量均比对照低。Cd2时,根、茎、叶、总生物量与CK相比,均未显著减少。Cd3浓度下的茎生物量比CK显著下降了16.3%(P<0.05),其余生物量与CK相比不显著。Cd4下的根、茎、总生物量均比对照显著下降,但叶生物量的减少与对照相比不显著。双腺藤的各生物量在Cd5时均显著低于对照,比CK降低了41.4%、39.5%、23.5%、29.1%(P<0.05)。不同浓度镉胁迫下,双腺藤的总生物量排列为Cd1>CK>Cd3>Cd2>Cd4>Cd5。
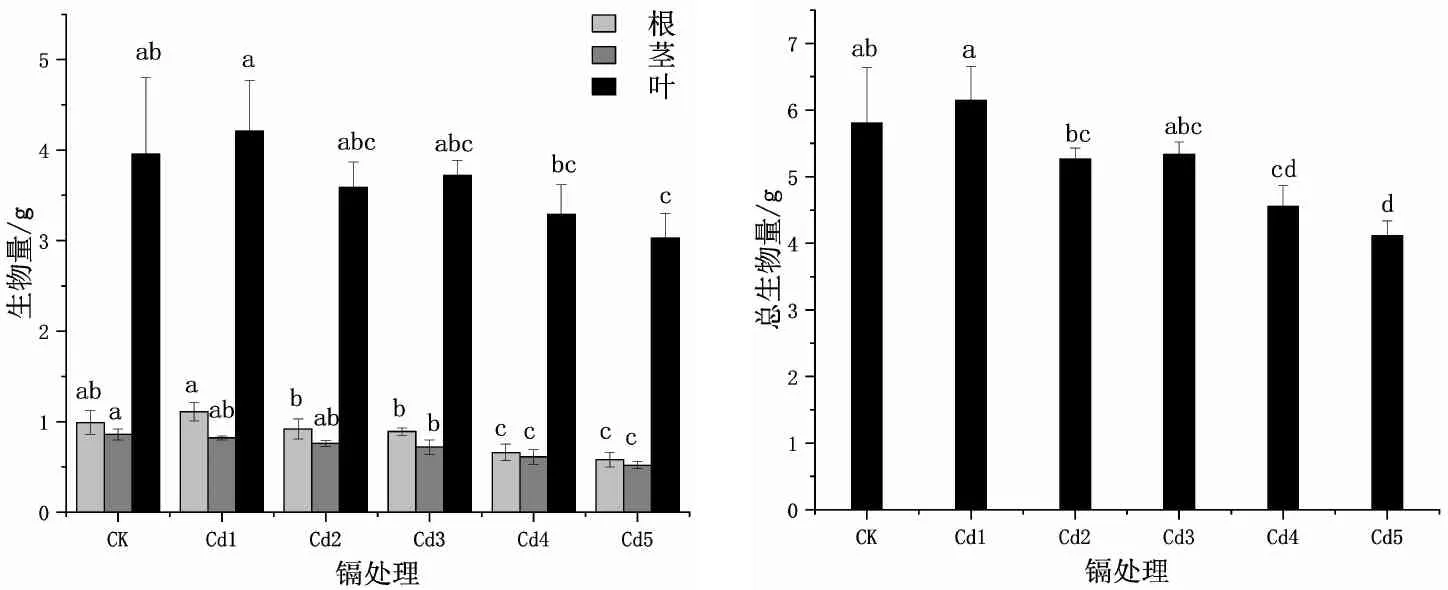
图1 不同浓度Cd2+处理对双腺藤幼苗根、茎、叶及总生物量的影响
2.3 不同浓度镉处理对双腺藤幼苗叶片膜脂过氧化及渗透调节物质的影响
从图2可知,不同浓度的Cd2+处理下,双腺藤的丙二醛含量均高于CK,Cd2+浓度越高,丙二醛含量越高。Cd1比CK增加2.7%,但差异不显著,Cd2、Cd3、Cd4与CK相比呈显著差异(P<0.05),分别增加了40.8%、50.1%、54.7%,Cd5的丙二醛含量最高,比CK增加了1倍多,差异显著。
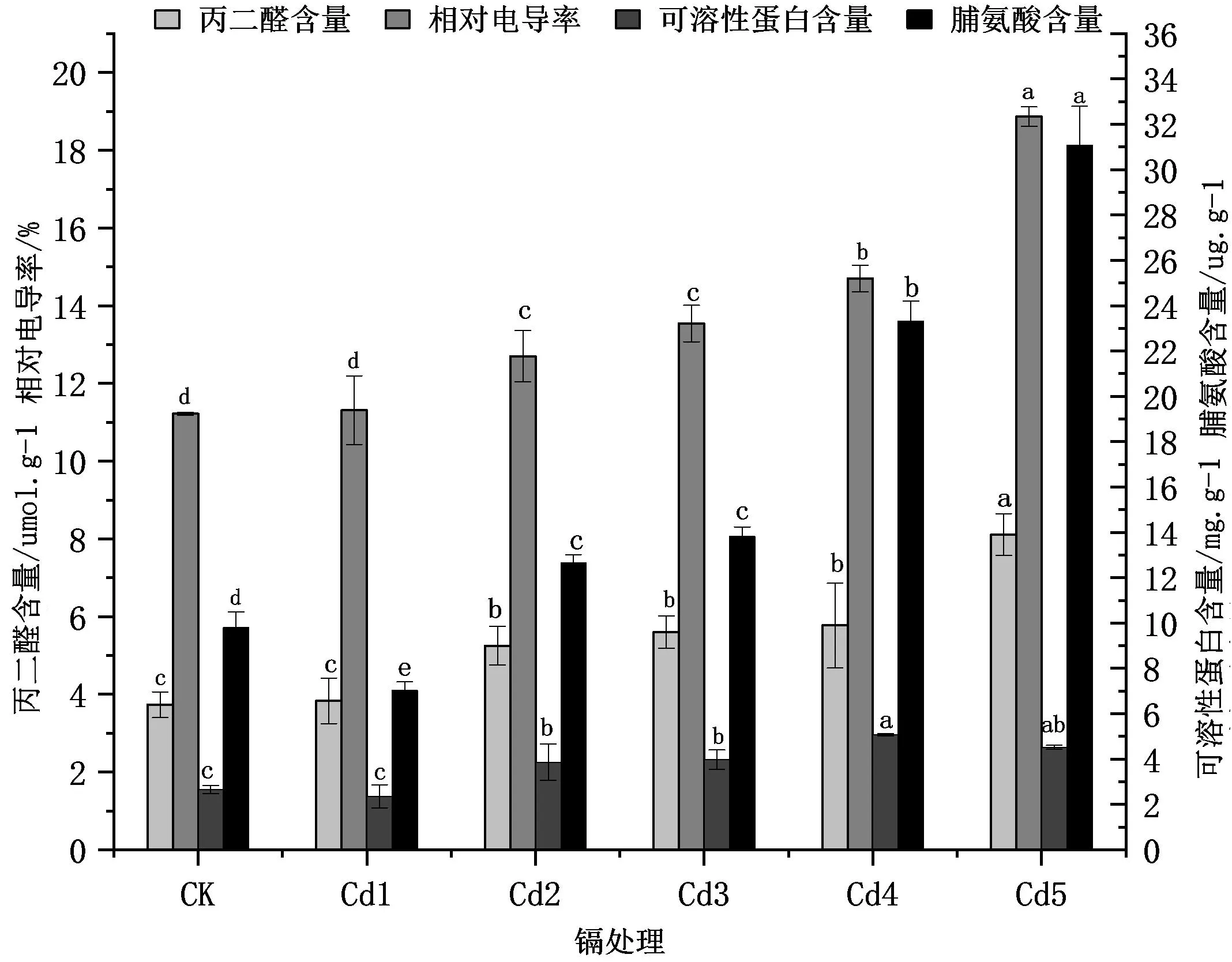
图2 不同浓度Cd2+处理对双腺藤幼苗叶片膜脂过氧化及渗透调节物质的影响Fig.2 Effects of different concentrations of Cd2+ treatments on membrane lipid peroxidation and osmotic regulatory substances in leaves of M.sanderi seedlings
随着Cd2+浓度增加,双腺藤幼苗叶片的相对电导率也随之增大(图2)。当Cd2+浓度较低时,例如Cd1,其相对电导率与CK相比差异不显著,但是当Cd2+浓度大于等于Cd2时,双腺藤的相对电导率相较于CK而言,差异均显著(P<0.05),Cd5的相对电导率最大,比对照增加了68.2%。
双腺藤幼苗叶片可溶性蛋白含量随Cd2+胁迫浓度增加呈现先降低后升高再降低的趋势,而脯氨酸含量则随Cd2+胁迫浓度增加呈现先降低后升高的趋势(图2)。Cd1处理下的可溶性蛋白含量与脯氨酸含量均比对照低,但其脯氨酸含量与对照相比差异显著,降低了28.4%(P<0.05),而其可溶性蛋白含量相较对照差异不显著。双腺藤幼苗叶片可溶性蛋白含量与脯氨酸含量分别在Cd4、Cd5时达到最大,分别为CK的1.9倍和3.2倍。
2.4 不同浓度镉处理对双腺藤幼苗叶片抗氧化酶活性的影响
由图3可见,随着Cd2+胁迫的增加,双腺藤幼苗叶片中的SOD、POD、CAT活性总体表现为先增后降的趋势,但各自变化趋势又有所不同。其中:叶片SOD活性在Cd1处理时达到最大值,相比于CK差异不显著,只增加了0.8%,之后,随Cd2+浓度增加,Cd2~Cd5处理的SOD活性逐渐下降,且均显著低于CK(P<0.05),尤其在Cd4和Cd5时较对照分别下降了近1/3和1/2;POD活性在Cd4时达到峰值,比CK显著高3.4倍(P<0.05),除Cd1的POD活性不显著高于CK外,其他浓度的处理均显著高于CK;Cd3、Cd4、Cd5的CAT活性均比CK显著增加,其中Cd4的CAT活性最高,比CK增加了约1.4倍(P<0.05),Cd1、Cd2的CAT活性也比CK高,但差异不显著。
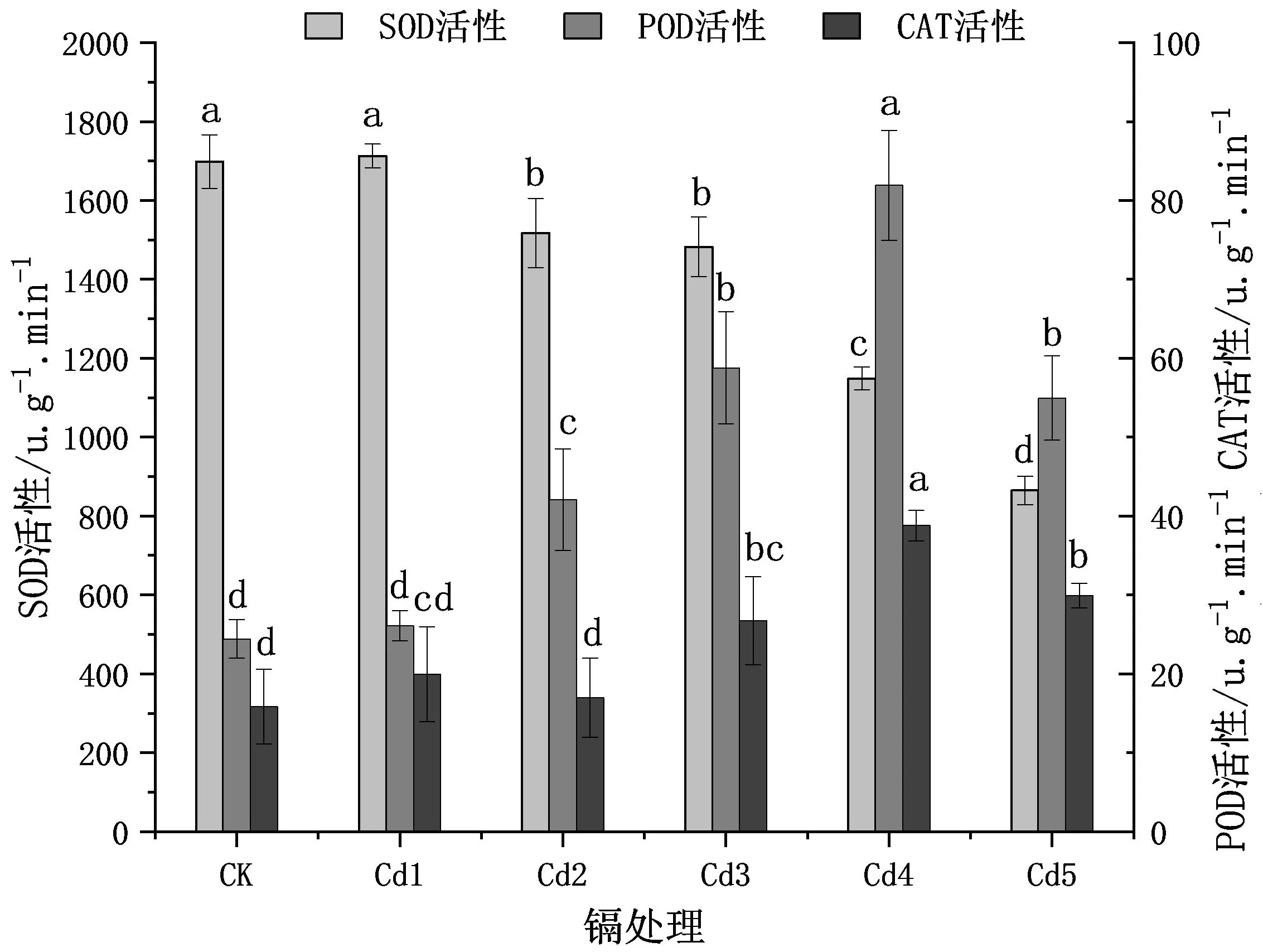
图3 不同浓度Cd2+处理对双腺藤幼苗SOD、POD、CAT活性的影响Fig.3 Effects of different concentrations of Cd2+ treatments on SOD,POD and CAT activity of M.sanderi seedlings
3 讨论与结论
镉胁迫通常会使植物的生长量、生物量以及膜脂过氧化和抗氧化酶系统受到影响。植株的生长指标和生物量等是较易观察的指标。研究表明,镉胁迫对某些植物具有“低促高抑”的现象[20-21]。本试验中,在低浓度(5 mg/kg)Cd2+胁迫下,双腺藤幼苗未受到明显影响,株高、根长、叶片数、分枝数和生物量相较对照略有增加,在高浓度(≥100 mg/kg)Cd2+胁迫下,双腺藤幼苗各生物量和生长指标都显著下降,说明低浓度Cd2+能促进双腺藤生长,而高浓度的Cd2+则对双腺藤的生长和生物量累积有抑制作用。吴朝波等[22]的研究中,海雀稗(Paspalumvaginatum)的耐性指数、生物量、根冠比在低浓度镉胁迫下与对照无明显差异,且镉浓度为1 mg/kg时地下部生物量较对照增加5.3%,高浓度镉胁迫下海雀稗生物量显著下降,这与本研究结果一致。刘柿良等[23]也通过研究发现,较高浓度镉处理(≥25 mg/kg)明显抑制了长春花(Catharanthusroseus)的生长,而较低浓度镉(≤10 mg/kg)下叶较长、叶宽较大、叶数较多。这几种植物都显示了对镉胁迫具有“低促高抑”的现象,但不同植物之间对相同浓度镉的响应又存在差异,这说明不同植物对同一浓度镉的耐受性不同,对镉胁迫的抗性也不同。本研究表明,双腺藤对低浓度镉污染具有较强的耐性。
植物受到胁迫时,细胞会产生大量的自由基,最终自由基的积累将加剧膜脂过氧化作用,而丙二醛就是膜脂过氧化最重要的产物之一,MDA含量的高低反映了膜脂过氧化的程度,间接反映膜系统受损程度以及植物的抗逆性强弱。杨塍希等[24]发现,随着Cd2+浓度的升高,火炬树(Rhustyphina)幼苗叶片中MDA含量呈逐渐上升趋势,其结果与本研究一致:随着Cd2+浓度增加,双腺藤叶片中MDA含量逐渐升高,显示着Cd2+胁迫对双腺藤植物膜系统有一定的损坏。逆境胁迫下植物叶片细胞的膜系统受到伤害后会导致细胞膜透性增大,内容物外渗,电解质含量增加,进而导致相对电导率增大[18]。贾茵等[25]的研究指出,小报春(Primulaforbesii)的相对电导率总体上随着镉浓度的增加不断增大,在低浓度(5 mg/kg)的镉处理下其相对电导率比对照略低,但差异不显著,此结果与本研究基本一致。双腺藤的相对电导率随着镉浓度的增加显示出不断增大的趋势,虽然在低浓度(5 mg/kg)的镉处理下相对电导率比对照略高,但差异亦是不显著,可能是低浓度的镉虽然对植物影响不大,但也可能会对植物体造成短时间的代谢紊乱。
植物在重金属胁迫下,可溶性蛋白含量增加,细胞渗透势增大,同时与重金属离子形成重金属结合蛋白,从而减轻对植物的毒害,增强植物抗性[26]。在逆境胁迫下,植物体内的游离脯氨酸含量会增加,脯氨酸是一种细胞质渗透调节物质,能够稳定生物大分子结构,游离脯氨酸积累可防止植物细胞结构和功能受损伤,使其伤害程度降低,此外,游离脯氨酸还可以清除植物体内的活性氧自由基[27]。张家洋等[28]研究报道,在两叶一心期,小麦(Ttiticumaestivum)和稗草(Beckmanniasyzigachne)游离脯氨酸含量均随处理浓度的增加而增加。程守梦等[29]的研究中,洋常春藤(Hederahelix)的可溶性蛋白随着镉浓度的增加不断增加。本试验中,可溶性蛋白、脯氨酸含量在低Cd2+处理下增加不明显,在高Cd2+胁迫下脯氨酸含量显著升高,而可溶性蛋白含量先增加,最后在Cd2+浓度为200 mg/kg胁迫下降低了。说明低Cd2+胁迫对双腺藤叶片影响不大,而高Cd2+胁迫则引起植物体内的可溶性蛋白、游离脯氨酸等渗透调节物质积极响应,增大细胞的渗透势,从而增强植物对镉的抗性。
在逆境条件下,植物细胞内将产生大量的自由基,体内的自由基平衡被破坏。植物体内的超氧化物歧化酶(SOD)、过氧化物酶(POD)以及过氧化氢酶(CAT)等抗氧化酶在参与活性氧代谢中扮演着重要的角色,由SOD催化分解自由基转化成为的H2O2被POD和CAT快速清除,以此来维持植物体内自由基的平衡[27]。在本研究中,双腺藤幼苗叶片中的SOD、POD、CAT活性随土壤中镉含量的增加总体上呈先升后降的变化规律,但各个酶的具体变化又不尽相同。双腺藤叶片中SOD活性最大值出现在镉浓度为5 mg/kg时,此时SOD 通过分解超氧自由基获得了大量H2O2,在此之后,为了减轻毒害,双腺藤植株体内的POD和CAT活性被激活;但是当镉浓度增大到一定值时,由于酶合成中断以及大量的超氧自由基积累导致酶亚基压力聚集变化,从而导致SOD、POD、CAT活性受到抑制,双腺藤的正常生理代谢受到影响。本试验中的SOD、CAT活性变化与贾茵等[25]的研究结果基本一致,但POD活性变化有所不同,表明不同植物对镉胁迫有不同的耐性以及不同的抗氧化酶响应机制。
在本试验中,不同浓度Cd2+胁迫下,双腺藤幼苗的生长及生理指标均受到不同程度影响,各项指标随Cd2+浓度的升高总体上呈先升高后降低的趋势。在低中浓度Cd2+胁迫下双腺藤幼苗表现出一定的适应性,低浓度(5 mg/kg)Cd2+胁迫对其生长甚至会有轻微促进作用,使其总生物量升高,而高浓度Cd2+胁迫下,双腺藤幼苗的叶片数、分枝数、株高、基径、根长以及生物量均会出现不同程度的减少。这表明植物在不同浓度的Cd2+胁迫环境下会做出相应的生长及生理响应,以维持其生存。随Cd2+胁迫浓度增加,双腺藤幼苗叶片的相对电导率以及丙二醛、可溶性蛋白质、脯氨酸含量持续增加,抗氧化酶活性呈先升高后下降的变化趋势。综上所述,苗龄2个月左右的双腺藤受到≤200 mg/kg的土壤镉污染后均能保持生存状态,表明双腺藤幼苗具有一定的耐镉能力,能够通过调节体内的渗透调节物质以及抗氧化酶活性等来抵抗土壤中的镉胁迫,在土壤镉污染修复中具有一定的应用潜力,可作为一种新型园林观花植物应用于土壤镉污染地区的园林绿化中。
猜你喜欢
中老年保健(2022年3期)2022-11-21
今日农业(2021年10期)2021-11-27
今日农业(2020年19期)2020-12-14
科学与财富(2019年15期)2019-10-21
湖南林业科技(2017年1期)2017-02-06
中国科技信息(2015年2期)2015-11-16
中国火炬(2015年12期)2015-07-31
植物营养与肥料学报(2014年1期)2014-03-11
中国火炬(2013年3期)2013-07-24
中国蜂业(2013年18期)2013-01-17
