
调亏灌溉对棉花叶片和根系保护酶活性的影响
2021-06-24 04:27:28孙路张世民罗新宁
塔里木大学学报 2021年2期
孙路,张世民,罗新宁*
(1塔里木大学植物科学学院,新疆 阿拉尔 843300)
(2新疆生产建设兵团种子管理总站,新疆 乌鲁木齐 830011)
由于气候变化,极端天气的频繁发生已经引起了科学家们的广泛关注[1]。降水量的减少使得干旱发生的频率不断上升,这限制了植物生长并降低世界范围内作物产量和生产能力[2-3]。干旱的环境条件极大影响了作物的生长和发育[4-5],在正常条件下植物体内活性氧含量处于动态平衡,然而在各种胁迫条件下活性氧逐渐积累使脂膜过氧化,最终使细胞死亡。为了应对各种逆境,植物在长期的进化过程中,产生抗氧化酶系统[6]。抵御活性氧的氧化反应能力取决于细胞的抗氧化酶,其中主要的活性氧清除酶包括超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)[7-8],它们能有效清除细胞内积累的活性氧,减缓对细胞膜造成的伤害,为植物生长发育创造稳定的内环境。丙二醛(MDA)是细胞膜脂过氧化的产物,研究者大多将其含量高低作为植物受逆境胁迫程度的指标[9]。许多科学家在玉米[10]、小麦[11]、水稻[12]等作物中发现抗氧化酶活和丙二醛含量与植物抗逆性密切相关。
棉花作为重要的纺织原料,在全球经济中占有重要地位[13],在我国主要分布在长江流域、黄河流域和西北内陆。新疆作为我国的产棉大省,光照资源丰富,降雨量稀少,棉花在生育期极易遭受干旱[14]。调亏灌溉是利用棉花的抗旱机制对棉花进行水分管理,该方式可大幅度减少棉花灌水量,且保持产量不受影响,因此该技术越来越受到重视。棉花叶片是重要的光合器官,90%以上的干物质积累均来自于叶片,后期叶片早衰对棉花产量有明显的负面影响[15-16]。BAUER P J等[17]和 PENG S等[18]发现叶片衰老会抑制光合作用和棉花纤维发育。此外,根系不仅参与矿质营养和水分的吸收,还参与激素和氨基酸合成与植物固定[19],而且它的生理特性与植物抗旱性密切相关,在干旱发生时反应最为敏感[20]。部分研究表明,棉花早衰也有可能是后期根系从表层土壤吸收养分能力较弱造成的。大量的研究发现并验证了棉花叶片的抗氧化酶系统在逆境中的作用,但当土壤干旱时,作物根系首先受到胁迫[21]。因此,只研究干旱对叶片抗氧化酶活性的影响并不能明确作物抗旱机制,需要同时研究叶片和根系两种器官的抗氧化酶活性对干旱的响应。本试验研究棉花叶片和根系保护酶在调亏灌溉条件下的变化趋势,以期揭示棉花的抗旱机制,为干旱和半干旱地区棉花抗旱研究提供理论基础。
1 材料与方法
1.1 试验地概况
试验于2018年在中国农业科学院棉花研究所阿拉尔试验站(40°36'N,81°18'E)进行,属暖温带极端大陆性干旱荒漠气候,光照充足、热量丰富,年均气温10.7℃,≥10℃积温4 113℃,无霜期220 d,年日照2 900余小时,年均降水量为40.1~82.5 mm,年均蒸发量1 876.6~2 558.9 mm。
1.2 供试材料与试验设计
本试验采用两因素随机区组设计,供试材料为CCRI-60和CCRI-92两个品种。灌水处理为:CK(4 500 m³/hm²)、W1(3 000 m³/hm²)和 W2(1 500 m³/hm²),其中W1和W2为调亏灌溉,6个处理,重复3次,共计18个小区,根据小区面积用流量计精确控制灌水量。4月16日播种,6月9日开始灌溉,每7 d左右灌一次水,7月6日打顶,10月1日收获。
1.3 测定指标及方法
1.3.1 取样
叶片取样:生育期内每隔10~20 d左右采集各处理6株棉花倒4叶,放入冰盒带回实验室,液氮速冻而后放入冰箱保存,备用。
根系取样:根系取样参考龚江等[22]方法。以棉株为圆心在距棉株 5cm,将 0~20 cm、20~40 cm、40~60 cm土壤分层取出。装入网兜用自来水冲洗干净,放入冰盒带回实验室,液氮速冻后放入冰箱保存,备用。
1.3.2 测定
丙二醛采用硫代巴比妥法(TBA)测定;过氧化物歧化酶采用试剂盒法测定(南京建成生物工程研究所);过氧化物酶采用愈创木酚法测定[23];过氧化氢酶采用紫外吸收法测定[24]。
1.4 数据处理
采用Microsoft Excel 2003软件对数据进行处理和绘图,采用DPS 9.50统计分析软件对数据进行差异显著性检验(LSD,α=0.05)。
2 结果与分析
2.1 调亏灌溉叶片及根系丙二醛含量的影响
由图1可知,随着干旱时间的延长,MDA含量逐渐增加,说明干旱造成脂膜过氧化。两品种比较发现,CCRI-92的CK、W2处理叶片MDA含量明显高于CCRI-60叶片,而CCRI-92和CCRI-60的W1处理没有明显差异。CCRI-60叶片W1处理比CK高,W2处理虽然在生育前期增加较慢,但在生育后期MDA含量增加较为明显,这可能是叶片衰老加速造成。CCRI-92的CK和W1处理全生育期MDA含量相差较小,而W2的MDA含量随着干旱时间的延长呈先增加而后下降又增加的趋势。
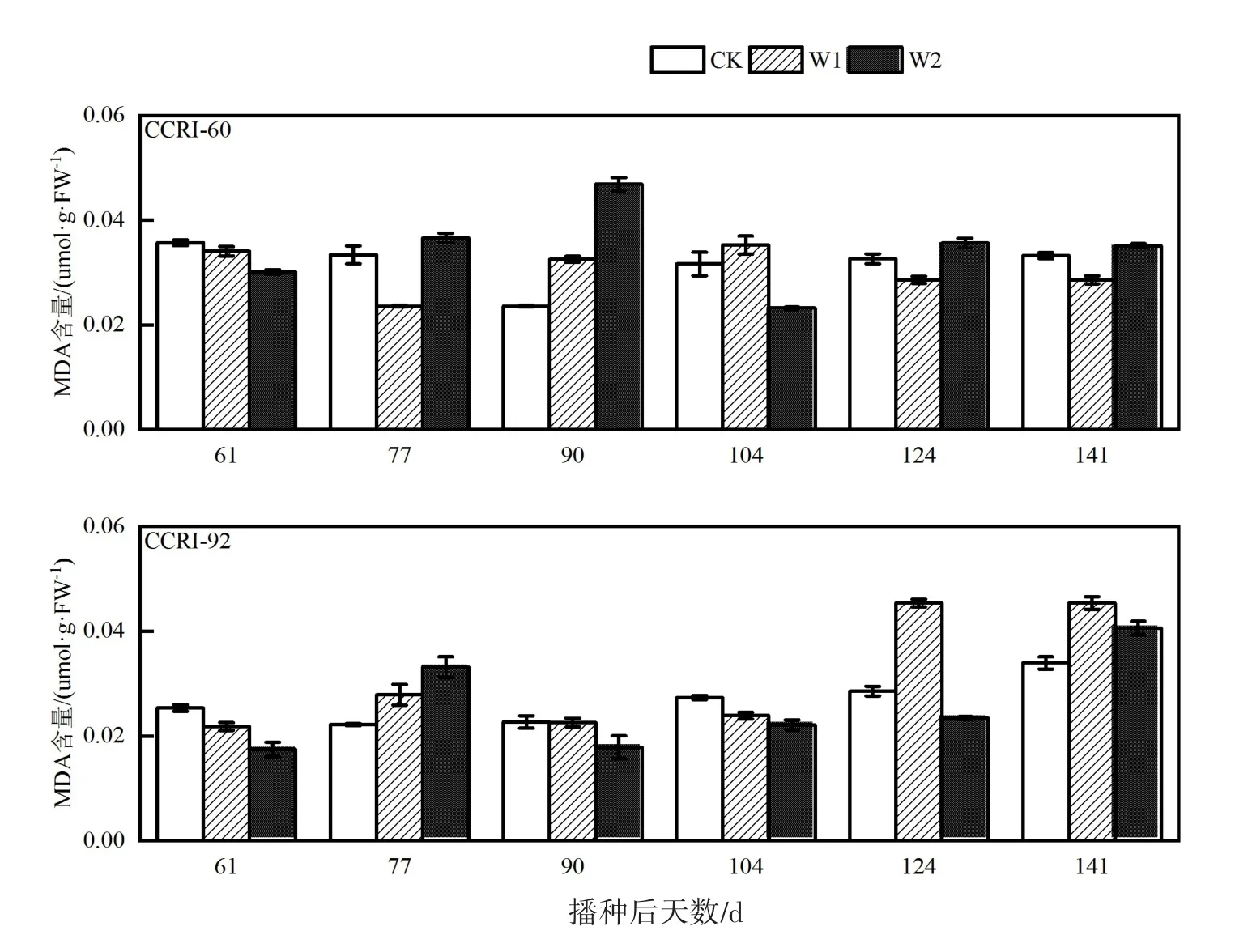
图1 生育期叶片丙二醛含量变化
由图2可知,在0~20 cm和40~60 cm分层取出的土壤中,CCRI-60的根系MDA含量呈现先降低后增加的趋势;20~40 cm呈缓慢下降趋势;CCRI-92的根系MDA含量在0~20 cm CK处理随播种天数的增加呈下降趋势,W1处理变化不大,W2处理呈不断增加趋势;CCRI-92的根系MDA含量在20~40 cm CK处理变化不大,W1处理呈下降趋势,W2处理呈增加趋势;40~60 cm各处理均呈增加趋势。

图2 生育期根系丙二醛含量变化
2.2 调亏灌溉对叶片及根系过氧化物歧化酶活性的影响
由图3可知,生育期CCRI-60叶片SOD酶活性整体上低于CCRI-92叶片SOD酶活性。CCRI-60各处理均呈现生育前期缓慢增加中期下降后期增加的趋势;W1处理、W2处理后期均呈现下降趋势,CK后期有所上升。CCRI-92 CK处理呈生育前期下降中期增加后期下降趋势,W1处理呈先下降后增加的趋势,W2处理呈持续下降趋势。

图3 生育期叶片过氧化物歧化酶活性变化
由图4可知,根系SOD酶活性变化:CCRI-60 0~20 cm CK、W1处理呈先增加后降低的趋势,W2处理呈逐渐增加的趋势;20~40 cm CK、W1处理先增加而后下降趋势,W2处理呈先增加后下降的趋势;40~60 cm CK处理呈先增加后下降再增加的趋势,W1、W2处理先增加后下降的趋势。CCRI-92 0~20 cm CK、W1处理呈先增加后下降的趋势,W2处理呈先下降后增加再下降的趋势;20~40 cm CK处理呈先增加后下降再增加的趋势,W1、W2处理呈先增加后下降的趋势;40~60 cm CK处理呈先增加后下降的趋势,W1处理呈先下降后增加再下降趋势,W2处理呈先增加后下降而后增加再下降的趋势。
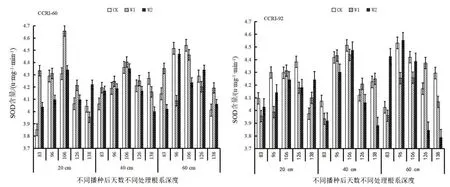
图4 生育期根系过氧化物歧化酶活性变化
2.3 调亏灌溉对叶片及根系过氧化物酶活性的影响
由图5可知,整体上CCRI-60叶片POD酶活性在棉花生长后期明显低于CCRI-92叶片。CCRI-60的CK、W1的叶片POD酶活性在全生育期呈先增加后下降再增加趋势。W2处理呈先下降后增加再持续增加趋势。CCRI-92各处理均呈现前期缓慢增加中期下降后期增加的趋势。CCRI-60与CCRI-92两品种生育期叶片POD酶活性各处理均呈现后期明显高于前期和中期。
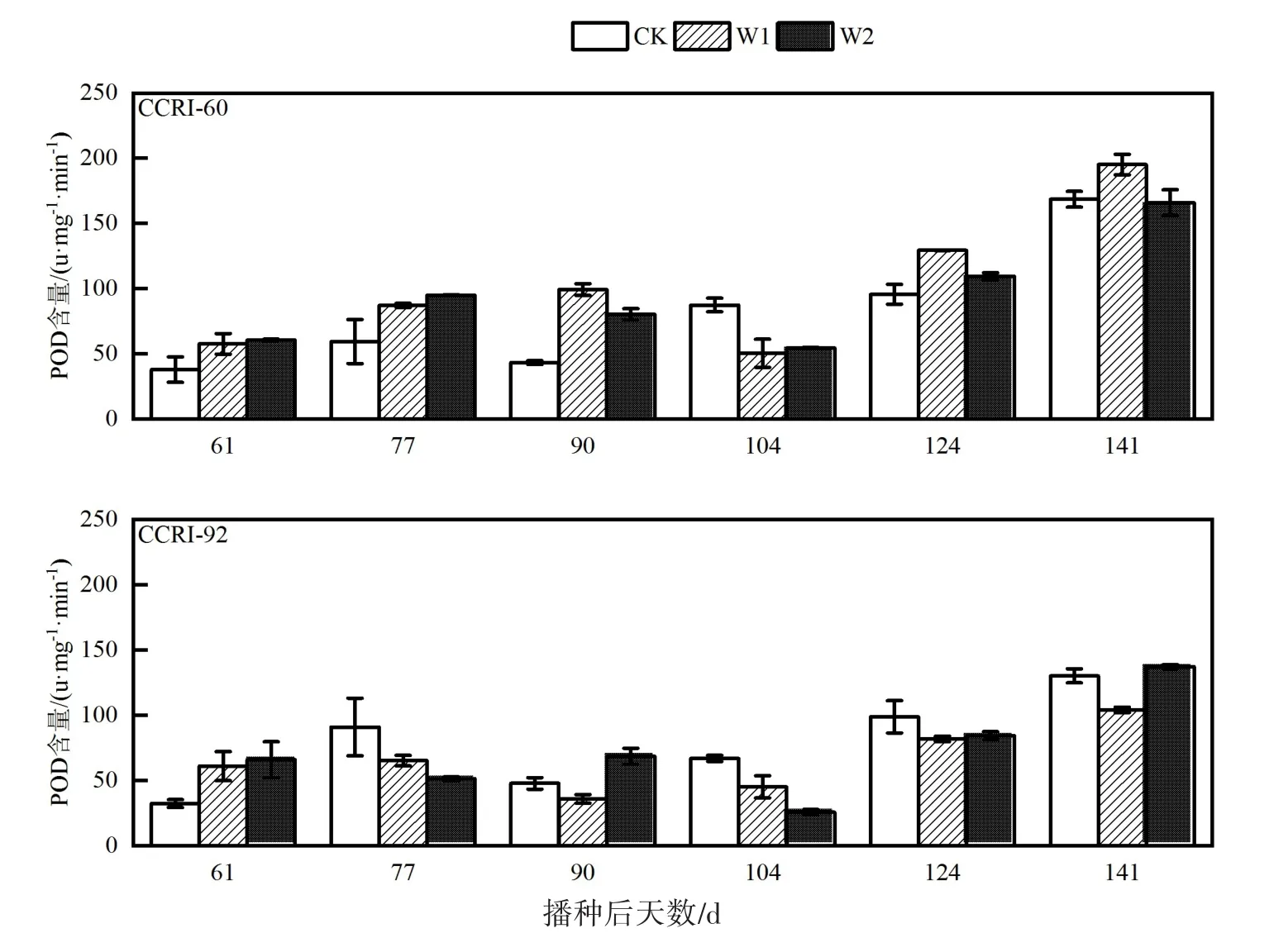
图5 生育期叶片过氧化物酶活性变化
由图6可知,根系POD酶活性变化:CCRI-60 0~20 cm CK、W2处理呈先增加后降低后增加最后降低的趋势,W1处理呈逐渐增加的趋势;20~40 cm CK、W1处理先增加而后降低再上升的趋势,W2处理呈先增加后下降的趋势;40~60 cm CK处理呈先增加后下降再增加的趋势,W1、W2处理先增加后下降的趋势。CCRI-92 0~20 cm CK处理呈先增加后下降的趋势,W1、W2处理呈先上升后下降再增加的趋势;20~40 cm各处理呈逐渐下降的趋势;40~60 cm CK、W1处理呈先增加后下降再增加的趋势,W2处理呈先增加后下降而后增加再下降的趋势。
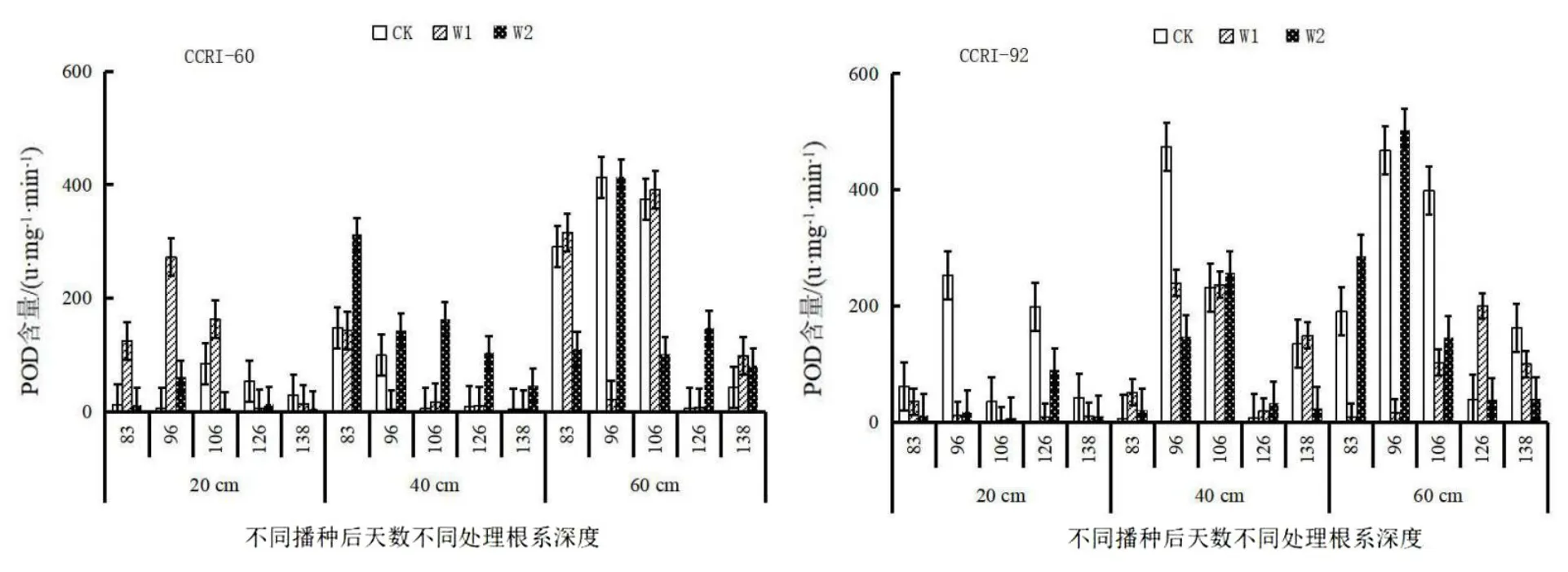
图6 生育期根系过氧化物酶活性变化
2.4 调亏灌溉对叶片及根系过氧化氢酶活性的影响
由图7可知,整体上CCRI-60叶片的CAT酶活性高于CCRI-92(CK处理除外)。CCRI-60处理下,CAT酶活性随灌水量的减少呈逐渐增加趋势;CCRI-92随灌水量的增加呈缓慢下降趋势。CCRI-60 CK处理呈先下降后增加的趋势,W1处理呈先下降后增加最后下降的趋势,W2处理呈先下降后增加再下降最后增加的趋势;CCRI-92 CK处理呈先增加后下降而后增加再下降的趋势,W1处理呈先下降后增加又下降再增加的趋势,W2处理呈先下降后增加的趋势。

图7 生育期叶片过氧化氢酶活性变化
由图8知,根系CAT酶活性变化:CCRI-60 0~20 cm CK、W2处理呈先下降后增加最后降低的趋势,W1处理呈先下降后增加的趋势;20~40 cm CK处理呈先下降后增加最后下降的趋势,W1处理呈先增加而后降低的趋势,W2处理呈先增加后下降而后增加最后下降的趋势;40~60 cm CK处理呈先下降最后增加的趋势,W1处理呈先下降后增加最后降低的趋势,W2处理呈先增加后下降而后增加再下降的趋势。CCRI-92 0~20 cm CK、W1处理呈先增加而后降低的趋势,W2处理呈先下降后增加而后下降再增加的趋势;20~40 cm各处理均呈先增加后下降最后增加的趋势;40~60 cm CK处理呈先下降最后增加的趋势,W1、W2处理呈先下降后增加最后降低的趋势。
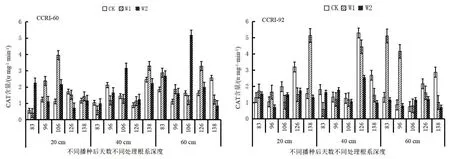
图8 生育期根系过氧化氢酶活性变化
3 讨论
试验研究结果表明,CCRI-60叶片中MDA含量随着生育期的延长逐渐增加,而CCRI-92变化不大,但一直维持较高水平,可认为其脂膜过氧化水平较为严重,由此表明CCRI-60与CCRI-92相比更为耐旱。WANG W B等[25]对于苗期苜蓿在干旱条件下抗氧化酶的研究也得到了相似的结果。SOD作为主要的抗氧化酶,能将超氧阴离子自由基歧化生成H2O2,H2O2能被CAT和POD降解为H2O和O2[26]。对于小麦的调亏灌溉研究认为,小麦抗旱性与SOD酶活性一致[27],本试验结果显示随着生育进程的推进两个棉花品种的叶片SOD酶活性持续降低,而根系SOD酶活性先增加后降低,可能是由于后期根系衰老造成了酶活性的降低。叶片POD酶活性随着生育期的推移持续增加,且灌溉亏缺程度较为严重的W2处理酶活性最高,可能是由于棉花叶片本身保护机制造成的。根系POD酶活性不同层次均表现为生育前期酶活性增加而中后期酶活性降低,且40~60cm根系酶活性最高。CCRI-60随着灌溉亏缺程度的加重叶片CAT酶活性逐渐增加,而CCRI-92则逐渐降低,不同层次差异较大,这可能是品种间的差异造成。
4 结论
棉花被认为是耐旱作物,但产量和品质受干旱影响较大。在西北内陆棉区棉花生产中,灌溉是必不可少的。本试验结果认为CCRI-60和CCRI-92两个棉花品种在干旱条件下都会产生过氧化危害。MDA含量随着生育期的延长和调亏程度的加重而增加,可以作为衡量棉花叶片膜脂过氧化程度的指标。适度的调亏处理会使POD酶活性增加而严重的调亏灌溉处理会使POD酶活性降低。SOD酶两品种间表现不同,发育中后期CCRI-60 CK处理与CCRI-92 CK处理差异明显,在生育末期CCRI-60 SOD酶活性达到最低值,CCRI-92 CK仍然维持较高的SOD酶活性,说明SOD酶活性受品种影响较大。CAT酶两品种根系差异明显,CCRI-60 CAT酶随调亏程度的加重而增加,CCRI-92相反,说明CAT酶受品种影响较大。不同品种MDA含量和SOD、POD、CAT酶活性差异较大,因此需根据品种特性来制定合适的灌量。
猜你喜欢
黑龙江大学自然科学学报(2022年4期)2022-11-17 08:07:52
河北科技师范学院学报(2022年2期)2022-08-26 08:55:46
少儿科学周刊·儿童版(2022年10期)2022-06-27 14:06:58
小读者(2020年4期)2020-06-16 03:33:54
中国塑料(2016年3期)2016-06-15 20:30:00
华东理工大学学报(自然科学版)(2015年4期)2015-12-01 04:00:36
西藏科技(2015年5期)2015-09-26 11:55:25
小天使·一年级语数英综合(2014年7期)2014-06-26 14:42:38
新疆农垦科技(2014年12期)2014-02-28 19:21:53
新疆农垦科技(2014年10期)2014-02-28 19:21:18
