
梁王山大花香水月季居群表型多样性分析
2021-06-22 00:46付荷玲王琛瑶张晓龙
西北植物学报 2021年5期
付荷玲,王琛瑶,张晓龙,罗 乐*
(1 花卉种质创新与分子育种北京市重点实验室,北京 100083;2 国家花卉工程技术研究中心,北京100083;3 城乡生态环境北京实验室,北京100083;4 北京林业大学 园林学院,北京 100083)
表型多样性是遗传因素和环境因素共同作用的结果,是遗传多样性最直接的体现,维持表型多样性对于物种的长期生存至关重要[1-2]。长期以来,利用表型性状的差异来检测物种的遗传多样性是最简便易行的方法[3-4],其研究结果对了解物种居群的遗传变异规律、适应环境变化的能力等具有重要意义,为植物的保护和利用提供了理论支持[5-6]。
大花香水月季(Rosaodoratavar.gigantea)亦称巨花蔷薇,系蔷薇科(Rosaceae)蔷薇属(Rosa)月季组(Sect. Chinenses)植物,主要分布于中国云南,缅甸、泰国、越南、印度阿萨姆邦等地亦有分布[7-8]。但近年来,受人为活动、自然环境变化等的影响,居群和居群内个体数量逐年减少[9]。大花香水月季具有芳香、花大、较耐寒[10]、低温开花、授粉率和结实率高[11]等优良的特性,是中国古老月季和现代月季的重要间接亲本之一[12-13],是重要的种质资源,在月季的起源与育种研究中占据着重要地位。在《中国植物志》中,大花香水月季被记载为香水月季的变种[14],但陈俊愉[12]赞成将其由香水月季变种撤出而独立成种(R.gigantea)。一般认为,大花香水月季与月季花(R.chinensis)杂交产生了香水月季(R.odorata)[12,15],但月季花的原始种单瓣月季花(R.chinensisvar.spontanea)目前主要分布于中国四川,其他省并未发现其野生分布群落[16-17],香水月季可能是人工栽培品种,而非自然杂交的结果[18]。因此,对野外大花香水月季展开野外调查与研究意义重大。
前人研究表明,大花香水月季具有很高的遗传多样性,且在一个相对较小的分布范围内,大花香水月季居群表型性状与环境的相关性不显著[19]。在居群和物种水平上,遗传距离与地理距离显著正相关,地理距离是影响大花香水月季遗传分化的主要因素[9]。因此,本研究以云南省澄江市梁王山一带距离较近、生境相似的3个大花香水月季居群为研究对象,纳入了更多的表型性状,并增加了居群内植株的研究数量,对大花香水月季花部和叶部表型性状进行测定和分析,进一步探讨较近距离内居群间与居群内表型性状的变异规律及分化情况,为大花香水月季的种质资源的收集、保护与开发利用提供理论基础,同时也为研究香水月季的起源与演化、遗传变异和栽培育种等工作提供参考。
1 材料和方法
1.1 居群选择与试验材料采集
对云南梁王山地区展开调查,并依据山谷走向、坡向等环境因子,对大花香水月季居群进行划分。选取了3个生境相似、距离较近、植株分布集中的大花香水月季天然居群进行表型多样性分析(表1,图1),各居群间水平距离仅1~2 km,居群内随机选择80株生长正常、无明显病虫害的植株。选定居群的每个采样植株均用GPS定位。
1.2 表型性状的选取及测定方法
选择具有代表性、遗传相对稳定、易于获得和测定的表型性状,包括花瓣、萼片、花梗、复叶和顶叶等的16个相关性状(图2,表1)。
所有花部性状于4月盛花期野外随机选取单花带回室内用游标卡尺测量,精度0.01 mm。其中,花瓣和萼片性状每朵花任选3枚测量。所有叶部性状均选取当年生枝条中部健壮无病虫害复叶,同期制成标本后用Digimizer软件测量。所有性状每个植株均进行5次重复。
1.3 表型多样性参数及统计分析方法
参照李斌等[20]的方法,采用IBM SPSS Statistics 25软件对各表型性状进行巢氏方差分析,线性模型为:Yijk=μ+Si+T(i)j+e(ij)k;式中Yijk为第i个居群第j个单株第k个观测值;μ为总均值;Si为居群效应(固定);T(i)j为居群内单株效应(随机);e(ij)k为试验误差。


表1 云南梁王山3个大花香水月季居群信息表

P1.新庄村居群;P2.平山居群;P3.饮马池村居群图1 大花香水月季居群生境图P1. Xinzhuang Village population; P2. Ping Mountain population; P3. Yinmachi Village populationFig.1 Habitat of R. odorata var. gigantea populations
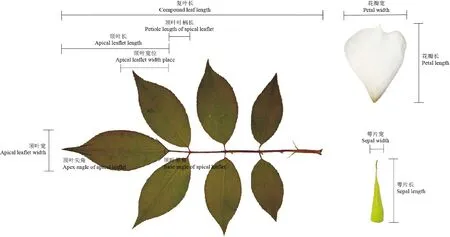
图2 表型性状测量示意图Fig.2 Schematic illustration of measurement of phenotypic traits
为群体间均方,MSs为群体内均方,MSe为随机误差均方,j为群体内样本数,k为观测重复数。
采用Excel 2019软件计算各性状的平均值、标准差和变异系数,表型变异系数CV(%)=σ/μ×100,式中σ为标准差,μ为平均值。采用IBM SPSS Statistics 25软件对表型性状进行Pearson相关性分析。
2 结果与分析
2.1 表型性状在居群间和居群内的差异
对所有表型性状均采用巢氏设计方差分析(表2),结果显示,萼片宽、花梗长、复叶长、顶叶长宽比、顶叶宽位/顶叶长与顶叶叶柄长仅在居群内不同植株间为极显著差异(P<0.01),在居群间无显著差异;其余性状在居群间和居群内不同植株间均具有极显著差异(P<0.01),表明大花香水月季表型性状在居群间和居群内都有较高的多样性 (表2)。
F值的大小表现了结果可靠性的高低,在一定程度上反映了不同表型性状在居群间和居群内差异程度的不同[22]。因此大花香水月季居群间花瓣长变异最为丰富,顶叶长宽比与顶叶叶柄长变异较小;居群内萼片长宽比与花梗长变异较丰富,顶叶尖角变异最小。
2.2 天然居群的表型分化
按巢氏设计方差分量组成比,进一步分析各方差分量占总变异的比例(表3)。16个表型性状在居群间和居群内的平均方差分量百分比分别为4.80%和54.49%,表明3个大花香水月季居群内的分化程度远大于居群间的分化程度。
大花香水月季16个表型分化系数(VST)平均值在-1.51%~26.18%之间,VST最大的性状为花瓣长(26.18%),表明该性状在居群间分化最大;VST最小的为顶叶长宽比(-1.24%)和顶叶叶柄长(-1.51%),萼片宽、花梗长和复叶长、顶叶宽位/顶叶长VST也较低,分别为1.21%、2.14%、2.10%和2.43%,表明这几个性状在居群间几乎无分化。VST总平均值仅为7.65%,即大花香水月季居群表型变异在居群间和居群内的贡献分别为7.65%和92.35%,说明大花香水月季居群内表型多样性程度远大于居群间表型多样性程度。
2.3 大花香水月季表型变异特征
变异系数 (CV) 反映了表型性状的离散程度(表4)。大花香水月季居群表型性状在居群内的平均变异系数为13.62%,变异幅度为6.85%~23.78%,离散程度相对较低。以 16个表型性状的变异系数进行居群间差异性检验,结果显示变异系数在居群间不存在显著差异,说明3个居群的表型性状的离散程度差别不大。以3个居群的变异系数进行不同性状间差异性检验,结果显示变异系数在表型性状间存在极显著差异(P<0.01),这说明表型性状之间的变异程度存在显著差别,体现了不同表型性状对环境适应能力的不同。
平均变异系数最大的是花梗长,为23.78%,表明该性状特征在居群内最不稳定;平均变异系数较小的是花瓣长、花瓣长宽比、萼片宽、顶叶长宽比、顶叶宽位/顶叶长、顶叶基角,表明这几个性状在各居群内变异幅度小,稳定性较高。顶叶长、顶叶宽、顶叶宽位、顶叶叶柄长、顶叶尖角等营养性状在居群内有较高的变异系数,表明这些性状可能具有较高的表型可塑性,更易受到环境的影响。综合来看,生殖性状变异幅度大多低于营养性状,具有更高的稳定性。

表2 大花香水月季居群表型性状的巢氏方差分析

表3 大花香水月季居群表型分化系数分析
P1居群的变异系数最大,结合野外调查来看,可能是因为此居群受到人为活动干扰稍小,遗传多样性水平更高;而P2居群和P3居群的变异系数接近,这两个居群可能遗传多样性水平较为接近。
2.4 大花香水月季的相关性分析
对大花香水月季的16个表型性状进行Pearson相关性分析(表5)。花瓣长与花瓣宽高度正相关,表明花瓣形状比花瓣大小更加稳定;复叶长与顶叶长高度正相关,表明顶叶越长,复叶也相应越长;顶叶宽位与复叶长、顶叶长、顶叶宽高度正相关,表明叶片形状较为稳定。
顶叶长宽比与顶叶尖角、顶叶基角呈中度负相关关系,表明顶叶尖角和顶叶基角越小,叶片形状越狭长。花瓣的长、宽与萼片的长、宽相关系数也较大,表明花瓣的大小与萼片的大小相关性较强。顶叶宽与复叶长、顶叶长中度相关,顶叶叶柄长与复叶长、顶叶长、顶叶宽、顶叶宽位相关系数较高。而花部性状与叶部形状间相关性较低或不相关。
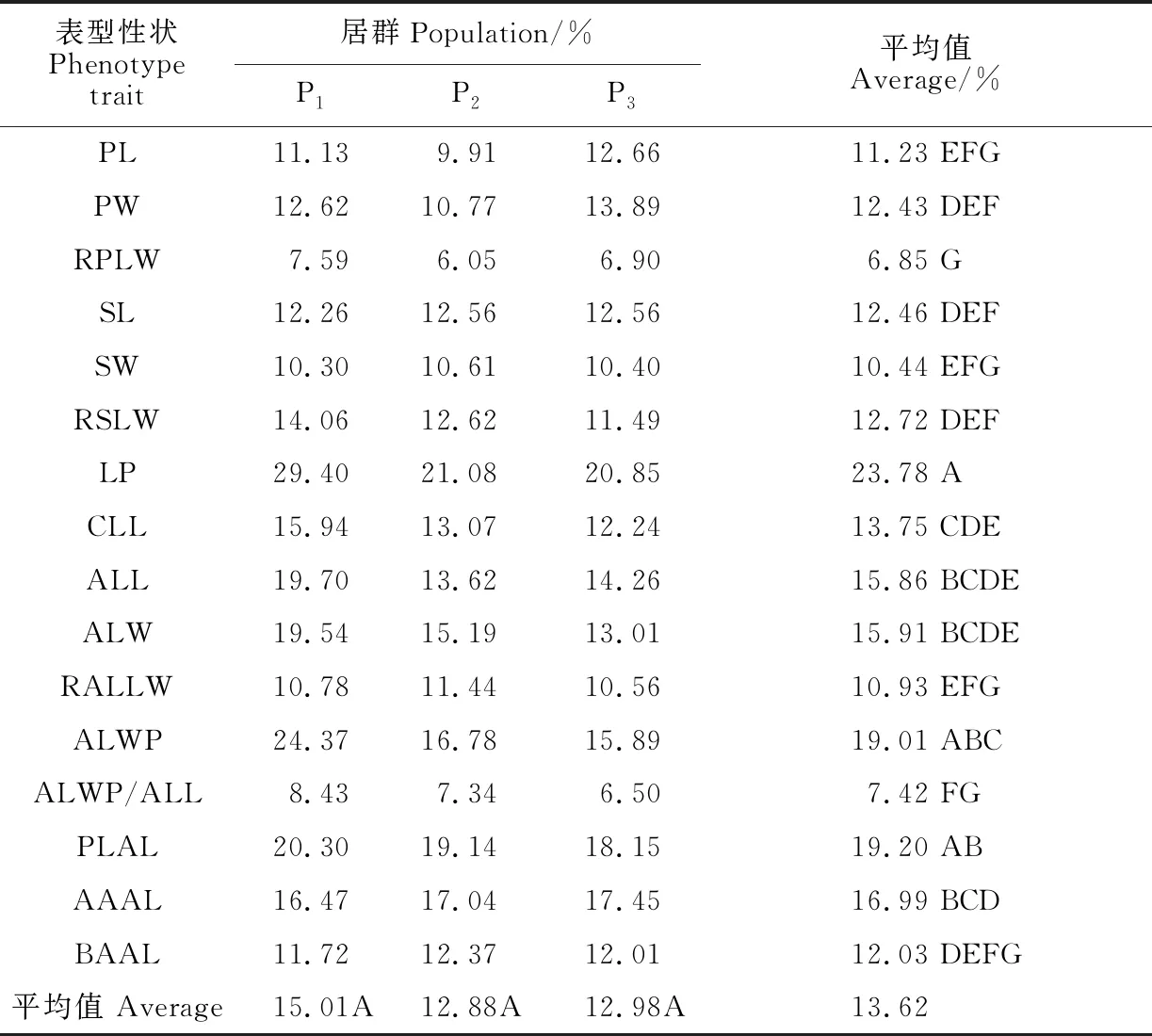
表4 大花香水月季居群表型性状变异系数分析

表5 大花香水月季表型性状间的 Pearson 相关性分析
3 讨 论
3.1 大花香水月季的表型分化情况
居群间多样性是种内多样性的重要组成部分[23]。本研究发现,大花香水月季3个居群表型分化程度较低,居群间变异仅为7.65%,说明居群间多样性远低于居群内多样性。居群间可能存在较强的基因交流,这可能与这3个居群距离较近有关,与其他大花香水月季的研究结论一致[9,19]。大花香水月季为异交昆虫授粉木本植物,较近范围的大花香水月季居群间的基因交流可能更频繁,发生遗传分化的可能性更小[24],这有利于维持大花香水月季的遗传多样性,同时也能降低遗传分化的发生。
然而,邱显钦等[24]的研究表明大花香水月季居群的分化程度与地理距离无显著的相关性,这可能是部分居群被孤立或居群内植株数量偏少导致近交和无性繁殖的概率增加,继而加剧了居群间的遗传分化[9]。近年来,随着大花香水月季栖息地缺失、人工采挖等问题的涌现,大花香水月季数量锐减,遗传多样性可能逐步丧失[7,25],同时,生境的片段化、遗传漂变等原因也将加速居群间的遗传分化[26-28]。通常,花瓣长是较为稳定的性状[29-31],但本研究中居群间花瓣长分化明显(VST=26.18%),表明该性状可能受到较强的自然选择作用[32-33]。而萼片宽、花梗长、复叶长、顶叶长宽比、顶叶宽位/顶叶长与顶叶叶柄长在居群间无显著差异且表型分化系数极低,表明这几个性状在居群间较为稳定,不易分化。
3.2 大花香水月季的表型变异规律
表型性状的变异对于植物适应环境及进化有重要作用,受自身遗传物质和生态环境共同的影响[2]。本研究中大花香水月季表型性状的离散程度较低(CV=13.62%),说明梁王山一带大花香水月季遗传多样性可能相对偏低,或者居群内环境条件较为一致。然而,Meng等[9]的研究发现,大花香水月季具有丰富的遗传多样性,其遗传多样性指数远高于弯刺蔷薇[34]、绢毛蔷薇[35]、峨眉蔷薇[36]和长尖叶蔷薇[37]等其他蔷薇属植物。大花香水月季适应性强,分布较广,这与其较高的遗传多样性密不可分,其丰富的变异也能为大花香水月季的种质资源保育和开发提供良好的物质基础。但邱显钦等[24]的研究也表明不同地区大花香水月季居群的遗传多样性存在较大差异。
大花香水月季居群表型性状变异幅度为6.85%~23.78%,不同性状的离散程度不同,体现出不同性状对与环境变化的响应方式不同[38]。其中,大花香水月季花瓣长宽比最稳定,说明花瓣长宽比是大花香水月季表型性状中相对稳定的性状。但同为蔷薇属的中甸刺玫[31]的花瓣长宽比离散程度却较高,表明不同蔷薇属植物表型变异情况存在差异。此外,花瓣长、萼片宽、顶叶长宽比、顶叶宽位/顶叶长和顶叶基角这几个性状在各居群内也相对较为稳定。大花香水月季平均变异系数最大的是花梗长,表明该性状可能拥有较高的表型可塑性,易受到外界条件的影响。总体来看,大花香水月季叶部表型性状变异幅度普遍大于花部表型性状,说明在面临复杂的野外环境时,营养器官通常具有更大的表型可塑性,这与其他植物表型多样性研究的结论相吻合[39-41]。
大花香水月季大部分性状相关性达到显著和极显著水平,反映出这些性状在对环境的适应中表现出相互调节的作用[42]。但花部性状与叶部形状间相关性较低或不相关,说明大花香水月季生殖生长受营养生长影响较小,宽刺蔷薇[43]、华中五味子[39]等研究中也得到了相似的结论。
香水月季人工栽培历史悠久,种内关系复杂,其起源与演化问题一直备受关注[44-46]。了解大花香水月季居群内和居群间的表型多样性,明确其物种特征和表型变异规律,有助于进一步挖掘其经济价值和科研价值,对香水月季的遗传变异和品种分类等方面研究具有重要的指导意义,同时也为大花香水月季和其他蔷薇属植物种质资源的保护、收集与管理等研究提供借鉴与参考。
猜你喜欢
河北科技师范学院学报(2022年2期)2022-08-26
今日农业(2021年18期)2021-11-26
浙江中医药大学学报(2021年6期)2021-07-12
小读者(2021年2期)2021-03-29
中国粮油学报(2020年12期)2021-01-09
文苑(2020年11期)2021-01-04
中国医学影像技术(2019年3期)2019-03-25
草地学报(2018年5期)2018-11-07
中国CT和MRI杂志(2016年11期)2017-01-18
安徽农业科学(2015年10期)2015-04-24
