
早期断奶仔猪脂类代谢变化及分子调控机制研究
2021-06-09 17:22严金昌肖英平王远霞吕文涛李景上刘秀婷
中国饲料 2021年11期
严金昌, 肖英平, 王远霞, 吕文涛, 李景上, 杨 华, 刘秀婷*
(1.嘉兴市秀洲区王店镇政务服务中心,浙江嘉兴314011;2.省部共建农产品质量安全危害因子
与风险防控国家重点实验室(筹),浙江农业科学院农产品质量标准研究所,浙江杭州310021)
随着我国集约化、规模化养猪业的发展,生产上通常对仔猪实施早期断奶,但由于早期断奶仔猪的消化道发育尚未完善,消化液及相关酶分泌不足,胃肠道的微生态区系不稳定且肠道的免疫功能低下,同时断奶过程中仔猪还需要经受环境、心理、营养等一系列的应激,因此早期断奶会给仔猪的正常生长发育带来一系列的影响,如厌食、生长迟滞、消化机能紊乱、腹泻水肿、饲料利用率低等不良现象,称为“仔猪早期断奶综合征”,严重影响我国养猪业的经济效益(寇莎莎等,2018;Wijtten等,2011)。早期断奶的仔猪采食量降低,需要消耗体内的脂肪以供机体的代谢需要(李国俊和崔荣飞,2018)。
参与肝脏脂类代谢的关键酶包括ATP-柠檬酸裂解酶(ACL)、乙酰CoA羧化酶(ACC)和脂肪酸合成酶(FAS)。近年来研究发现,脂肪甘油三酯脂肪酶(ATGL)也是启动脂肪动员的关键脂肪酶,其能特异性地水解甘油三酯(TG)的第一酯键,是TG水解过程的限速酶(Luca等,2019;Zimmerman等,2004)。固醇调节元件结合蛋白-1(SREBP-1)是细胞内调控脂类代谢的关键转录因子,主要通过促进脂肪和胆固醇合成中关键酶基因的表达来促进脂质新生(吴铁梅等,2016)。
早期断奶仔猪的机体代谢状况必然发生变化,然而仔猪机体的脂类物质含量变化及肝脏和脂肪代谢相关基因的表达水平变化仍然未知。因此,本试验拟通过研究早期断奶对仔猪机体脂类物质含量和肝脏及脂肪组织中脂类代谢相关基因表达水平的影响,探究影响早期断奶仔猪代谢的关键因素,为早期断奶仔猪的营养代谢功能调控研究奠定理论基础。
1 材料与方法
1.1 试验材料 选取4个窝的16头18日龄仔猪,将来自于不同窝的4头仔猪称重,采血后直接用于屠宰取样,作为基础数据;另外12头进行断奶处理,为断奶组,每4头一起饲养在与母猪栏环境相同的小栏中,饲喂配合饲料。母猪正常哺乳窝中剩余的仔猪,为非断奶组。在断奶后第1、3、7天(分别对应仔猪19、21、25日龄),从断奶组和非断奶组各取4头仔猪称重,采血后屠宰取样。试验期间自由饮水和采食。哺乳母猪饲粮组成及营养水平见表1(杨华等,2018)。仔猪饲料由浙江国茂饲料有限公司提供,参考美国NRC(1998)标准配制,具体组成及营养水平见表2。
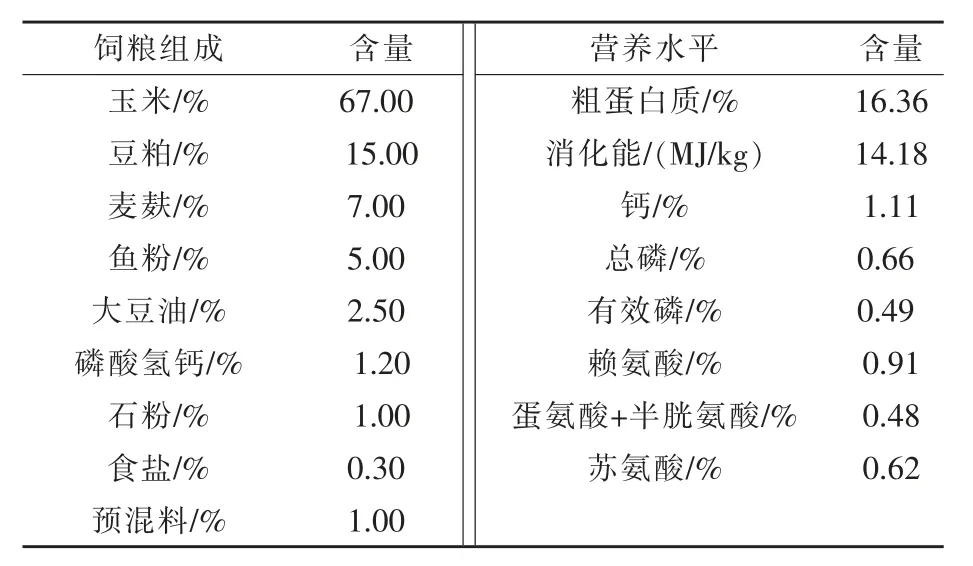
表1 哺乳母猪饲粮组成及营养水平(饲喂基础)
1.2 样品采集 于分组处理后0、1、3 d和7 d分别从断奶组和非断奶组选取4头仔猪进行前腔静脉采血,4℃下3000g离心10 min后得到血清样本,然后屠宰取肝脏与背最长肌样本,于-80℃保存备用。
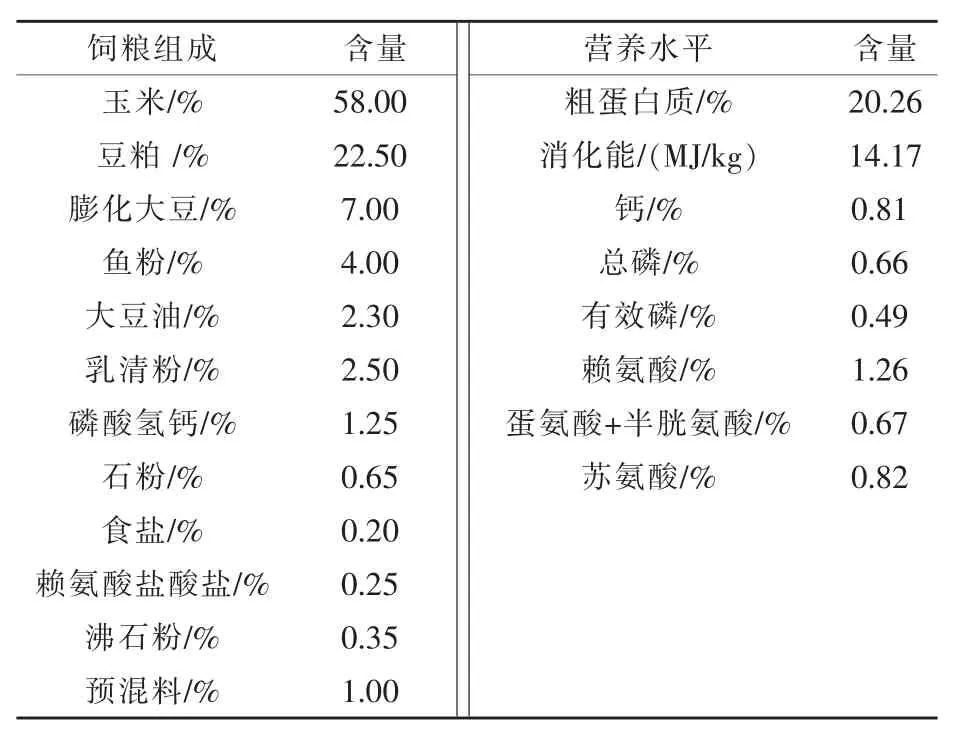
表2 断奶仔猪饲粮组成及营养水平(饲喂基础)
1.3 血清脂类物质含量的测定 采用南京建成生物工程研究所提供的试剂盒分析血清中甘油三酯(TG)、胆固醇、高密度脂蛋白(HDL)和低密度脂蛋白(LDL)指标,试验操作按照相应的检测试剂盒说明书进行。
1.4 脂类代谢相关基因表达分析 采用总RNA抽提试剂盒提取肝脏组织的总RNA,利用Nano Drop微量分光光度计测定RNA的纯度,以OD260nm/OD280nm评价RNA的纯度,琼脂糖凝胶电泳检测RNA完整性。使用Invitrogen反转录试剂盒SuperscriptTMⅡRTase反转录cDNA。
采用Premier 6.0和Beacon designer软件进行目的基因和18S rRNA内参基因引物的设计,引物序列见表3。采用TaKaRa SYBR实时荧光定量试剂盒SYBR Premix Ex TaqTM进行基因的实时荧光定量检测。PCR扩增反应体系为25μL,具体 如 下:ddH2O 10.5μL,SYBR Premix Ex TaqTM(2×)12.5μL,上、下游引物各1μL,cDNA模板1.0μL。反应条件:95℃,1 min;45个循环:95℃,10 s;62℃,25 s;由55℃连续上升到95℃,每5 s温度上升0.5℃,以制备熔点曲线。

表3 脂肪合成相关目的基因和18S rRNA内标基因引物参数
1.5 数据处理 肝脏基因表达数据根据系统自动分析的ACL、FAS、ACC、ATGL和SREBP 1以及18S rRNA标准曲线的斜率得到PCR扩增效率(E),E=10(-1/slope)。ACL、FAS、ACC、ATGL和SREBF 1的扩增效率都接近100%,且相互间效率偏差在5%以内,以目的基因mRNA的拷贝数与对应样品内参基因mRNA的拷贝数的比值来表示基因mRNA的相对表达量,各个基因的相对表达水平以2-△△Ct进行统计分析。每个待测样品设置3个重复,对检测得到的3个Ct值取算术平均值(杨华等,2018)。血清中脂类物质的含量以及基因表达所有数据采用SPSS 16.0软件进行单因素方差分析。试验结果采用“平均值±标准误”表示,P<0.05表示差异显著。
2 结果与分析
2.1 早期断奶对仔猪血清脂类物质含量的影响从图1可看出,早期断奶可显著降低仔猪血清TG、总胆固醇、HDL和LDL含量,并表现出一致的变化规律,在断奶后第3天开始下降。
与非断奶仔猪比较,仔猪在断奶后的第3、7天,血清TG含量分别降低了26.51%(P<0.05)和36.71%(P<0.05);胆固醇含量分别降低了26.18%(P<0.05)和40.93%(P<0.05);HDL含量分别降低了29.69%(P<0.05)和31.34%(P<0.05)。断奶仔猪血清LDL含量在断奶后的第1、3、7天分别比非断奶仔猪降低了24.05%(P<0.05)、37.65%(P<0.05)和23.17%(P<0.05)。

图1 早期断奶仔猪和对应同日龄哺乳仔猪血清TG、胆固醇、HDL和LDL含量
2.2 早期断奶对仔猪肝脏脂类代谢相关基因表达的影响 由图2可知,早期断奶处理可提高仔猪肝脏中ACL基因的表达水平,但是肝脏中ACC、FAS和SREBP-1基因的表达水平降低,并表现为一致的变化趋势。与非断奶同日龄仔猪比较,在断奶后第1、3、7天,断奶仔猪肝脏ACL基因的mRNA表达水平分别提高了113.33%(P<0.05)、106.15%(P<0.05)和100.00%(P<0.05),ACC表达水平分别降低了26.67%(P<0.05)、53.51%(P<0.05)和33.86%(P<0.05),FAS表达水平分别降低了72.57%(P<0.05)、85.71%(P<0.05)和27.66%(P<0.05),SREBP-1表达水平分别降低了58.72%(P<0.05)、47.69%(P<0.05)和11.69%(P<0.05)。
2.3 早期断奶对仔猪脂肪组织脂类代谢相关基因表达的影响 如图3所示,早期断奶使仔猪皮下脂肪组织中ATGL的表达水平升高,使FAS的表达水平降低。在断奶后的第1、3、7天,脂肪组织中ATGL表达水平分别升高了51.02%(P<0.05)、50.00%(P<0.05)和26.50%(P>0.05),FAS表达水平分别降低了73.33%(P<0.05)、60.17%(P<0.05)和37.10%(P<0.05)。
3 讨论
哺乳动物多余的能量主要以甘油三酯(TG)的形式储存在脂肪组织中,当动物处于饥饿或当机体能量匮乏时,贮存的脂质就会发生脂解,由TG分解成非酯化的脂肪酸,释放进入血液参与新陈代谢提供能量,脂肪的储存及动员处于动态平衡状态(Bell等,2005;Flier,2004);沉积发生在仔猪初生后的各个生长发育阶段。Lodge等(1978)研究指出,初生时期仔猪的体脂肪含量仅为1%~2%(脂肪型猪约为4%),随着日龄增加脂肪沉积逐渐增多,1周龄时体脂量约7%,2周龄时为12%~15%,到4周龄时体脂量是初生时的90倍左右,但是体重仅为5倍多;16周龄后,虽然脂肪沉积量仍在增加,但其脂肪合成率开始下降。
早期断奶的仔猪因受到断奶应激的影响,会通过消耗机体的脂肪以满足其代谢需要。ACC和FAS是体内脂肪酸合成的关键酶。ACC是长链脂肪酸从头合成的限速酶,催化乙酰辅酶A生成丙二酸单酰辅酶A,其活性的高低可影响肝脏及肌肉组织中的脂肪代谢(Sone等,2016)。FAS存在于脂肪、肝脏及肺等组织中。动物的体脂沉积所需要的脂肪酸大多来自于脂肪酸的全程合成,即由FAS催化乙酰CoA和丙二酰CoA生成长链脂肪酸,然后再合成甘油三脂(TG)(Simith等,2003)。FAS活性的高低直接控制着体内脂肪合成的强弱,对控制动物体脂的沉积具有重要的调控作用(Wang等,2004)。FAS表达水平升高能显著增加TG在体内的沉积,从而导致肥胖。脂肪组织中FAS与酮体脂肪量及脂肪率成极显著正相关。通过抑制FAS的活性,可阻滞脂肪产生通路,减少脂肪合成,升高丙二酰CoA浓度,从而达到降低饮食的目的。
在仔猪早期断奶阶段,采食日粮由含乳糖、乳蛋白和乳脂等易于消化的液态乳汁转变成以玉米-豆粕为主的固态日粮,日粮的转变使其采食量急剧下降,能量摄入骤减,从而导致仔猪能量代谢处于负平衡状态(Xiao等,2012)。Pié等(2004)研究发现,仔猪在断奶后的24 h内仅采食到了11 g饲粮,在断奶后2 d和8 d分别增加至214 g和384 g。在本试验中发现,早期断奶仔猪血清中TG、胆固醇、HDL、LDL等脂类物质含量均降低,同时肝脏ACC、FAS和SREBP-1基因表达水平显著降低,脂肪组织中FAS基因表达水平也相应地降低,表明早期断奶使仔猪脂肪合成能量下降,仔猪早期断奶阶段为了维持机体的能量供给,动员已储存的脂肪,通过脂肪的脂解作用,保障机体能量代谢的平衡。ACL是糖代谢与脂肪酸合成代谢之间的桥梁,位于脂质代谢通路的最上游,其活性受磷酸化和去磷酸化途径的调节,磷酸化酶是ACL活性形式(Sun等,2010)。研究表明,在早期断奶仔猪的肝脏中ACL基因表达量升高,这可能是由于机体糖类代谢的需要,提高了ACL基因的表达水平以参与机体的糖类代谢。
ATGL是一种启动脂肪细胞脂动员的脂肪分解酶,能够特异性地分解TG(Zimmermann等,2004)。猪、人和鼠的ATGL主要在脂肪组织中表达,是脂肪及其他组织中TG水解起始步骤的限速酶(Shan等,2008;Jenkins等,2004)。ATGL基因表达水平受到日粮、激素等因素的影响。当动物限饲或禁食时,ATGL将TG分解成游离脂肪酸(FFA)以供其他组织利用。Kershaw等(2006)研究指出,限饲可诱导鼠ATGL基因的表达。Villena等(2004)研究发现,大鼠限饲12、24 h和48 h,脂肪组织中ATGL表达水平先升高后降低;重新补饲后,ATGL表达水平继续下降。本试验发现,在早期断奶仔猪的脂肪组织中,ATGL表达水平显著升高,与前人的研究结果(Kershaw等,2006)相一致。
4 结论
仔猪在早期断奶过程中,血清中甘油三酯(TG)、胆固醇、高密度脂蛋白(HDL)和低密度脂蛋白(LDL)含量降低。同时,肝脏脂类代谢的关键酶乙酰辅酶A羧化酶(ACC)、脂肪酸合成酶(FAS)和固醇调节元件结合蛋白-1(SREBP-1)基因表达水平均降低,皮下脂肪组织中FAS基因表达水平降低,甘油三酯脂肪酶(ATGL)的基因表达水平升高,反映了仔猪在早期断奶阶段通过调节肝脏及脂肪组织的脂类代谢关键酶相关基因的表达水平动员脂肪,从而为机体提供能量以保证供给平衡。
猜你喜欢
今日农业(2022年2期)2022-11-16
昆明医科大学学报(2022年1期)2022-02-28
中老年保健(2021年9期)2021-08-24
青岛大学学报(医学版)(2020年6期)2020-11-16
考试与评价·高二版(2020年1期)2020-09-10
家庭科学·新健康(2018年12期)2018-03-01
江苏农业科学(2016年1期)2017-05-17
家庭医学(2015年6期)2015-07-03
健康博览(2015年3期)2015-07-03
新校园·中旬刊(2014年10期)2014-12-01
