
奶牛热应激遗传机制研究知识图谱分析
2021-05-28 07:53:28张海亮罗汉鹏米思远邱文卿王雅春
畜牧兽医学报 2021年5期
张 帆,张海亮,罗汉鹏,米思远,邱文卿,初 芹,王雅春*
(1.新疆农业大学动物科学学院,乌鲁木齐 830052;2.中国农业大学动物科技学院,北京 100193;3.北京市农林科学院,北京 100089)
泌乳奶牛正常体温为38.5~39.3 ℃,其适宜的环境温度范围为5~25 ℃[1]。当环境温度高于25 ℃时,奶牛机体受到超过自身体温调节能力的高温刺激,作用于垂体-肾上腺皮质系统而引起机体非特异性防御反应和特异性障碍在内的全身性适应症,即产生热应激综合征(下文简称热应激)[2]。热应激状态下,奶牛的采食量下降,奶牛易处于能量负平衡状态,从而降低产奶量和繁殖性能。随着全球气候变暖,热应激给畜牧业带来的危害愈发严重,仅在美国,每年因热应激造成的畜牧业直接经济损失高达23.6亿美元[3]。了解奶牛热应激的遗传机制,选育奶牛的热应激抗性已成为奶牛育种的新目标[4]。
奶牛热应激抗性是一个复杂性状,其遗传力较低。传统育种对热应激抗性的选育难度较大,分子育种手段为热应激抗性性状的选育提供了新的可能[5],研究奶牛热应激遗传机制是开展上述工作的基础。随着分子生物学及分子遗传学的发展,奶牛热应激遗传机制的研究也不断深入。在2010年以前,主要集中在热应激对细胞和早期胚胎发育的影响,主要包括卵母细胞和囊胚前期胚胎[6-7]。2010年以后,研究逐步深入到热应激相关基因多态性和转录水平[4,8-9]。此外,热应激还可以通过增加热休克转录因子1(heat shock transcription factor 1,HSF1)活性提高热应激相关基因的转录水平[10],通过某些miRNA促进或抑制一些基因的表达。例如,热应激可通过miRNA-541影响HSP27的表达[11]。当下,国际上有关奶牛热应激遗传机制研究的文献与日俱增,2000年以前,该领域年平均发文量在20篇以下,2010年之后,年平均发文量达50篇以上,该领域的年发文量目前仍在高速增加。因此,对该领域的科技文献进行深入挖掘十分必要,能帮助科研工作者及时了解奶牛热应激遗传机制领域及其研究热点,筛选有价值的信息。
知识图谱分析是将应用数学、图形学、信息可视化技术和信息科学等学科的理论方法与计量学引文分析、共现分析等相结合,利用可视化图谱展示学科的核心框架、发展历史、前沿领域和整体知识框架的多学科融合的一种研究方法[12]。Citespace V软件系统是一款用于计量和分析科学文献数据的信息可视化软件,其主要利用Web of Science核心合辑数据库,通过主题搜索和下载搜索记录(全记录与参考文献)获得供其分析的原始数据进行分析。本文利用Citespace V对国际和国内有关奶牛热应激遗传机制的科技文献进行了可视化分析,包括文献共被引分析和关键词共现分析,剖析了国内外该领域的研究趋势,对一些高被引文献和高产作者进行了解读,并对热应激的部分转录调控方式进行了简要分析。
1 数据来源与研究工具
1.1 数据来源
为了解国际上奶牛热应激遗传机制研究的总体情况,本文对该领域近 30 年文献进行了知识图谱分析。所收集的数据来自美国汤森路透公司开发的Web of Science专业信息检索平台中的核心合集。以TS=((“Thermal stress” OR “Temperature stress” OR “Heat shock” OR “Heat tolerance” OR “Heat resistance”) AND (“Gene*”) AND (“Cattle” OR “Dairy cow” OR “Holstein” OR “Bovine” OR “Brahman” OR “Senepol” OR “Sahiwal” OR “Jersey”))为检索式进行了主题检索,检索时间段为数据库默认的1990—2020年,共检索到1 026条文献数据。分析的基本单元包括作者姓名、标题、摘要、发表日期、研究单位和引文,数据库的截止时间为2020年7月14日。
为了对比国内在相关领域的研究近况,本文还以SU=(热应激+热休克+耐热性)*(基因+遗传+表达+转录+调控)*(牛+荷斯坦+反刍动物+羊)为检索式在中国知网进行专业检索,检索时间段为1990—2020年,共检索到 52 篇文献。数据库的截止时间为2020年7月14日。
1.2 研究工具
本研究运用文献计量学方法,同时,借助绘制学科知识图谱展示和分析奶牛热应激遗传机制的研究状况和研究热点,使用的文献信息可视化工具为Citespace V软件[13]。
2 奶牛热应激遗传机制研究的国际发文情况
2.1 奶牛热应激遗传机制研究的国际发文量
第一篇关于牛热应激的文章发表于1979年,其后10年间未发表过相关文献,直至1990年,热应激遗传机制的研究再次出现在人们的视野中;1990—2020年,奶牛热应激遗传机制领域的国际发文量如图1所示。这一时期,该领域的国际年发文量呈上升趋势。根据Web of Science数据库的检索结果,2006年之后该领域的年发文量激增。2020年数据采集时的最后更新时间为2020年7月14日,该年度的发文量为不完全统计。
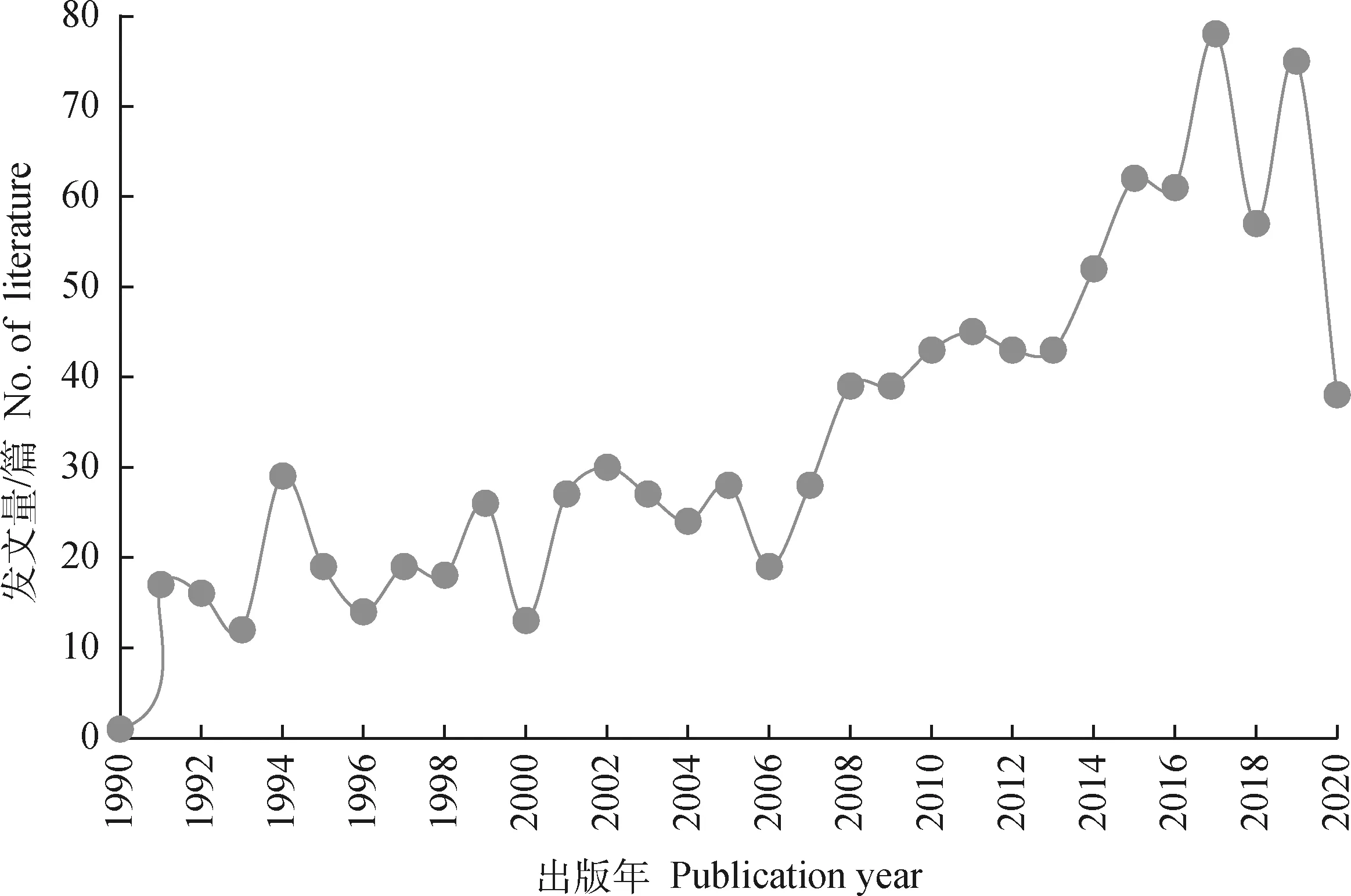
图1 1990—2020年奶牛热应激遗传机制领域的国际发文量Fig.1 The number of international literature published annually on genetic mechanisms of heat stress in dairy cows from 1990 to 2020
2.2 奶牛热应激遗传机制研究的高被引文献
通过共被引文献分析,国际上奶牛热应激遗传机制研究影响力较大的时期为2008—2012年,高被引文献主要集中于该时期,被引频次最高的前10篇文献[14-23]见表1。前10篇高被引文献中,5篇为综述类文章,5篇为研究性文章。前10篇高被引文献中的5篇发表在《Journal of Dairy Science》期刊上,该杂志是发表奶牛热应激遗传机制研究的主阵地。
被引频次排名第1的文献是Collier等[14]于2008年发表在《Journal of Dairy Science》期刊上名为“Invited Review:Genes involved in the bovine heat stress response”的综述。该文章首先提出当奶牛皮肤温度超过35 ℃时开始储存热量,个体的基因表达开始出现一系列变化,包括:1)热休克转录因子1(heat shock transcription factor 1,HSF1)激活;2)热休克蛋白(HSP)表达增加;3)糖代谢增加;4)内分泌系统激活;5)外泌热休克(HSP)蛋白激活免疫系统。Collier等在文章中介绍了牛对热应激的适应可能是由一系列基因表达变化所导致,结合最新的研究进展,表观遗传学修饰可能是造成基因表达改变的原因之一。第5篇是Basiricò等[18]于2011年在《Cell Stress Chaperones》上发表的“Cellular thermotolerance is associated with heat shock protein 70.1 genetic polymorphisms in Holstein lactating cows”,该研究发现,HSP70启动子区域的两个SNPs与奶牛原代培养细胞(peripheral blood mononuclear cells,PBMC)的mRNA表达有关,能够降低热应激下PBMC的凋亡率,并指出,这两个SNPs能增强细胞对热应激的免疫应答能力。第7篇文献[20]“Heat shock-induced apoptosis in preimplantation bovine embryos is a developmentally regulated phenomenon”介绍了热应激导致定植前牛胚胎凋亡率增加,且这种凋亡主要发生在16细胞期,作者还发现,胚胎早期经历轻度热应激可增加胚胎的耐热性。第8篇文献[21]“Genetic components of heat stress for dairy cattle with multiple lactations”利用生产性能数据,通过随机回归测定日模型估计了奶牛热应激耐受性的方差组分,结果显示,热应激耐受性的遗传方差随胎次的增加而增加,不同胎次之间热应激耐受性的遗传相关均高于0.8。第10篇文章[23]为“Effect of heat stress on the expression profile of Hsp90 among Sahiwal (Bosindicus) and Frieswal (Bosindicus×Bostaurus) breed of cattle:A comparative study”,该研究对比了夏季沙希华牛(Sahiwal)和荷斯坦牛血液中热休克蛋白90(heat shock protein 90,HSP90)含量及PBMC在体外热应激情况下的活力,发现沙希华牛血液中HSP90含量和PBMC活力均显著高于荷斯坦牛,作者认为血液中的HSP90增加了牛的耐热性。
2.3 研究奶牛热应激遗传机制的主要机构与作者
2.3.1 主要发文机构 1990—2020年,奶牛热应激遗传机制研究的主要机构按发文频次排序的前10位如表2所示,这些机构分别位于美国东南部、巴西、法国、印度、中国和澳大利亚,主要处于热带和亚热带地区,这些国家和地区所饲养的牛群均会受到不同程度的热应激。这10个机构的论文产出量之和占全部文献(1 026篇)的26.2%。

表2 奶牛热应激遗传机制研究发文量前10的研究机构Table 2 The top 10 institutions with the greatest number of paper on the genetic mechanism of heat stress in dairy cows
2.3.2 主要发文作者 根据Web of Science数据库的检索结果,1990—2020年发文量排序前10名作者如表3所示,这些作者发表的文献从9~43篇 不等。对发文量较高作者的研究方向进行分析发现,Peter团队主要进行牛繁殖过程与热应激有关的研究,其在转录组水平[24]、全基因组水平[25]、基因水平[26]和生理水平[27]均进行了研究,研究对象主要为牛卵母细胞和早期胚胎。例如,2016年其发文介绍了HSPA1L(heat-shock 70-kDa protein-1-like,70kDa热休克蛋白1样蛋白基因)启动子区域的缺失突变有利于受精卵在热应激状态下存活[28]。Kumar团队对热应激遗传机制进行了相关研究,2003年其发表的第一篇文章指出,HSP70(heat shock protein 70,热休克蛋白70)基因启动子区AP2盒区域的胞嘧啶缺失导致热应激时HSP70基因表达水平降低,从而降低了奶牛的耐热能力[29];其发表最高被引文章指出,耐热性更好的沙希华牛PBMC的活力和HSP90基因的表达量均高于荷斯坦牛[30];在最近的研究中,其从单个基因的表达水平拓展到全基因组水平的转录研究,2020年对热应激下荷斯坦牛皮肤成纤维细胞进行了全基因组表达分析发现,上调基因主要影响蛋白质加工和NOD样受体(NOD-like receptors)途径,而下调基因则与细胞周期、新陈代谢和蛋白质转运有关[31]。
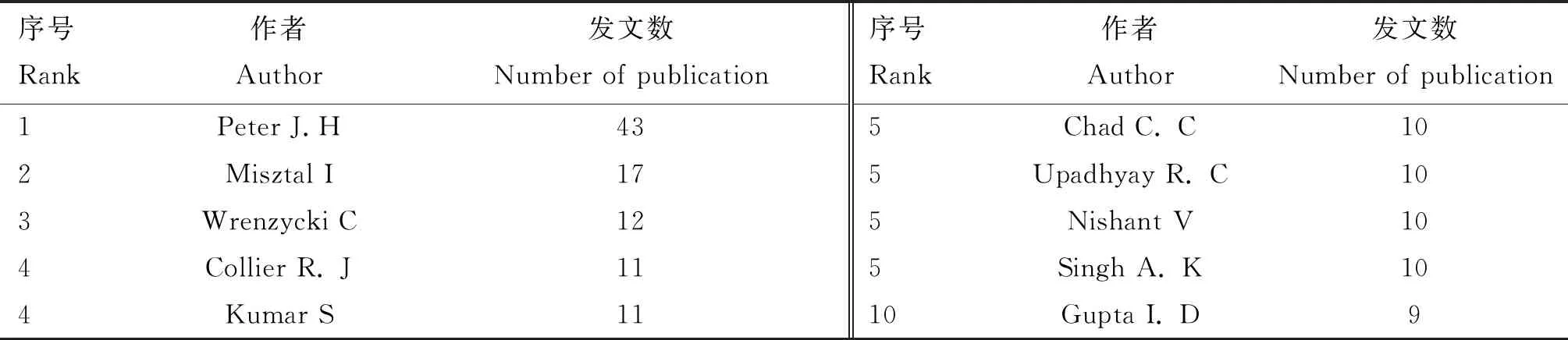
表3 奶牛热应激遗传机制研究发文量频次排前10的作者Table 3 The top 10 authors with the greatest number of paper on the genetic mechanism of heat stress in dairy cows
3 国际上奶牛热应激遗传机制研究的热点词
3.1 高频热点词汇的界定
本文对奶牛热应激遗传机制文献中出现的关键词频次(以软件中统计的频次为准)和时间段进行了统计,以透视该领域不同时期的科研热点及发展历程。图2 和 图3 展示了1990—2020年奶牛热应激遗传机制研究中热点词汇和突现强度排名前20的词汇。通过“gene”、“expression”、“heat shock protein”、“chaperone”、“embryo”和“bovine oocyte”这些词汇能看出,奶牛热应激遗传机制的前期研究主要以热休克蛋白和其他相关基因的表达及作用为主要切入点,研究的主要对象包括卵母细胞和胚胎。“thermotolerance”、“SNP”和“oxidative stress”的突现时间一直持续到现在,说明这些主题是目前的研究热点,筛选牛耐热性的遗传标记成为了热门话题,热应激引起氧化应激也逐渐被大家所关注。
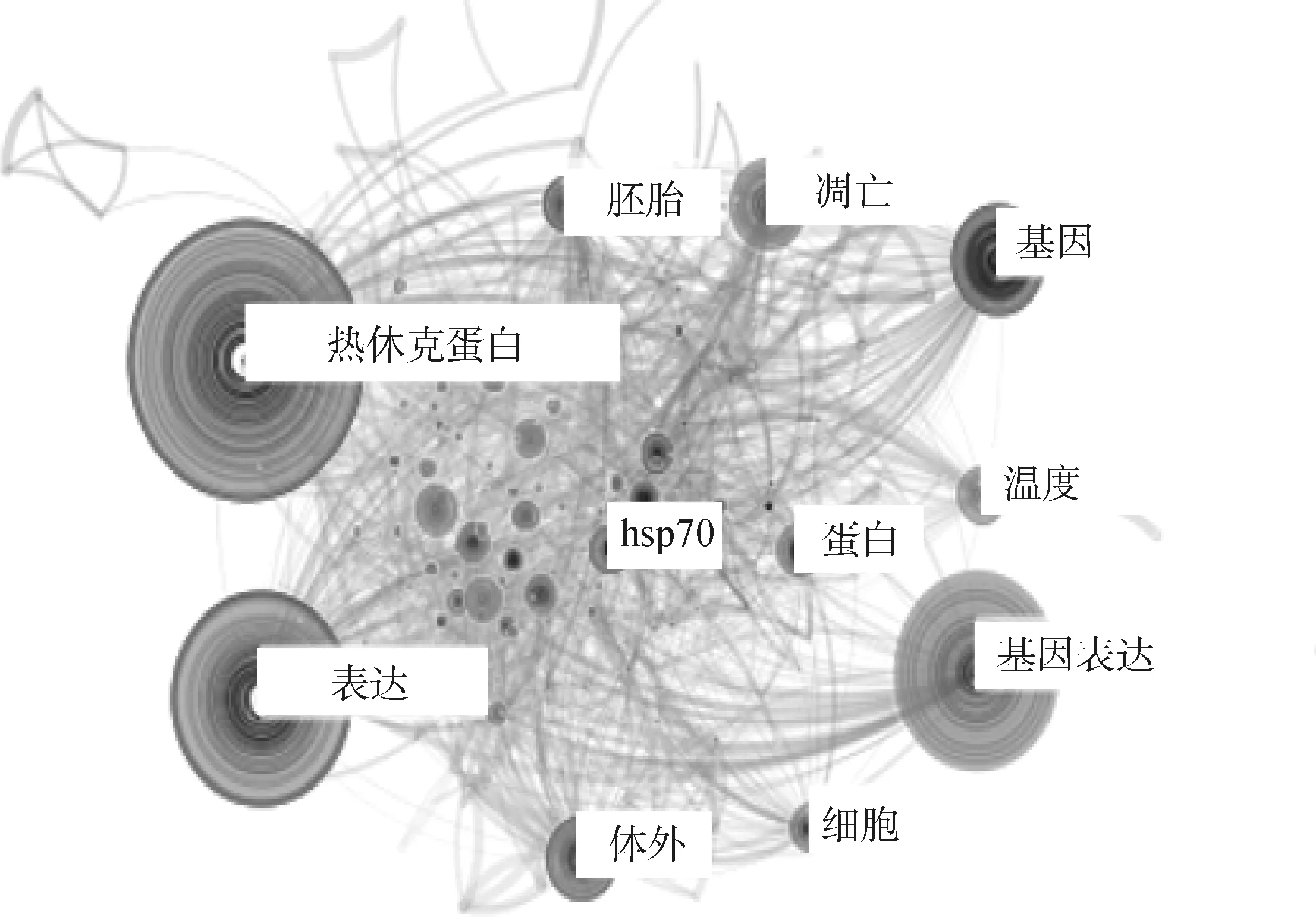
图2 奶牛热应激遗传机制研究的热点词汇Fig.2 Hot vocabulary on genetic mechanism of heat stress in dairy cows
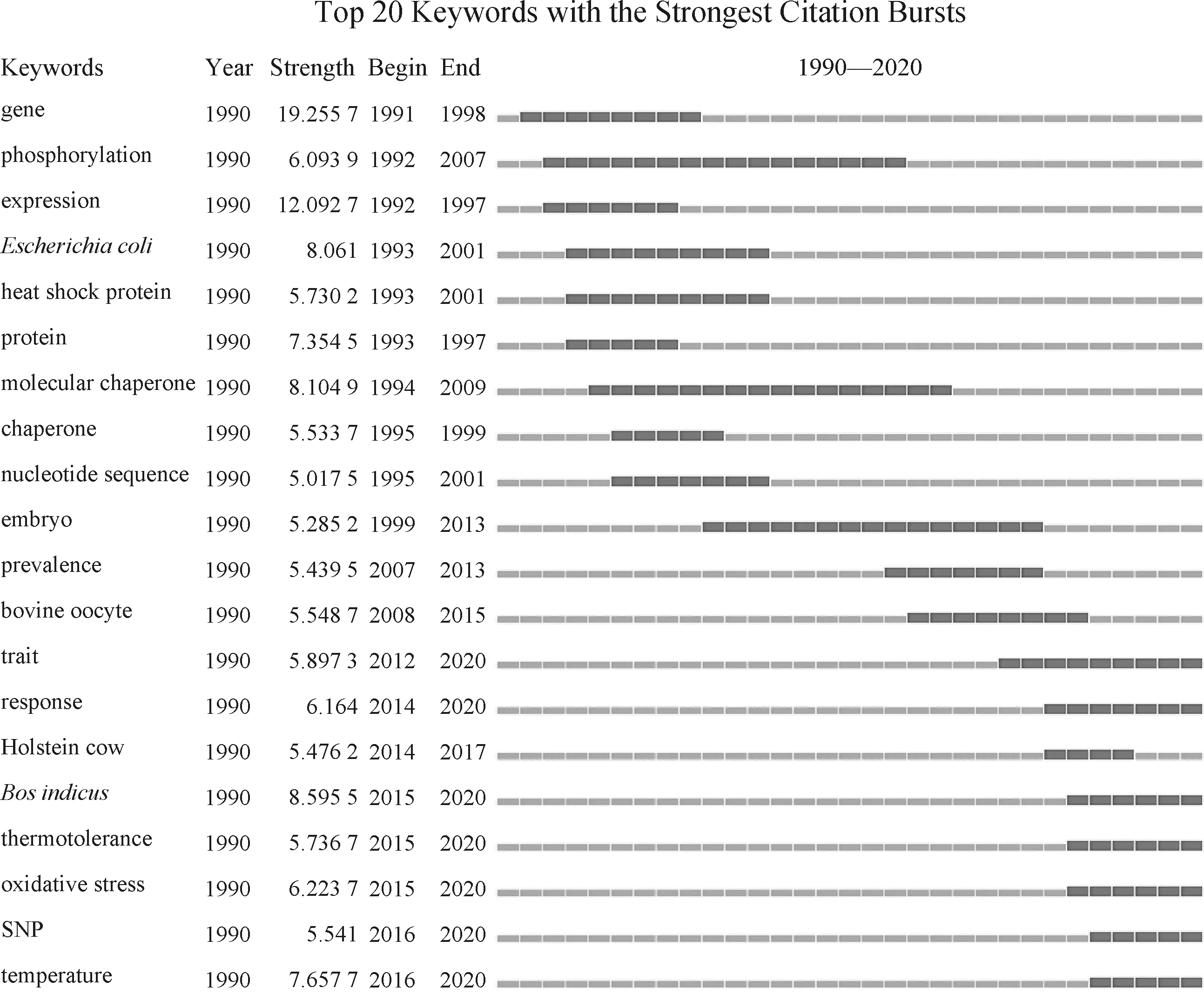
图3 1990—2020年突现强度前20的热点词汇Fig.3 The top 20 keywords with the strongest citation bursts from 1990 to 2020
3.2 奶牛热应激遗传机制领域的研究状况
3.2.1 热应激对奶牛繁殖性能的影响 热应激状态下,奶牛卵巢的生长发育活动与功能被抑制,发情表现变差、卵泡发育和生长减慢、卵母细胞质量变差、繁殖性能降低[32]。此外,高温高湿环境易引起奶牛阴囊和睾丸温度的升高,造成睾丸生殖上皮变性,影响精子的发育与成熟,改变精液品质[33]。由于热应激对奶牛繁殖性能的重大影响,且繁殖过程中生殖细胞极易受表观遗传修饰的影响,故热应激对奶牛繁殖性能的影响机制一直受到关注。公畜繁殖力的研究中发现,在睾丸中过氧化物酶体增殖物活化受体γ共激活因子-1(peroxisome proliferator-activated receptor γ coactivator-1,PGC-1)和HSP70可以保护生殖细胞和睾丸间质细胞免受热应激和氧化应激的影响[34]。此外,Sigdel等[35]通过全基因组测序及通路富集分析发现,受精和发育、蛋白质错误折叠、细胞对氧化应激的反应和钙离子稳态这些过程中显著富集了热应激下与奶牛受孕能力相关的基因。
3.2.2 奶牛热休克蛋白 热休克蛋白是机体在不利环境因素刺激下,合成的一组特殊蛋白[36]。其在应激条件下合成增加,主要生物学功能包括分子伴侣、胚胎发育、细胞凋亡和免疫应答等。鉴于热休克蛋白(heat shock protein,HSP)在细胞分裂、分化和凋亡等生命基本过程中的关键作用,大量研究集中于HSP及其基因表达的反应调节,热应激条件下产生的HSP及其相关基因的表达将会对个体的生理功能产生不同程度的影响,这些基因可能对奶牛热应激的研究有重要意义,为筛选耐热、高产奶牛提供理论依据。
3.2.3 奶牛热应激相关的单核苷酸多态及候选基因 随着测序技术的发展,挖掘与热应激相关的SNP和候选基因已经成为了热应激遗传机制研究领域的新热点。自2016年起,相关文献的数量呈现激增趋势并且延续至今,所涉及的基因包括HSF1[37]、HSPB8[38]、ASHA2[39]、MYO1A[40]、MTOR[41]、HSP70-1[42]、EIF2AK4[43]、HSP90AA1[44-45]和ATP1A1[46]等,部分SNPs和候选基因信息见表4。与热应激相关的SNPs位于这些基因的各个位置,包括内含子、外显子和3′UTR等。
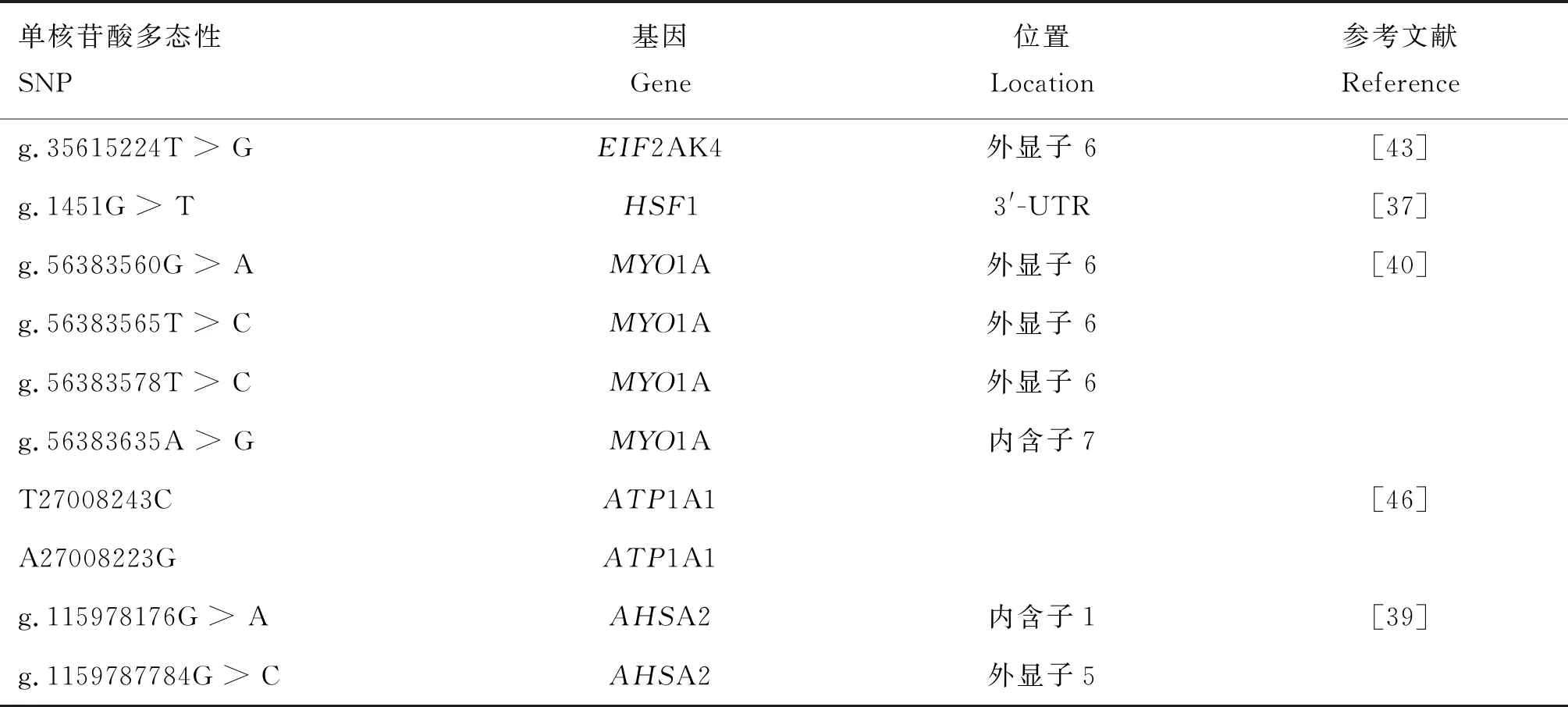
表4 部分与奶牛热应激相关的部分SNPs和基因Table 4 Some SNPs and genes related to heat stress in dairy cows
3.2.4 氧化应激(oxidative stress) 在机体正常生理状态下,线粒体电子传递过程中总氧气的1%[47]~24%[48]会通过电子泄露而被还原成超氧化物,再通过超氧化物歧化酶还原成H2O2,再进一步降解。正常的生理状态下,机体的氧化和抗氧化随时存在并处于平衡状态,并不会产生氧化应激而影响机体健康[49]。当热应激发生时,细胞的能量需求增加;急性热应激时,细胞的能量需求是正常情况的两倍[50-51]。线粒体产能增加后,电子载体(如辅酶Q)大量被还原[52]、电势差变大[53]和线粒体内氧浓度升高等可能会导致活性氧(ROS)增加。随着研究的深入,越来越多的研究人员意识到热应激在诱导产生氧化应激过程中的重要作用[54-55]。
4 奶牛热应激遗传机制研究国际发展脉络及现状
1990—2008年,奶牛热应激遗传机制的研究主要关注热休克蛋白基因和其他相关基因在热应激时表达量的变化、热休克蛋白对细胞的保护作用[56-57]以及热应激所引起的生理变化情况[45,58-59]。2008—2014年是热应激遗传机制研究的转折点,这期间相关文献呈突发性增长,对热应激相关基因表达的研究更加深入,同时,开始出现对热应激相关基因多态性的研究。2014—2020年,筛选热应激相关SNP的研究成为新的热点[45],同时,研究人员也对表观遗传和miRNA在热应激转录调控过程中所起的关键作用进行了深入探究。此外,热应激对动物繁殖性能的影响也成为了研究的热点[60-62]。
目前,最新的研究显示,热应激涉及复杂的转录调控网络,并且越来越多的证据表明,表观遗传调控和miRNA在热应激诱导的转录调控中起着重要的调控作用。在Shen等[63]的研究中,热应激能够导致细胞质中miRNA-541表达量减少,使HSP27表达量增加,进而抑制热应激下细胞凋亡,促进细胞自噬。此外,miRNA-370和miRNA-543显著下调HSP40的表达量[64],miRNA-122通过靶向调控FOXO3基因mRNA进而调控HSP70的表达量[65],上述研究都证明,miRNA在热应激调控机制中起到关键的作用。
表观遗传也能通过调控HSP70进一步调节热应激反应。例如,2017年,Kisliouk等[66]发现,热应激会使HSP70基因启动子区域CpG甲基化水平升高,减少核小体去乙酰化酶的募集,增加HSP70启动子上的组蛋白3乙酰化水平,并最终导致HSP70的转录增加。2020年,Kisliouk等[67]又发现,热应激直接引起zeste同源物2的增强子(enhancer of zeste homolog 2,EZH2)和脑源性神经营养因子(brain-derived neurotrophic factor,BDNF)mRNA N6-腺苷酸甲基化(m6A)水平增加,EZH2和BDNF基因表达水平降低,并降低雏鸡的耐热性。此外,最新研究发现,表观调控和miRNA对热应激调控具有协同作用。例如,Rosenberg等[68]发现,热应激通过miRNA-26a降低EZH2的转录水平,导致IL6内含子1上组蛋白3的甲基化水平增加,从而减少了NFkB与白细胞介素-6(Interleukin-6,IL6)内含子1结合,最终提高雏鸡耐热抗性。
5 国内奶牛热应激遗传机制研究进展
目前,我国研究人员针对奶牛热应激遗传机制进行的研究较少。因此,将其他畜种的相关研究也纳入考察范围。共搜索到52篇相关文献。其中,以热应激、热休克蛋白和多态性为主题的文献最多,共计35篇,7篇以中国荷斯坦牛为研究对象。2008—2010年间共发表15篇文献,占总文献的29%,在2009年前后,有关文献发表量最多。目前,国内对奶牛热应激遗传机制的研究较少,研究不够深入,这一领域还处于初级阶段。
在国内,热休克蛋白同样是热应激遗传机制研究的热门话题,包括HSP70、HSP90和sHSP等。例如,程维杰等[42]发现,HSP70基因编码区1 623位点的SNP与奶牛直肠温度显著相关,GA个体的直肠温度显著低于GG和CC个体;吴国良等[69]发现,HSP70基因3′侧翼区存在与热应激相关的SNP,并且该位点与HSP70的转录相关。2009年以前,国内对热应激遗传机制的研究主要集中于:1)热应激相关基因在热应激条件下的差异表达情况[70-72];2)热休克蛋白及其他热应激相关基因对热应激下细胞的保护作用[73]。2009—2014年,从研究热休克蛋白基因及其他相关基因的表达情况逐步拓展到对这些基因多态性的研究,进一步挖掘这些基因上存在的耐热遗传标记[74-75],热门的基因有HSP70[75]、HSF1[76]和PPAR-α[77]等。2014—2020年,开始分析热应激信号通路[78]及研究热应激相关的非编码RNA[79]。例如,胡煜等[80]发现,热应激会显著下调miRNA181a和miRNA486的表达,极显著上调miRNA486的表达;此外,郑月等[81]也发现,miRNA181a在热应激奶牛的血清中显著下调,miRNA181a 的靶基因与应激反应以及免疫功能密切相关。
6 总结与展望
基于Web of Science核心合辑数据库和中国知网收录的国际和国内有关奶牛热应激遗传机制的文献,本文运用Citespace V软件绘制了该领域的共现知识图谱,分析了该领域的研究和发展情况,并重点分析了高被引文献及研究前沿的热点词汇。国际上,从1990年开始,研究奶牛热应激遗传机制的热潮一直延续至今,该领域的发文量和研究热度不断提高。热休克蛋白、单核苷酸多态性、氧化应激和与繁殖性能的关系一直是该领域的热门话题。在热应激相关基因的表达中,表观修饰和miRNA是目前研究最广泛的两种调控方式。随着测序技术发展和多组学概念的引入,远程增强子调节基因表达、非编码RNA调控网络、组蛋白表观修饰等调控机制可能成为研究热应激遗传机制的新热点。将来,更多与热应激相关的候选基因、通路和相关调控机制被挖掘出来,将为我们提供充足的奶牛耐热性的遗传标记,为奶牛耐热性选育奠定理论基础。
猜你喜欢
今日农业(2022年3期)2022-11-16 13:13:50
今日农业(2021年10期)2021-07-28 06:28:00
今日农业(2020年14期)2020-08-14 01:10:16
家庭医学(下半月)(2020年4期)2020-05-30 12:42:40
小天使·一年级语数英综合(2019年12期)2019-01-13 01:32:29
阅读(低年级)(2018年10期)2018-05-14 20:42:01
阅读(低年级)(2018年11期)2018-05-14 09:37:53
阅读(低年级)(2018年12期)2018-03-23 11:39:02
哈尔滨医药(2015年4期)2015-12-01 03:57:56
小天使·三年级语数英综合(2014年9期)2014-09-12 14:45:46
