
化州不同林龄人工林土壤养分及林下植物功能群特征研究
2021-05-24 12:34曾庆圣董辉宋霞王丹枫
热带林业 2021年1期
曾庆圣,董辉,宋霞,王丹枫
1.广东省森林资源保育中心,广东广州510650;
2.中国科学院华南植物园,广东广州510650;
3.广东化州国营丽岗林场,广东化州525100
桉树(Eucalyptus robustaSmith)是桃金娘科桉属树种的总称,其具有速生、高产、优质的特点,是中国南方速生丰产林的战略性树种。大面积的种植桉树导致了土壤地力衰退、林分产量下降的现象,维持桉树人工林长期稳定的可持续发展已成为国内外关注的焦点[1]。目前,有关桉树研究主要集中在桉树林土壤养分[2-3]、生物量和碳储量[4-5]及桉树水文效应[6]等,而桉树人工林生物多样性一直是个颇具争议的生态学问题。以往研究表明,桉树林多样性指数显著低于次生林,一般不高于乡土树种人工林,但总体上好于其他外来树种人工林。连栽桉树人工林植物多样性随连栽代次的增加而减少,物种生态优势度增加[1]。在中国,桉树人工林占据重要地位,2019年桉树人工林面积已突破546万hm2,在国家木材安全保障、应对气候变化等过程中发挥了重要作用[7]。湿地松(Pinus elliottii)原产美国东南部,具有适应性强、生长快、成活率高、材质好等特点,是一种重要的脂材双用松科树种。现阶段,中国对湿地松造林培育的研究主要集中在育苗、间伐栽培试验、林分密度、材积模型和病虫害防治等方面[8]。在桉树、湿地松人工林的传统经营中,人们往往只注重营林措施对林木和林分木材产量的作用,极少关注营林措施对林下植被、植物多样性、植物功能群、土壤理化性质等的影响,结果导致桉树、湿地松人工林生态服务功能减弱、木材生产与生态服务失衡[9],从而引发地力衰退、生物多样性下降、生态系统稳定性弱化等一系列争议[10-11]。
物种多样性与生态系统功能的关系一直是当代生态学研究的重大科学问题之一。物种多样性越丰富人工林越稳定,群落生产力也越高,也越能促进生态系统功能的发挥。林下植被是人工林生态系统中非常重要的组成部分,在维持人工林生态系统稳定性、调控地上地下养分循环及能量转化、促进生物多样性发展等方面发挥着不可替代的作用[12-13]。以往对林下植被多样性的研究主要集中在灌草层的种类特征及数量统计分析方面,其结果不能很好地解释林下植被物种间的复杂性及其在森林生态系统中的运作机制。已有研究表明,植物功能群的组成是影响群落生产力及其稳定性的主要因子[14-15]。植物功能群能更好地解释具有相似适应特征的植物物种在生态系统中的作用,反映植被随环境的动态变化,有效解释生态系统的构建机制,是评价生态系统结构和功能变化的重要依据[16-17]。
该研究以桉树纯林(PureEucalyptusplantation,PE)、湿地松纯林(PurePinus elliottiiplantation,PPE)、混交林(Mixed plantation,MP)的3种林龄阶段林分为对象,探究不同林龄林地土壤因子和林下植物丰富度及重要值,明确林下植物功能群与环境因子之间的关系,以期为化州人工林可持续经营管理提供科学依据。
1 材料与方法
1.1 研究区域及各林分类型概况
化州市属南亚热带季风气候,夏长冬短,气候温和,阳光充足。年平均日照时数1870.1h,年平均温度在22.2℃~23.9℃之间。冬季寒潮入侵,偶有严寒。化州市降水充沛,年平均降雨量1890mm[18]。地带性土壤类型为砖红壤。林内地下水位较高,土壤略呈黑色,属粘性土。
各林分类型调查样地分别位于化州市国营丽岗林场和平定林场,样地的地形地貌、海拔、母岩、土壤类型等相同或相近,坡度都在15°以下;样地造林密度和造林抚育措施基本一致,株行距一般为2m×3m,造林后前3年进行水平带状砍杂除草,种植穴复土施肥等抚育措施,施肥量保持一致,头年抚育2次,后2年抚育3次,3年后基本不再抚育管理。林龄分别为5a、10a、15a,此外,在丽岗林场场部附近的卜岭村选取100a罗汉松(Podocarpus macrophyllus)40a窿缘桉(Eucalyptus exserta)后自然演替的风水林作为人工混交林的成熟林。
1.2 样方设置与林下植物群落调查
2020年9月,用相邻格子法,在不同林龄PE、PPE、MP内各设置3个20m×60m的调查样方,分成12个10m×10m的小样方,对样带内胸径(DBH)≥1cm的个体进行每木检尺,记录其种名、树高、胸径和冠幅等,在每个小样方内随机设立1个5m×5m的灌木层样方和1个2m×2m的草本层样方,调查DBH<1cm、树高大于50cm的灌木和草本层样方中的所有草本植物,记录其种类、高度和冠幅(或盖度),同时记录样带内出现的层间植物及其盖度,对样带内上层优势树种的所有林下个体均调查其基径、高度和冠幅。
1.3 土壤样品采集与测定
在每个样方(20m×60m)中随机选择6个采样点,采用环刀法测定0cm~20cm土层的土壤容重。用内径为8.5cm的不锈钢土钻钻取深度为0cm~20cm土壤样品,去除植物根系及石砾后混合均匀,过2mm孔径筛,将样品分为2份,一份风干用于测定土壤理化性质,另一份保存于4℃冰箱用于土壤铵态氮、硝态氮的测定。
土壤容重(Soil bulk density,SBD)采用环刀法测定,土壤含水量(Soil moisture content,SMC)采用烘干法测定,土壤pH采用pH计测定(土:水=1:2.5,w/v),有机碳(Soil organic carbon,SOC)采用K2Cr2O7-H2SO4外加热法测定,全氮(Total nitrogen,TN)和有效氮(Available nitrogen,AN)(铵态氮和硝态氮)采用连续流动分析仪(AA3)测定,全磷(Total phosphorus,TP)采用H2SO4-HCLO4-钼锑抗比色法测定,有效磷(Available phosphorus,AP)采用双酸(HCL-H2SO4)浸提-钼锑抗比色法测定。
1.4 数据分析
参考相关文献[16]的方法,结合植物生态型概念,将林下植被分成3类植物功能群,分别为阳性植物功能群(Positive plant functional group,PFG)、阴性植物功能群(Negative plant functional group,NFG)、耐荫性植物功能群(Shade-tolerant plant functional group,SFG)。阳性植物功能群即群落中对光照要求比较高,只有在足够光照条件下才能正常生长的植物群体;阴性植物功能群是指对光照要求远比阳性植物功能群低,可低于全光照的2%;耐荫植物功能群是对光照具有较广的适应能力,对光的需求介于上述二者之间。采用每种林下植物物种的重要值(importance value,IV)来表征其在相应群落中的地位和优势大小,计算方法如下:

其中,Ra为相对多度,即样方中某个种的个体数占全部种的个体总数的百分比;Rp为相对频度,即样方中某个种出现的频度占全部种的频度的百分比;Rc为相对盖度,即样方中某个种的盖度占全部种的盖度的百分比。
数据在SPSS 24.0中分析,采用单因素方差分析(one-way ANOVA)检验不同林分间土壤养分、功能群物种丰富度、重要值的差异显著性,采用LSD法进行多重比较,显著性水平设置为P<0.05。采用多变量的相关性分析,检验土壤因子与林分林下功能群重要值间的相关性。
2 结果与分析
2.1 土壤C、N、P及其化学计量比
土壤风干含水量最大的为林分PE(7.50±0.34%),显著高于PPE和MP,PPE的风干含水量也显著小于MP的。PPE的土壤容重介于PE和MP之间,且与后二者无显著性差异;而MP的土壤容重(1.11±0.01g·cm-3)比PE(1.01±0.01g·cm-3)高9.9%,二者差异显著。PE的土壤全氮、全磷、全钾含量均显著高于PPE和MP,其中PE的土壤全氮含量(1483.16±81.92mg/kg)分 别比PPE(953.42±47.28mg/kg)和MP(1037.73±57.47mg/kg)高55.56%和42.92%;PE的土壤全磷含量(210.22±8.67mg/kg)也显著高于PPE(159.42±7.18mg/kg)和MP(184.51±4.17mg/kg);土壤全钾含量在3种林分间也存在显著差异,其中最大的还是PE(13.93±0.86g/kg),比PPE(12.59±0.81g/kg)、MP(11.33±0.28g/kg)分别高10.64%、22.94%(表1)。

表1 不同林分的土壤理化特性(0cm~20cm)Tab.1 Soil Properties(0cm~20cm)in Different Stands
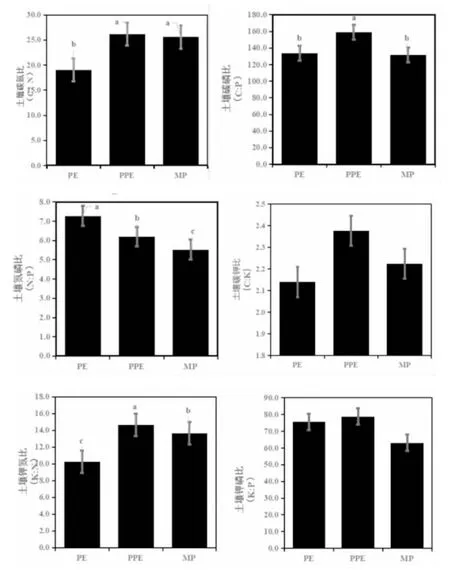
图1 不同林分土壤C、N、P、K含量化学计量比Fig.1 Soil C,N,Pand K Concentrations Atoichiometric Ratio in Different Stands
由图1可知,不同林分的土壤有机碳无显著差异(P>0.05),其含量大小排序为PE>MP>PPE。在不同林分土壤元素含量化学计量比方面,3种不同林分土壤C:N、C:P、K:N均存在显著性差异,且3种都是PPE的值最大,其次为MP,最后是PE。但在N:P方面则是PE最高且与PPE和MP有着显著差异。单因素方差分析显示,不同林分土壤碳钾比和土壤钾磷比都无显著差异,但比值最大的还是PPE。
2.2 不同林龄人工林林下植物功能群物种丰富度的影响
不同林龄林分需要不同的经营模式,不同经营模式对林分林下植物物种组成及其重要值具有显著影响。该研究调查共记录150种维管植物,分属58科111属,其中:蕨类植物7科8属12种;裸子植物4科4属4种;双子叶植物42科89属122种;单子叶植物5科10属12种。MP样地林分的物种最多,共有维管束植物36科54属70种,其中:蕨类植物5科5属5种;裸子植物2科2属2种;双子叶植物28科46属62种;单子叶植物1科1属1种。其次PE林分物种共有66种,PPE林分物种最少,共有52种。从表2可以看出,将调查的所有人工林林下植被分为3个植物功能群,不同林分林下植物群落的优势物种组成发生了显著的变化。

表2 样地林下植被组成Tab.2 Composition of Understory Vegetation in the Plot
由图2及单因素方差分析结果显示,不同林分类型不同林龄人工林林下耐荫性植物丰富度存在显著差异(P<0.05),大于或等于15a的林分的耐阴性植物功能群丰富度显著大于5a和10a的,10a的林分耐阴性植物功能群丰富度也显著大于5a的,这说明人工林群落在0a~40a间耐荫性植物功能群的丰富度一直处于快速增长期,且混交林增长速度最快。在不同林分类型的不同林龄林分中,阳性、耐荫性、阴性植物功能群的物种丰富度也存在显著性差异(P<0.05),总体呈现耐荫性植物功能群物种丰富度>阳性植物功能群物种丰富度>阴性植物功能群。不同林分各功能群物种丰富度也呈现相同的规律,耐荫性、阳性植物功能群物种丰富度存在显著差异(P<0.05),MP显著大于PE和PPE,阴性植物功能群物种丰富度间无显著差异,这一定程度上反映的该地区人工林林下阴性植物功能群物种丰富度相似性。
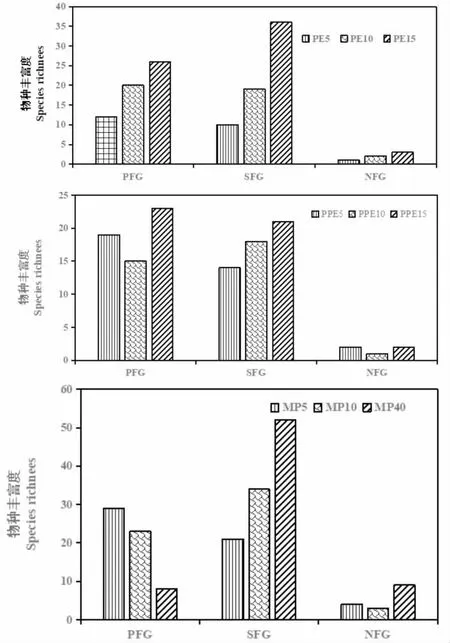
图2 不同林分林下植物功能群的物种丰富度Fig.2 Species Richness of the Understory Plant Communities in Different Stands
2.3 不同林龄人工林林下植物功能群重要值情况
不同林龄林分间阳性植物功能群的重要值存在显著差异(P<0.05)。PE与MP5a林分阳性植物功能群的重要值显著高于10a和15a(40a)的,但PPE林分为10a>5a>15a。不同的林龄林分耐荫性、阴性植物功能群的重要值无显著性差异(P>0.05)。不同林分阳性、阴性植物功能群的重要值间都存在显著差异性(P<0.05)(图3),PPE和PE阳性植物功能群重要值极显著大于MP阳性植物功能群,这说明单一树种林分结构较为单一,容易受环境条件干扰。不同林分耐荫性植物功能群重要值间无显著差异,但物种丰富度较大的混交林各林龄阶段耐荫性植物功能群重要值变幅最小,群落最稳定。

图3 不同林龄人工林林下植物功能群重要值Fig.3 Important Values of Functional Groups of Understory Plants in Artificial Forests of Different Ages
2.4 土壤因子对不同林龄人工林林下植物功能群的影响
相关性分析结果表明(表3),土壤全氮、土壤全钾和土壤钾氮比与3种林分林下阳性植物功能群重要值都呈极显著的负相关关系,相关系数分别为-0.966、-0.926和-0.911;土壤钾氮比还与阴性植物功能群重要值间存在极显著的负相关关系,相关系数为-0.985,其余土壤因子和所有林下各植物功能群的物种重要值的相关性都不显著。耐荫性植物功能群重要值与土壤因子之间没有一个具有显著相关性的指标,这可能是因为耐荫植物在群落中相对于阳性植物和阴性植物,其受土壤因子影响较小,较为稳定。由表3还可看出土壤因子与植物功能群重要值具有显著相关性的都包含钾和氮元素,说明化州人工林植物功能群构建可能与土壤中氮元素和钾元素有一定的相关性。

表3 土壤因子与不同林龄人工林林下植物功能群物种重要值的相关性分析Tab.3 Correlation Analysis Between Soil Factors and Important Values of Understory Plantfunctional Groups in Artificial Forests of Different Ages
3 结论
化州桉树林土壤全氮、全磷、全钾都显著高于湿地松林和阔叶林,土壤全氮、土壤全钾和土壤钾氮比与植物功能群重要值有着显著的相关性,土壤全氮、土壤全钾和土壤钾氮比对林下植物群变异都发挥着重要作用。在不同林分类型的不同林龄林分中,阳性、耐荫性、阴性植物功能群的物种丰富度存在显著性差异,总体呈现耐荫性植物功能群物种丰富度>阳性植物功能群物种丰富度>阴性植物功能群物种丰富度。不同林分耐荫性、阳性植物功能群物种丰富度和重要值也存在显著差异,MP耐荫性、阳性植物功能群物种丰富度显著大于PE和PPE,但MP耐荫性、阳性植物功能群物种重要值显著小于PE和PPE。不同林分类型人工林,随着林龄或混交树种增加,阳性植物功能群的丰富度增加,重要值降低,群落逐渐向成熟阶段发展。
4 讨论
物种多样性是物种丰富度和分布均匀性的综合反映,体现了林分的结构类型与组织水平以及发展阶段、稳定程度和生境差异等情况[19]。该研究发现随着林龄的增长,耐荫性植物功能群的物种丰富度在人工林生长初期丰富度较低,之后逐渐升高,因为该研究只选取了5a、10a、15a(40a)的桉树林、湿地松林、阔叶林,所以其物种丰富度没有呈现先升高后逐渐下降的趋势[20]。在幼龄林阶段,林分郁闭程度较低,林下有充足的光照条件,为喜光物种提供良好的生存条件,不利于耐荫性植物生长。到中龄林、成熟林阶段,林下生境的光照条件和温湿度等环境因子发生改变,耐荫性物种逐渐进入,物种多样性也相应增加;随着林龄增加,林分郁闭度增加,不利于喜光植物的生存,林下种类对生存空间和养分的竞争加剧,不能适应环境的物种则被淘汰,尤其是一些阳性植物功能群植物减少[21-22]。其次在该研究的3个林型中,发现随着林龄增加,林分郁闭度增加,其林下阴性植物功能群的丰富度不同林龄林分之间无显著差异,耐荫的蕨类植物林分生长初期快速入侵定居,随着林分郁闭度增加,外界环境更加适于其生长,最终成为占据林下优势的植物功能群。
植物功能群是基于植物生理形态及生活史或其他与生态系统功能相联系的功能属性的差异划分的类群[23]。在森林生态系统中,随着林龄的增加,演替也在不断进行,林下植物功能群组成也在不断变化[24]。该研究发现,在郁闭度小的5a的3种研究林分中,林下阴性植物功能群和阳性植物功能群占据了主要优势,各功能群的重要值排序为NFG>PFG>SFG;10a、15a林分郁闭度居中的3种林分中,林下各个功能群的重要值排序为PFG>NFG>SFG,还是阳性植物功能群和阴性植物功能群占据主要优势;而在林分郁闭度最高的40a林分中,林下功能群重要值的排序SFG>PFG>NFG,占据主要优势变为耐荫性植物功能群。这一定程度上说明随着林龄增加,阳性物种功能群和阴性物种功能群在群落中的优势逐渐减小,耐荫性物种植物功能群优势逐渐变大。
森林生态系统中的土壤既是物质循环和能量转换的重要场所,也是植物群落更新演替过程的载体[25-26],还是主要影响植物群落分布的环境因素之一。该研究发现,林下物种丰富度最高的混交林中,其土壤全氮含量显著高于物种丰富度较低的桉树纯林和湿地松纯林。可见,混交林不仅能充分利用地上地下空间及资源[27],还能提高土壤的固氮能力。土壤肥力、水分、质地等对植物群落的分布影响重大[28],如桉树人工林去除林下植物功能群会导致土壤养分有效性显著降低,呈现富集状态,说明植物功能群的性状与土壤肥力有效性之间存在密切且复杂的依存关系。该研究结果表明土壤全氮、土壤全钾和土壤钾氮比是阳性植物功能群物种重要值的主要影响因子,阴性植物功能群在群落中的重要性也主要受土壤土壤钾氮比影响,耐荫性植物功能群在群落中的重要值与土壤因子呈现一定的负相关关系。然而影响植被生长与分布的环境因素错综复杂,有学者发现土壤磷含量与植物群落物种多样性存在明显的相关性。该研究结果与其他研究有所异同[24-26],一定程度上反映了森林生态系统不同的组成结构与时空尺度的异质性。此外,在该研究中发现其他土壤因子对林下植物功能群的影响并不显著,其原因可能与这些因子在各个林分中差异程度较小有关。
猜你喜欢
防护林科技(2022年5期)2022-08-22
农村科学实验(2021年27期)2021-11-12
防护林科技(2020年10期)2020-12-22
四川林业科技(2020年1期)2020-08-31
防护林科技(2020年6期)2020-08-12
绿色科技(2019年6期)2019-04-12
绿色科技(2019年6期)2019-04-12
儿童故事画报·发现号趣味百科(2019年9期)2019-02-02
环球时报(2019-01-03)2019-01-03
百科知识(2016年22期)2016-12-24
