
不同经营强度下毛竹丛枝菌根共生体对氨氧化微生物的影响*
2021-05-22 07:22:44葛江飞杨为中高雄飞孙思哲陈俊辉梁辰飞徐秋芳邬奇峰
土壤学报 2021年2期
葛江飞,杨为中,高雄飞,孙思哲,陈俊辉,梁辰飞,徐秋芳,邬奇峰,秦 华†
(1. 浙江省森林生态系统碳循环实验室,浙江农林大学,浙江临安 311300;2. 浙江农林大学环境与资源学院,浙江临安 3113003;3. 浙江省临安区农业技术推广中心,浙江临安311300)
毛竹(Phyllostachys pubescens)生长周期短、用途广泛、经济价值高,已经成为我国南方地区尤其是山区农民的主要经济来源之一。近二十余年,为追求更高的经济效益,大量毛竹林由粗放经营模式逐渐转变为集约经营,主要措施为大量施用氮肥、深翻土壤以及林下清除灌木和杂草等,从而获得更高的竹材和竹笋产量。集约经营模式虽然增加了农户的经济收益,但是大量施用氮肥也带来了土壤酸化、微生物活性降低以及水体富营养化等一系列环境问题[1]。此外,已有学者研究发现毛竹林施肥、翻耕等集约经营措施也会导致土壤某些功能菌群如固氮菌和氨氧化微生物的群落结构发生显著改变[2],对土壤氮循环产生不利影响并且会加剧温室气体排放[3]。
丛枝菌根(Arbuscular mycorrhiza,AM)真菌是自然界分布最广泛的一类菌根真菌,能侵染超过80%的高等陆生植物根系,帮助宿主植物提高水分及养分利用效率[4]。由于氨氧化微生物是化能自养微生物,早期研究表明,AM 真菌对土壤氨氧化微生物没有影响[5]。但近年来有研究发现,AM 真菌可能通过与氨氧化微生物竞争土壤中的有效氮(NH4+)从而影响氨氧化微生物的丰度及群落[6-8]。Teutscherova 等[7]的室内培养实验研究发现接种AM真菌处理的土壤中氨氧化细菌(Ammonia-oxidizing bacteria,AOB)的丰度显著上升,其他许多研究结果也表明AM 真菌对AOB 丰度的影响是积极的[9-11],并且AOB 的群落结构也会因为AM 真菌的存在发生显著的改变[12]。但也有一些研究发现AM 真菌的存在会对氨氧化微生物产生负面的影响[6,13-14],并且这些负面的影响大多数是在土壤NH3较低的情况下发生的,如土壤肥力低或者土壤pH 较低土壤中的氨多以NH4+的形式存在,不利于被氨氧化古菌(Ammonia-oxidizing archaea,AOA)或者AOB 利用。
虽然在菌根共生过程中AM 真菌与氨氧化微生物之间的养分竞争假设已成为目前较为流行的观点,但是这些研究未将AM 真菌与根系的效果分开,因此无法准确评估AM 真菌与根系对氨氧化微生物群落演变的贡献。此外,在不同的毛竹林经营强度下,菌根对氨氧化微生物影响如何目前尚未有明确的结论。因此,本研究利用原位微宇宙试验,研究毛竹林生态系统中丛枝菌根及菌丝对氨氧化微生物群落的影响及其对毛竹经营强度的响应机制,为毛竹林科学合理施用氮肥,减缓全球气候变化提供理论依据。
1 材料与方法
1.1 试验地概况
试验地点位于浙江省杭州市临安区青山镇(119°42′ E,30°14′ N),海拔50 m,属亚热带季风气候,温暖湿润,光照充足,年降水量1 420 mm,年均气温约15.6 ℃,年均无霜期约230 d,地形为低山丘陵,土壤类型为红壤。选取集约经营毛竹林作为研究对象,同时选取附近立地条件相似的粗放经营毛竹林作为对照。集约经营毛竹林的经营措施主要是隔年采伐老竹留养新竹,去除林下灌木、杂草,并对毛竹进行钩梢。每年6 月中下旬采用撒施的方式对毛竹林进行施肥,然后进行深翻,化肥用量为:尿素(CO(NH2)2)450 kg·hm–2、过磷酸钙(Ca(H2PO4)2)450 kg·hm–2、氯化钾(KCl)150 kg·hm–2。粗放经营毛竹林不施肥,隔年采伐老竹留养新竹,林下植被主要有连蕊茶(Camellia cuspidate)、狗脊蕨(Woodwardia japonica)、兔儿伞(Syneilesis aconitifolia)、络石(Trachelospermum jasminoides)、白花败酱草(Patriniavillosa)、宝铎草(Disporum sessile)等。
1.2 试验设计
于2017 年6 月分别在集约经营和粗放经营的毛竹林样地各建立4 个10 m×10 m 的样方,各样方之间距离均大于30 m,共计8 个样方。在每个样方内设置2 个不同的微宇宙系统,共计16 个。微宇宙系统由PVC 筒(直径16 cm,高度20 cm)和尼龙筛网制成,PVC 筒壁上开有8 个孔(4 个6 cm 孔径和4 个3 cm 孔径),底部开有4 个孔(6 cm 孔径),所有的孔均安装有不同孔径的尼龙筛网以区分不同菌根分室处理,其中2 mm 孔径的筛网可以使得毛竹细根及AM 真菌菌丝均能通过(+细根+AM 真菌),设为菌根室(RA)处理;35 μm 孔径的筛网仅允许AM 真菌菌丝通过(-细根+AM 真菌),设为菌丝室(AH)处理。分层取出样地0~20 cm 土壤,去除明显的植物根系及石块,放置好PVC 筒后,再尽快按照原来顺序将土壤分层放入筒内。在微宇宙系统布置完成后原地培养3 个月,用土钻采集微宇宙系统内土壤带回实验室进行分析。鲜土样混合后过2 mm筛,一部分土壤立即进行DNA 提取及硝化势的测定,另一部分风干进行理化性质分析。
1.3 土壤理化性质及硝化势测定
土壤理化性质参照鲁如坤等[15]方法:土壤有机质采用重铬酸钾-硫酸外加热法测定;碱解氮采用碱解扩散法测定;有效磷采用盐酸-氟化铵溶液浸提,钼锑抗比色法测定;速效钾采用醋酸铵提取,火焰分光光度计测定;土壤pH 采用水土比2.5∶1,玻璃电极法测定。
土壤硝化势的测定采用好气培养法[16]并作略微修改。称取 10 g 新鲜土样置于锥形瓶中,加入(NH4)2SO4调节NH4+-N 浓度到400 mg·kg–1,以干土计算,将含水量调节至田间最大持水量的60%,同时以不加(NH4)2SO4溶液的土壤作为对照。将锥形瓶放于25 ℃的培养箱中避光培养48 h,培养结束后,用50 mL 1 mol·L–1KCl 溶液浸提并过滤,流动分析仪测定NO3--N 含量。
1.4 土壤DNA 提取及定量PCR
土壤总 DNA 的提取采用 MoBio PowerSoil®DNA Isolation Kit 试剂盒(MoBio Laboratories,美国),称取0.5 g 冷冻干燥土壤样品,按照试剂盒说明提取DNA。提取的DNA 片段大小经1%(m/v)的琼脂糖凝胶电泳检测,并用微量分光光度计(NanoDrop ND-1000,Thermo Scientific,美国)进行浓度测定,保存于–40 ℃用于后续分析。
使用CFX96TMReal-Time System(Bio-Rad)仪器进行氨氧化古菌和氨氧化细菌 amoA 功能基因实时定量分析,以确定样品的基因拷贝数。AOA和AOB amoA 基因的PCR 扩增采用的引物对分别是Arch-amoAF:5′-A ATGGTCTGGCTTAGACG-3′/Arch-amoAR:5′-GCGGCCATCCATCTGATTGT-3′和amoA-1F:5′-GGGGTTTCTACTGGTGGT-3′/amoA-2R:5′-CCCCTCGGGAAAGCCTTCTTC- 3′[17],每个样品3 个重复,反应体系为:2×SYBR Premix Ex Taq 10 μL,50 μmol·L–1上游引物和下游引物各0.2 μL,DNA 模板1.0 μL,无菌水补足至20 μL。AOA 和AOB 的扩增程序见参考文献[18]。
1.5 高通量测序及数据处理
采用带barcode 的AOA 和AOB 特异性引物,利用Illumina Miseq 平台进行高通量测序,测序服务委托杭州联川生物技术有限公司完成。用QIIME2将Miseq 测序完成后得到原始的下机数据进行处理,利用overlap 将双端数据进行拼接,并进行质控、嵌合体过滤,获得高质量的有效数据。对最终获得的有效数据采用Vsearch2.3.4 算法按照97%的相似度聚类,为了降低假阳性率,过滤掉singleton 序列,获得最终的OTU 丰度及代表序列,进一步进行多样性分析、物种分类注释和差异分析等。
1.6 数据分析
采用SPSS19.0 对土壤理化性质、硝化势、AOA和AOB amoA 基因拷贝数、α 多样性指数以及优势属相对丰度均进行单因素方差分析(One-way analysis of variance)和双因素方差分析(Two-way analysis of variance)比较各处理之间的差异显著性(P<0.05)。对土壤理化性质与AOA 和AOB 的amoA基因拷贝数进行Pearson 相关性分析。采用R 语言Vegan 包对不同处理土壤AOA 和AOB 群落结构进行非度量多维尺度转换排序分析(Non-metric multidimensional scaling,NMDS),并通过相似性检验(Analysis of similarities,ANOSIM)检验不同群落之间的差异显著性(P<0.05)。为了进一步了解经营强度和不同分室处理对氨氧化微生物种群之间相互作用的影响,利用MENA 平台对OTU 丰度进行网络分析。
2 结 果
2.1 土壤理化性质及硝化势
无论是集约经营还是粗放经营毛竹林,AH 处理土壤pH 均显著低于RA 处理(P<0.05)。集约经营毛竹林AH 处理土壤碱解氮也显著低于RA 处理(P<0.05),有机质及其他养分含量均未发生显著变化(表1)。与不同分室处理相比,不同经营强度对土壤养分含量的影响更为明显。集约经营毛竹林RA处理土壤有机质、pH 显著低于粗放经营毛竹林,而碱解氮含量显著高于粗放经营毛竹林土壤。同时,集约经营毛竹林AH 处理土壤有效磷和速效钾含量显著高于粗放经营毛竹林,而 pH 显著降低(P<0.05)。双因素方差分析结果表明,经营强度和分室处理均显著影响土壤pH 和碱解氮含量,但两者没有显著的协同效应(表1)。
在同一类型毛竹林中,不同分室处理之间土壤硝化势均没有显著差异(表1),而集约经营毛竹林RA 处理土壤硝化势显著高于粗放经营毛竹林(P<0.01)。双因素方差分析结果表明,经营强度对硝化势影响显著(P<0.01),不同分室处理对硝化势影响不显著,但两者对土壤硝化势具有显著的协同作用(P<0.05)。
2.2 氨氧化微生物amoA 基因丰度
荧光定量PCR 结果显示,无论是集约经营还是粗放经营,不同分室处理之间毛竹林AOA 的基因丰度均无显著差异(图1)。而在同一分室处理条件下,集约经营毛竹林RA 和AH 的AOA 基因丰度均显著高于粗放经营(P<0.05)。双因素方差分析结果表明,经营强度对AOA 基因丰度影响显著(P<0.01)。
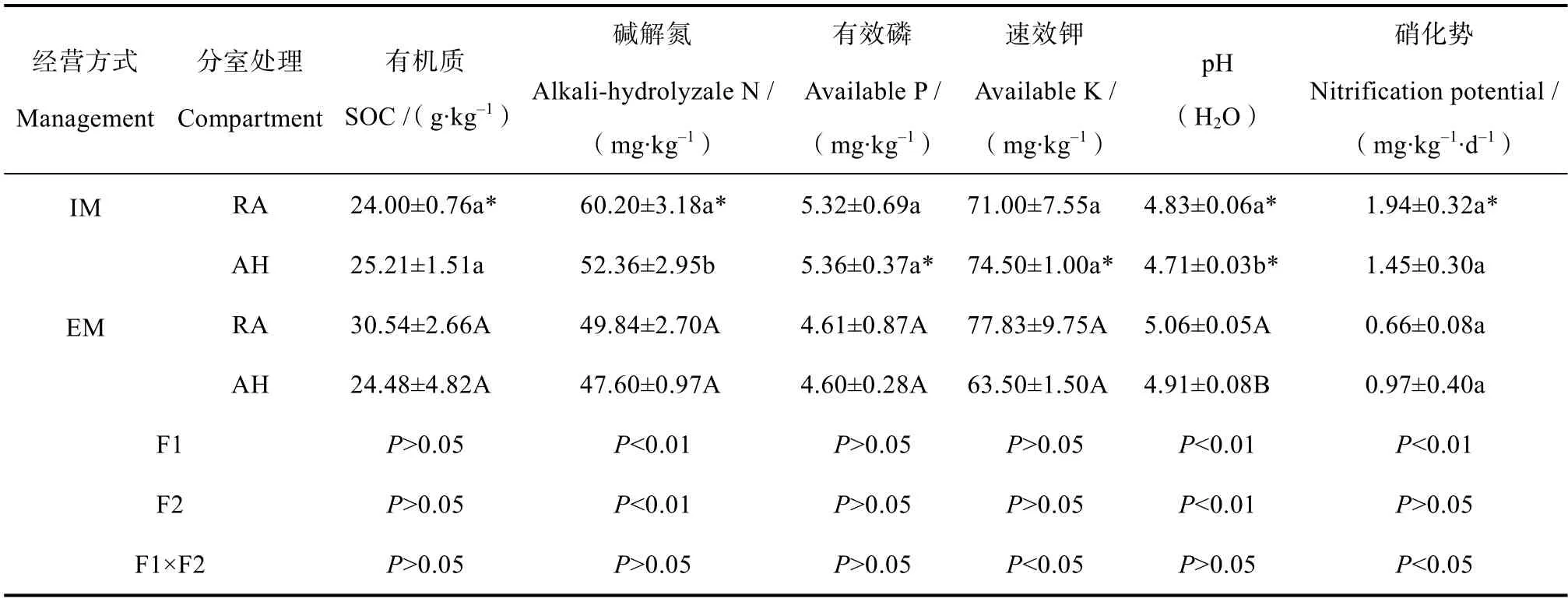
表1 毛竹林土壤理化性质及硝化势Table1 Soil physic-chemical properties and nitrification potentials of the Moso bamboo forest relative to management intensity(mean±SD,n=3)
无论是RA 处理还是AH 处理,AOB 在不同经营强度毛竹林之间均无显著差异(图1)。在同一经营强度下,无论是集约经营还是粗放经营,RA 处理土壤AOB 基因丰度均显著高于AH 处理(P<0.05)。双因素方差分析结果表明,经营强度和分室处理均对土壤AOB 基因丰度影响显著(P<0.05),两者交互作用也达到显著水平(P<0.05)。
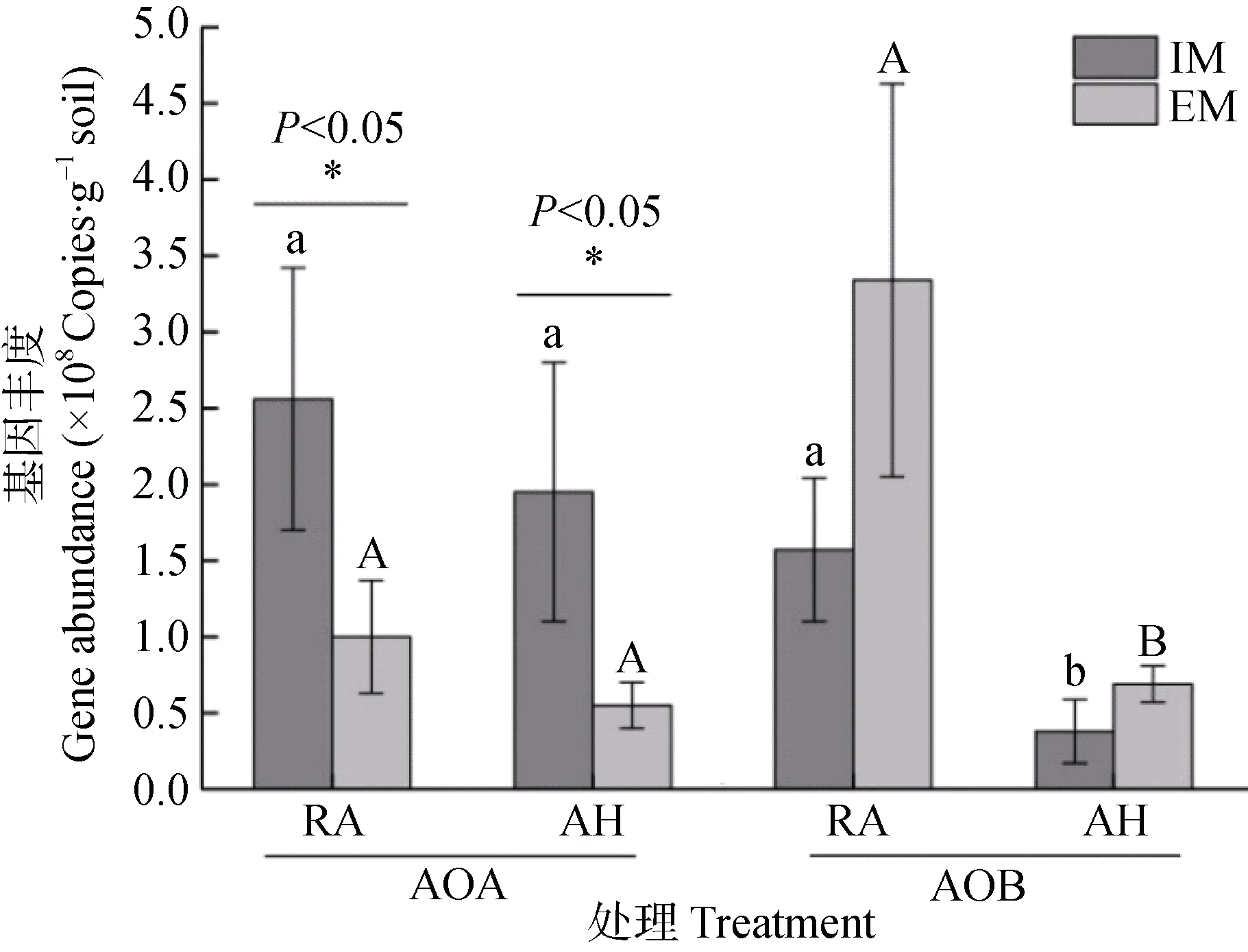
图1 毛竹林土壤AOA 及AOB 基因丰度Fig. 1 Abundance of AOA and AOB in Moso bamboo forest soils relative to management intensity
2.3 氨氧化微生物amoA 基因丰度与土壤理化因子及硝化势的相关性
对氨氧化微生物丰度与土壤理化因子及硝化势的Pearson 相关性检验结果表明,AOA 丰度与硝化势、碱解氮显著正相关,与土壤pH 显著负相关,而AOB 丰度则与土壤有机质和pH 显著正相关(表2)。
2.4 氨氧化微生物的群落结构
集约经营毛竹林中,AH 处理AOA 的物种数显著低于RA 处理,而粗放经营毛竹林则相反,AH 处理AOA 的物种数及α 多样性指数均显著高于RA 处理(P<0.05)。两种经营方式相比,粗放经营毛竹林AH 处理的物种数及α 多样性指数均显著高于集约经营毛竹林(表3)。双因素方差分析显示经营强度对AOA 的物种数及α 多样性指数影响显著,不同分室处理无显著影响,但是两种因素对物种数和Chao1 指数具有极显著的交互作用(P<0.01)。与AOA 相比,AOB 物种数及α 多样性指数在不同经营强度和分室处理之间均无显著差异(表3)。双因素方差分析结果表明,仅经营强度对 AOB 群落Chao1 指数影响显著(P<0.05),经营强度及分室处理对 AOB 群落香农指数具有显著的交互作用(P<0.05)。
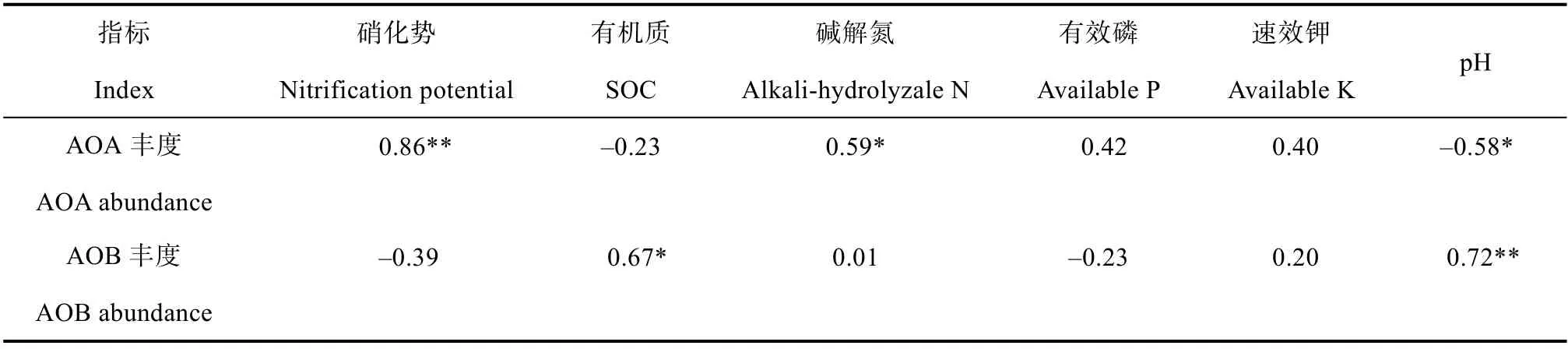
表2 氨氧化微生物基因拷贝数与土壤理化及硝化势相关性分析Table2 Correlation analysis of abundance of ammonia oxidizers with soil properties and soil nitrification potential
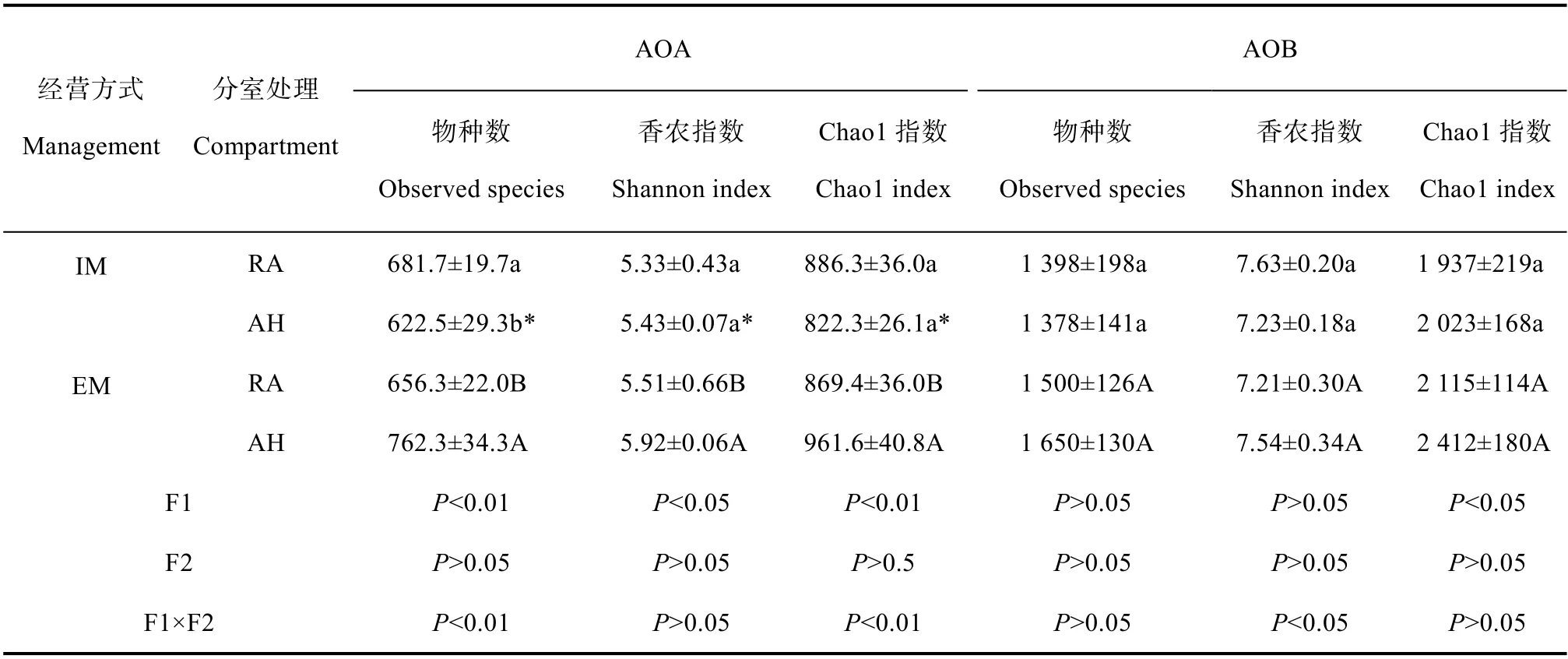
表3 毛竹林土壤AOA、AOB 群落多样性指数Table3 Diversity indices of the AOA and AOB communities in the Moso bamboo forest soils
ANOSIM 分析结果显示,无论是经营强度还是不同分室处理均未对AOA 和AOB 的群落结构产生显著影响,但在集约经营毛竹林土壤中,RA 和AH处理的AOA 群落有明显区分。对AOB 而言,不同经营强度毛竹林AH 处理之间区分明显(图2)。
2.5 氨氧化微生物分子生态网络特征
基于两种经营强度毛竹林的分组方式,对AOA和AOB 的高通量测序数据进行网络分析。AOA 的网络分析结果显示,粗放经营毛竹林网络中节点数高于集约经营,但是连接数和正相关比例均低于集约经营(表4)。而AOB 的网络分析结果则显示,粗放经营毛竹林网络中节点数和连接数均高于集约经营,但正相关比例低于集约经营毛竹林(表4)。不论AOA 还是AOB,网络中正相关的连接数均表现出集约经营高于粗放经营的趋势。将网络节点与环境因子进行曼特尔相关性分析(Mantel test)发现,在每个分类水平环境因子并未对网络中的不同物种产生显著影响。
3 讨 论

图2 AOA、AOB 群落组成NMDS 排序图Fig. 2 NMDS ordination plots of the community compositions of ammonia-oxidizing bacteria and archaea
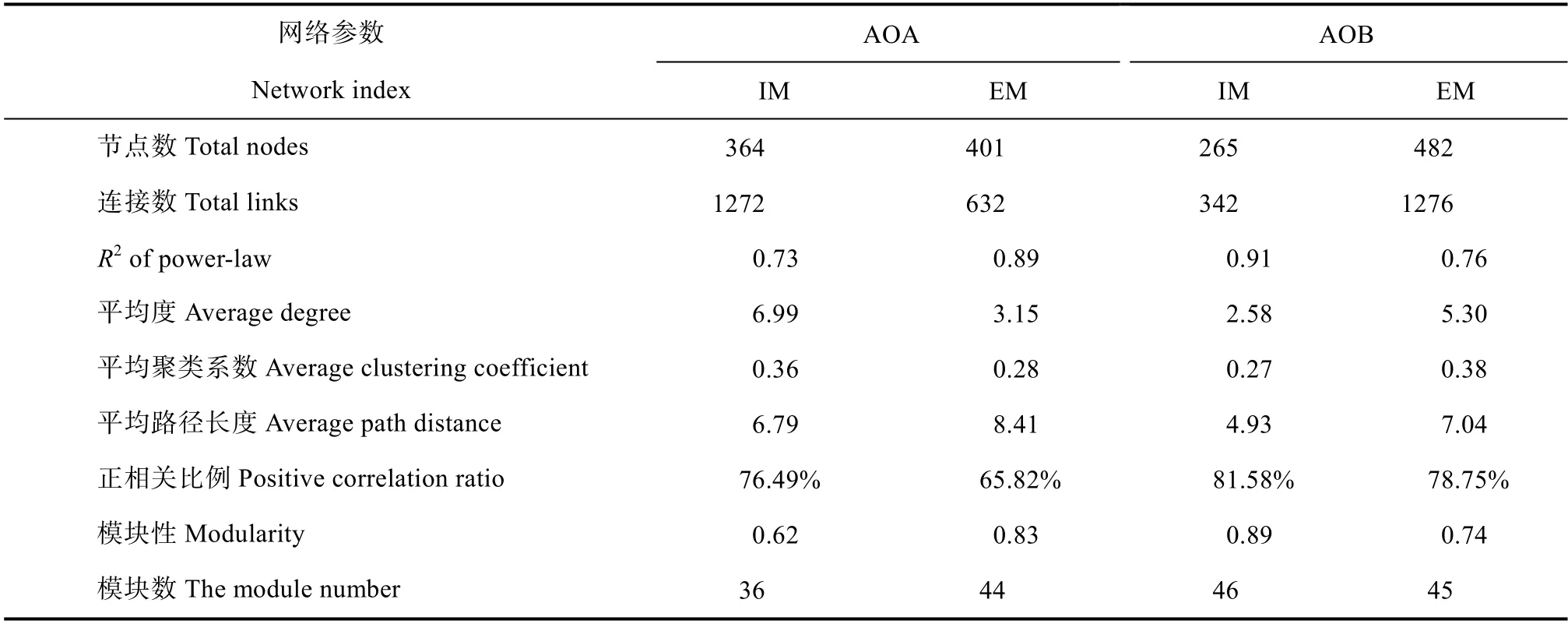
表4 毛竹林土壤AOA、AOB 分子生态网络特征Table4 The properties of the molecular ecological networks of the AOA and AOB communities in the Moso bamboo forest soils
微宇宙试验结果表明无论是集约经营还是粗放经营毛竹林,AH 处理土壤 pH 均显著低于 RA(P<0.05),这与Veresoglou 等[13]的研究结果一致,主要是由于AM 菌丝体分泌的有机酸可降低土壤pH。其他学者分别以烟草和沙丁旺为宿主植物研究AM 真菌对其土壤理化性质的影响也得到了相同的结果[19-20]。集约经营毛竹林AH 处理土壤碱解氮也显著低于RA 处理(P<0.05)。虽然在菌根系统中,植物根系、AM 真菌以及氨氧化微生物可能会竞争土壤中的有效氮素,但在有根系存在的菌根系统中,植物根系随时都在进行着更新和分解,产生氮素归还到土壤[21],而在无根系存在的菌丝室中,氮的来源补充远少于菌根室,菌丝吸收氮素导致土壤有效氮含量降低,这可能是导致RA 处理中土壤碱解氮高于AH 处理的主要原因。
荧光定量PCR 结果表明,毛竹林经营强度对AOA 基因丰度产生了显著的影响,对AOB 的影响不显著,导致这一结果的原因可能是因为pH 降低影响了AOA 的丰度,相关性分析也显示AOA 基因丰度与pH 显著相关。Nicol 等[22]发现,AOA 基因丰度随着土壤pH 的增加而降低 (在4.9~7.5 之间),即相较于AOB 而言AOA 在自然界中拥有更广泛的生态位,更适于在酸性土壤条件下生存并且在酸性土壤中发挥的作用更大[23]。本研究中集约经营毛竹林大量的施肥导致土壤pH 下降,为AOA 提供了一个较为适宜的生存条件。研究结果发现不同的分室处理对AOB 影响显著,不论是集约经营还是粗放经营,AH 处理AOB 的基因丰度均显著减少。相反,RA 处理根系和AM 真菌共存的菌根系统中AOB 丰度更高。相关性分析显示pH 极显著地影响了AOB的丰度,并且AH 处理土壤pH 变化规律与AOB 丰度一致。因此,AH 处理AOB 丰度均显著减少可能是因为AM 真菌菌丝通过降低pH 间接地影响了AOB,较低的pH 会降低AOB 底物的利用效率[24]。这一结果与Teutscherova 等[7]的盆栽结果一致,AM真菌与植物共生对AOA 的丰度没有显著的影响,但是却显著提高了AOB 的丰度。除此之外,在RA 处理中植物根系与AM 真菌共生也会使得土壤孔隙度增加,根系分泌物和菌丝分泌物的双重作用会增加土壤团聚体含量使得土壤更疏松[25]。鉴于AOB 为一类好氧氨氧化微生物,疏松的土壤环境更有利于AOB 的生存。虽然经营强度和分室处理分别对AOA和AOB 的丰度产生了显著的影响,但对AOA 和AOB 群落结构影响并不显著。Cavagnaro 等[5]在2007年对一有机农场的AOB 的研究结果也发现,AM 真菌并没有对其群落组成和结构产生显著的影响,其原因可能是因为研究时间相对较短导致差异性不显著。此外,AOA 和AOB 的群落结构对土壤理化因子的响应非常复杂,其群落结构的变化取决于多种因子的综合效应,这也是本研究中集约经营导致的土壤性质变化仅仅改变了AOA 和AOB 的基因丰度,而对其群落结构并未产生显著影响的可能原因。
近年来,分子生态网络已经被广泛应用于了解微生物种间交互多样性及其生态系统稳定性[26]。本研究中对网络节点数(Total nodes)、连接数(Total links)、平均度(Average degree)的对比分析均发现AOA 网络中集约经营连接数和平均度大于粗放经营,说明集约经营毛竹林AOA 分子生态网络中物种丰富程度、物种交互复杂性高于粗放经营。AOB网络中,粗放经营网络节点数、连接数、平均度三项指标的值均大于集约经营,说明粗放经营毛竹林AOB 生态网络的物种丰富度、物种交互复杂性优于集约经营,这也说明了AOA 和AOB 在土壤中具有不同的生态位,对环境的响应具有较大的差异。网络中连接的正负相关性可以用于推断微生物之间的相互关系,正相关代表生态位一致或共生关系,微生物共建生物膜或者交换代谢产物,负相关代表着竞争或捕食[26]。本研究构建的四个生态网络中,网络节点连接之间均以正相关为主,表明AOA 和AOB的微生物之间合作关系大于竞争关系,并且AOA和AOB 的生态网络均表现出集约经营的正相关关联大于粗放经营,表明集约经营毛竹林中AOA 和AOB 存在更多的共生关系,而粗放经营毛竹林中氨氧化微生物之间的竞争更为激烈,可能是由于土壤养分供应少于集约经营。集约经营中氨氧化微生物之间的互作和共生关系较粗放经营更加紧密,而粗放经营毛竹林中氨氧化微生物之间的竞争比集约经营激烈,造成这一结果的原因可能是因为毛竹林集约经营过程中施肥增加了土壤中的氮素含量,一定程度上缓解了氨氧化微生物之间的养分竞争。
4 结 论
无论毛竹林经营强度如何AM 真菌菌丝均显著降低了土壤pH,且在集约经营毛竹林中AM 真菌菌丝显著降低了土壤碱解氮含量。不同菌根分室处理并未对硝化势产生显著影响,但经营强度影响显著。集约经营和AM 真菌菌丝分别对AOA 和AOB 基因丰度影响显著,虽然两者对氨氧化微生物群落结构没有显著影响,但分子生态网络分析显示集约经营缓解了氨氧化微生物之间的养分竞争。
猜你喜欢
天津农林科技(2022年2期)2022-04-19 10:48:14
昆明医科大学学报(2022年2期)2022-03-29 00:51:58
食品安全导刊(2021年20期)2021-08-30 06:40:50
乡村科技(2018年8期)2018-06-27 03:41:40
环境保护与循环经济(2017年7期)2018-01-22 02:58:12
湖南林业科技(2017年1期)2017-02-06 05:29:18
中国环境科学(2016年3期)2016-02-08 15:07:14
水生生物学报(2015年1期)2015-02-28 16:01:05
应用海洋学学报(2014年2期)2014-11-26 01:20:46
河南科技(2014年18期)2014-02-27 14:14:54
