
滇池流域磷矿山区优势植物叶片与土壤养分生态化学计量特征①
2021-05-16 13:34赵洛琪吴晓妮付登高徐子萱朱安琪
土壤 2021年2期
赵洛琪,吴晓妮,付登高*,徐子萱,朱安琪
(1 云南大学生态与环境学院暨云南省高原山地生态与退化环境修复重点实验室,昆明 650091;2 昆明学院农学与生命科学学院,昆明650214)
选择合适的植物物种进而构建植物群落一直是恢复生态学研究的重点领域。在受损生态环境中,植物不仅要面临土壤养分含量缺乏的限制,甚至会受到极端环境的胁迫,所以在植被恢复的过程中,植物的选择尤为重要[1]。针对不同类型 (如森林、草地、农田、湿地、湖泊、河流、海洋)、不同程度的退化生态系统,其恢复方法亦不同[2],而对于退化山区的植被恢复,首要任务是增加贫瘠土壤的养分含量,以缓解土壤养分含量对当地植被生长的限制作用。
生态化学计量学集生物学、化学计量学和其他相关学科的一些基本原理于一身[3-4],是研究生物系统能量平衡和多重化学元素平衡的一门学科[5]。目前国外有很多关于 C、N、P 化学计量学研究,研究对象由水生生态系统和湿地生态系统延伸到陆地生态系统,主要集中在种群动态变化和生态系统养分供求平衡及其应用[6-7]。土壤和植被是陆地生态系统的重要组成部分,两者之间具有密切的相互作用。土壤是植物生长的基础,为植物提供生长所需的各种养分,对植物群落的发生和演替,群落的物种组成、多样性以及生态系统的结构和功能具有重要影响[8-9]。因此,深入了解土壤及植物不同元素含量及其生态化学计量对探究土壤养分元素在循环过程中的相互耦合关系及其生态效应具有重要作用[10]。例如,植物体内的 C:N 和 C:P 通常能反映植物对N 和 P 养分的利用效率,一定程度上也能表征土壤中 N 和 P 的供应状况[11]。植物叶片的N:P 临界值被认为可以指示土壤对植物生长的养分供给状况,并且被广泛应用于诊断植物个体、群落和生态系统的 N、P 养分限制格局[5]。土壤作为植物 N、P 养分来源,直接影响植物对N、P 养分的吸收利用及植物化学计量关系[12],尤其在土壤养分比例失衡的立地条件下,探讨土壤和植物之间的养分含量及其计量特征将有助于生态修复进程及有效管理。
滇池流域南部是我国著名的磷矿开采区,占据了滇池流域总面积的13%。在该区域退化山地环境中,土壤中极高的P负荷极易产生淋溶和侵蚀,大量的P随着地表径流汇入滇池[13]。磷矿山区内一般土壤中含P量平均达到 0.42% 以上,远远高于本地土壤P正常值 0.14%,可溶磷含量达到 52 ~160 mg/kg,远远高于本地可溶磷正常值17.8 ~32.0 mg/kg,目前流域内人类的各种活动已显著地影响P素的生物地球化学循环。因此,该区域的生态修复对滇池富营养化的防控具有重要影响。前期调查发现该区域内存在多种不同生活型的优势植物,其中云南松(Pinus yunnanensis)能够应对水分与干旱的胁迫并适应云南雨旱分明的自然条件[14];马桑(Coriaria nepalensis)作为固N物种[15],能够减少土壤低N环境对自身生长发育的限制;蔗茅(Erianthus fulvus)在云南各地有着广泛的分布;而紫茎泽兰(Euapatorium adenophorum)作为入侵植物,对当地生物多样性和生态系统过程及功能的影响受到人们的持续关注[16]。因此本研究选取了滇池流域磷矿退化山区内4种不同生活型优势植物,通过对植物叶片以及在植物影响下的土壤营养元素含量及其计量比的分析,深入了解该区域土壤和植物的营养元素格局及影响植物生长发育的主要限制因子,并在此基础上为该区域生态修复过程中的植物选取及植物群落组配提供参考,以期达到固持水土、削减P素流失的效果。
1 材料与方法
1.1 研究区域概况
研究样地位于滇池流域南部,地理位置为24°36′ ~ 24°37′N,102°41′ ~ 102°42′E,海拔1 936 ~2 256 m,平均温度14.7 ℃,多年降雨量797 ~1 007 mm,大多集中在雨季的7、8月份。地貌为山地半山地,处于自东南向西北汇入滇池的柴河子流域内。该流域是滇池流域主要的磷矿开采区,也是滇池流域内3个面积超过300 km2的子流域之一。该研究区内有多处早年磷矿开采面,多分布于1/3 ~ 2/3山腰区域。土壤类型为山地黄红壤与棕红壤,土壤含水率为14.29%,pH 6.23,土壤全氮0.78 g/kg,全磷6.57 g/kg,全钾3.36 g/kg,有机质32.1 g/kg。流域内自然植被以亚热带半湿润常绿阔叶林为主,由于前期人类活动,几乎已没有成片分布。而在经自然恢复后植被覆盖度较好的区域内,广泛分布着成片的灌草丛及云南松林,两种植物群落的立地条件、土壤母质和水热条件等均相对一致。其中云南松林的优势乔木及灌木分别为云南松(P. yunnanensis)和碎米花杜鹃(R. spiciferum);灌草丛中优势灌木为马桑(C. nepalensis),优势草本植物为蔗茅(E. fulvus)和典型的外来入侵物种紫茎泽兰(E. adenophorum)。
1.2 样品的采集与参数测定
基于前期调查基础上,于2017年8月在该区域磷矿山区内选择典型的坡向、坡位相似的荒坡灌草丛及云南松林样地,各样地设4个大小为10 m ×10 m的样方,样方之间的距离不少于100 m。在灌草丛样地中,根据马桑、蔗茅及紫茎泽兰的生长情况,各物种随机选择植株高度相对一致、生长良好且成熟的植株各3株,4个样方中每个物种共选择12株个体。在云南松样地中,选择8株高度及胸径相似且长势良好的云南松,每个样方各2株。植株选择完毕后,在植物冠幅下采集土样,由于不同生活型植物根系分布的差异,故云南松采集0 ~30 cm土壤,其余3种植物采集0 ~ 20 cm土壤,在马桑、蔗茅及紫茎泽兰灌丛中随机采3个样点土壤样品进行均匀混合;由于云南松冠幅相比灌木和草本较大,故在云南松冠幅内随机采6个样点并均匀混合。采集样方内植物叶片,在4个方向各随机采集目标植物成熟叶片样品。叶片样品带回实验室后,用蒸馏水清洗干净后置于105 ℃ 烘箱内杀青20 min后调至80 ℃ 烘干至恒重,过0.1 mm筛后装瓶待测;土壤样品按测试要求风干研磨过2 mm和0.25 mm筛,以测定土壤养分指标。
土壤和叶片全氮的测定采取半微量凯氏定氮法,土壤速效氮的测定采取碱解扩散吸收法,土壤和叶片全磷的测定采取酸溶-钼锑抗比色法,土壤有效磷的测定采取双酸浸提-钼锑抗比色法,土壤和叶片有机质的测定采取重铬酸钾氧化外加热法,土壤pH的测定采取电极法[17]。
1.3 数据分析
利用单因素方差分析比较不同植物影响下的土壤和植物叶片的养分含量及其计量比之间的差异;利用相关性分析了解土壤与叶片养分含量之间的关系。其中土壤N有效性=碱解氮/全氮,土壤P有效性=有效磷/全磷。使用Excel 2007对数据进行整理归纳,所有统计分析均使用SPSS 23.0进行。
2 结果与分析
2.1 植物对土壤养分的影响
不同植物影响下的土壤养分含量如表1所示:云南松影响下土壤有机质、全氮、碱解氮、有效磷含量最高,分别为22.42 g/kg、1.85 g/kg、140.78 mg/kg和1 048.89 mg/kg,其中有机质含量显著高于其他植物;云南松下的土壤全磷含量最低,为2.51 g/kg,显著低于其他植物。pH与有效磷含量在不同植物之间差异并不显著。
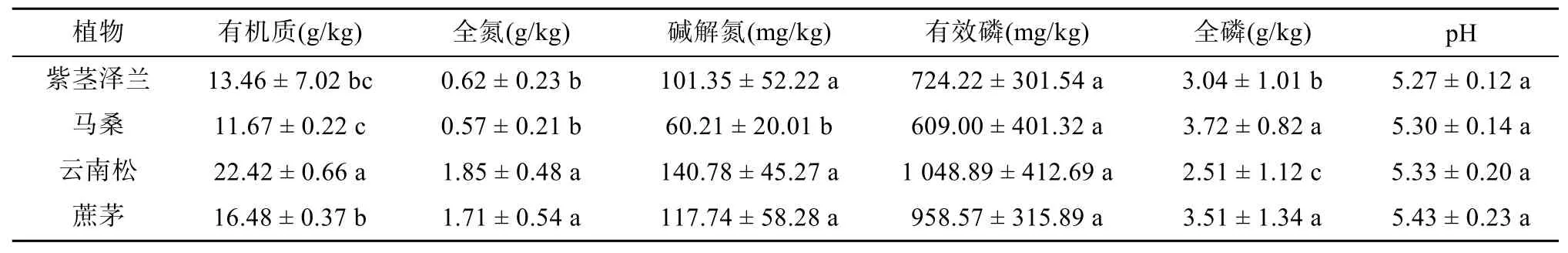
表1 不同植物影响下的土壤养分含量Table 1 Soil nutrient contents under different plants
2.2 植物对土壤生态化学计量比及氮磷有效性的影响
分析不同植物对土壤生态化学计量比及N、P有效性的影响可以发现(表2):云南松影响下土壤C︰P、N︰P分别为8.93和0.74,显著高于其他植物土壤,且P有效性最高;紫茎泽兰C︰N及N有效性最高,但差异并不显著。
2.3 不同植物叶片养分含量及生态化学计量比特征
不同植物叶片养分含量及其计量比特征具体见图1。不同植物叶片C含量表现为云南松叶片C含量显著高于其他植物。不同植物叶片N含量表现为紫茎泽兰>马桑>蔗茅>云南松,其中紫荆泽兰和马桑叶片N含量显著高于云南松和蔗茅。不同植物叶片P含量表现为紫茎泽兰>马桑>蔗茅>云南松,且相互之间均差异显著。不同植物叶片C︰N和C︰P均表现为云南松>蔗茅>马桑>紫茎泽兰,且云南松显著高于其他植物;不同植物叶片N︰P表现为马桑和紫茎泽兰显著高于蔗茅和云南松。
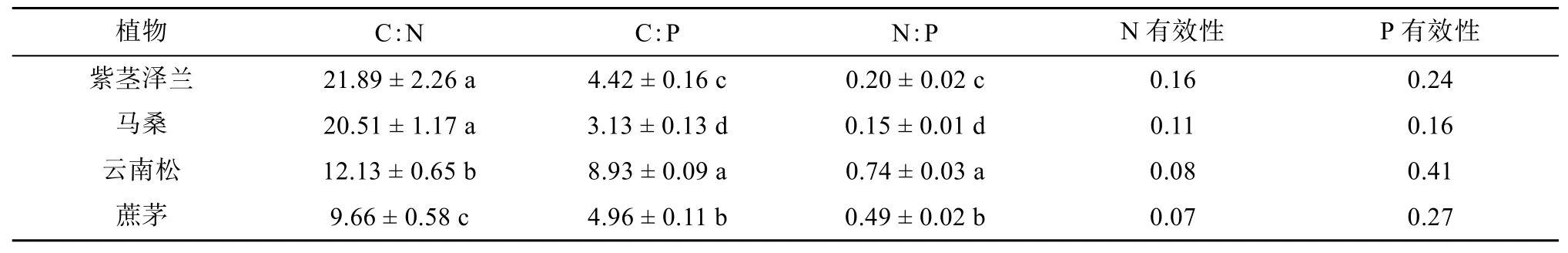
表2 不同植物影响下的土壤C、N、P计量比及土壤N、P有效性Table 2 Eco-stoichiometric ratios among soil C, N and P and soil N and P availability under different plants
2.4 不同植物影响下土壤与叶片养分含量相关关系
不同植物影响下土壤与叶片养分含量及其化学计量比的相关关系见图2。其中土壤C含量与叶片C含量及叶片C︰P均表现出显著的正相关关系。土壤N含量与叶片C含量、叶片C︰N呈显著正相关,而土壤N含量与叶片N︰P表现出显著的负相关关系。土壤P含量与叶片P含量呈显著正相关,但与叶片C含量及叶片C︰P均表现出显著负相关。
3 讨论
3.1 磷矿山区土壤养分含量及其生态化学计量特征
从整体来看,磷矿山区不同植物影响下土壤C、N、P含量为16、1.19、3.19 g/kg,相较于该区域磷矿开采废弃地土壤C、N、P含量1.82、0.66、20 g/kg[18],C、N含量明显上升,P含量明显下降。而磷矿山区土壤在不同植物影响下,各项养分指标表现出不同的变化。云南松影响下土壤C、N、P含量分别为22.42、1.85、2.51 g/kg,云南松能够提升土壤C、N养分含量,并且通过吸收削减土壤P含量;荒坡灌草丛土壤C、N、P含量分别为13.87、0.97、3.42 g/kg。相较于滇中非磷矿区土壤平均C、N、P含量20.25、0.78、0.2 g/kg[19],磷矿区荒坡灌草丛土壤C含量偏低,N含量相近;而云南松群落土壤养分状况较好。
研究表明土壤养分组成是植物外部环境中重要的影响因子,植物的光合作用、矿质代谢等过程与土壤养分供应状况关系密切[20]。土壤养分计量比可以从以下4个方面进行分析(表2):①土壤C︰N:土壤C︰N是土壤质量的敏感指标,影响土壤中有机C和N的循环[21]。一般来讲,土壤C︰N与有机质分解速率呈反比关系[22]。本研究表明植物影响下的土壤C︰N的范围为9.66 ~ 21.89,大体高于中国土壤C︰N平均水平10 ~ 12[23],说明该区域土壤有机质的分解速率相对较慢,进而限制养分循环的速度,尤其对于云南松和蔗茅,较低的C︰N表明其凋落物分解是影响养分循环的关键过程之一;②土壤C︰P:较低的C︰P 可以作为土壤磷素高有效性的一个指标[9],在本研究中植物影响下的土壤C︰P为3.13 ~ 8.93,P的有效性平均为0.28,虽然研究表明该地区最容易被植物利用的树脂态磷在土壤全磷中只占0.15%[24],但由于研究区域内土壤P含量的高背景值,并结合不同植物叶片P含量来看,土壤P含量能够满足当地植物生长发育的需要,所以相较于土壤P,植物生长受到C的限制;③土壤N︰P:土壤中的N、P是植物生长所必需的矿质营养元素和生态系统中最常见的限制性元素,二者之间具有重要的相互作用,因此将N︰P作为衡量系统N、P养分限制的一个指标。本研究中土壤N︰P介于0.15 ~ 0.74,远低于全球森林土壤N︰P(6.60)[25],这不仅与研究区域内土壤较高磷素背景值有关,并且表明研究区域内植物生长受到土壤N含量限制;④土壤N、P有效性:土壤N、P有效性可以表明土壤N、P养分的有效利用程度,其中由于较高的P素背景值,其有效磷不是影响植物生长的限制因子;但对于N有效性而言,云南松和蔗茅明显低于马桑和紫荆泽兰,说明云南松和蔗茅不仅受到N素含量的影响,其N的有效性也是影响该两种植物生长的限制因子。综合以上分析,在磷矿山区,土壤C、N是影响植物生长的主要限制因子,尤其对于云南松和蔗茅而言,其养分限制程度更高。
3.2 磷矿山区不同生活型植物叶片养分含量及其生态化学计量特征
云南松叶片C含量为492.86 g/kg,大于全球叶片平均C含量(461.6 g/kg)[23],荒坡灌草丛植物叶片C含量低于全球平均水平,原因可能为根系较浅,无法获得足够的水分,导致叶片的气孔导度降低从而减少叶片CO2的吸收[25];叶面积减小,ATP 的合成减弱,羧化酶活性降低,导致光合作用受阻,阻碍了C的固定[26]。叶片N含量介于11.26 ~ 24.68 g/kg,接近于中国陆地植被叶平均N含量(18.6 g/kg)[27],N元素是植物叶片光合作用酶的重要组成成分,叶片N含量可以表征植物叶片光合作用速率的高低,并且植物体内的 N 以蛋白酶的形式参与 C 的固定[28];紫茎泽兰叶片N含量最高,旺盛的光合作用导致紫茎泽兰叶片中C的积累与自身快速生长繁殖。植物叶片P含量介于2.78 ~ 4.07 g/kg,远高于中国陆地植被叶平均P含量 1.21 g/kg[27],这与当地土壤高P含量有关。P以核酸的形式参与蛋白酶的装配并且以ATP的形式进行能量的传递[28],高P含量有利于植物自身的能量传递。
通常情况下,植物叶C︰N和C︰P能反映植物N和P的养分利用效率,植物在养分供应缺乏的情况下往往具有较高的养分利用效率,相反,营养元素供应充足时,养分利用效率较低[29]。研究区域内植物叶片C︰N介于17.46 ~ 43.95,而全球植物叶片平均C︰N为23.8[27],表明在C、N养分相对匮乏的情况下,云南松和蔗茅叶片能够固定更多的C;相反紫茎泽兰和马桑叶片能够固定较多的N,在土壤N含量限制的条件下也能够较为良好地生长发育。Rong等[30]的研究结果表明叶片C︰N和C︰P较高的植物,在固碳和养分利用策略等方面都具有优势。植物叶片C︰P均远低于全球平均水平(300.9)[27],可能与研究区域内土壤P含量的高背景值有关。植物叶片的N︰P可衡量植物生长对于N和P的倚重程度的大小,Koerselman和Meulema[31]研究发现,当叶片的N︰P<14时,植物生长主要受N限制;当14
3.3 不同植物叶片、土壤养分相关性特征
植物与土壤作为生物地球化学循环的不同环节,两者之间存在必然的联系,植物体内养分含量体现了植物对环境适应特征,土壤养分条件反映了植物的营养状况[32]。李从娟等[8]研究表明在自然条件下植物的生境存在异质性,土壤养分也具有明显的差异性,而这种差异则会影响植物叶片的化学计量特征。在本研究中,植物叶片C含量与土壤N含量存在显著正相关关系,这与陆远鸿等人[33]的研究结果相同。土壤N含量能够有效促进植物C含量的积累,表明植物生长发育受到高度的N限制。植物叶片C︰N与土壤N含量表现出极显著的正相关关系,说明植物在生长发育过程中不仅受到N限制,并且植物对N元素的利用效率较高。植物叶片与土壤P含量负相关,说明土壤P含量过剩[34]。磷矿山区植物叶片及土壤C、N、P含量化学计量比的变化以土壤C、P元素变化为主导,在土壤N元素缺乏的条件下,固N物种的存在十分必要。
3.4 不同生活型植物属性特征及其磷矿山区的植被恢复
云南松为当地优势乔木,土壤有机质、氮及有效磷含量均为最高,表明云南松对土壤有很高的改良效益,云南松林内枯落物厚度大,能够有效增加地表粗糙程度,枯落物腐烂增加土壤有机质含量、土壤孔隙度等,改变土壤入渗能力、含蓄水分,进而增强植物群落改良土壤的能力[35]。虽然云南松叶片P含量较低,但由于云南松生物量远高于其他目标植物,能够固定大量土壤P,达到削减土壤P素流失的目的。马桑土壤的有机质含量虽然最低,但马桑作为固N物种,由于固N菌的存在,能在根际形成较高的N含量,能够适应并逐步改良磷矿山区土壤N缺乏的条件[36-37]。蔗茅作为本地优势物种,其贫瘠土壤的适应能力较强,并能够改善土壤C和N含量,但其土壤N的有效性不高,因此可以与固N植物马桑构建荒坡灌草丛群落以提高土壤养分,并改善土壤水土保持功能。作为入侵植物的紫茎泽兰,其能够在土壤C、N元素缺乏的条件下快速生长繁殖,这对于进一步探究植物对C、N养分贫瘠土壤适应的机制具有重要意义,但其作为入侵植物,强烈的化感作用容易形成单优种并对当地的生态环境造成一系列的负面影响,故不推荐用于植被恢复。
研究结果表明:磷矿山地土壤有机质及N是影响植物生长的主要限制因子,在这种C、N养分相对匮乏的情况下,云南松和蔗茅叶片能够固定更多的C;相反马桑叶片能够固定较多的N。因此,结合不同生活型植物的属性特征及该区域内群落的演替特征,磷矿山区废弃地的生态恢复可以通过构建蔗茅和马桑为主的灌草丛植物群落,一方面能够适应较贫瘠的立地环境,另外也能在一定程度上改善立地环境。当立地达到一定条件,引入乔木云南松,形成乔灌草立地群落,并逐步向云南松林进行演替;由于云南松生长受土壤N素的限制,在演替后期,引入固N阔叶乔木,如旱冬瓜,形成针阔混交林[38]。
因此,在磷矿退化山区的植被恢复过程中,从土壤养分含量的角度出发,应在开始阶段人工引入马桑与蔗茅形成荒坡灌草丛群落,以逐步缓解土壤N含量限制的状况;进而引入云南松以增加土壤C含量,随后适当种植以旱冬瓜为代表的固N阔叶树种形成针阔混交林,以达到全面改善土壤养分,保持水土并控制土壤P素流失的目的。
猜你喜欢
中国土壤与肥料(2022年4期)2022-06-14
农业工程学报(2022年1期)2022-03-25
有色金属(矿山部分)(2021年4期)2021-08-30
英语文摘(2021年3期)2021-07-22
落叶果树(2021年6期)2021-02-12
文苑(2020年12期)2020-04-13
小星星·阅读100分(低年级)(2019年10期)2019-11-26
安徽文学(2019年6期)2019-06-28
海峡姐妹(2019年1期)2019-03-23
