
舟山近岸海域春秋季鱼类幼鱼资源现状与保护分析
2021-04-29 06:44俞存根于南京张佩怡蒋巧丽牛威震
渔业研究 2021年2期
刘 坤,俞存根,于南京,张佩怡,蒋巧丽,牛威震
(浙江海洋大学水产学院,浙江 舟山 316022)
舟山渔场在历史上曾久负盛名,因其受大陆入海径流、台湾暖流以及黄海冷水团的交汇影响,海域生物资源种类繁多,是多种海洋经济鱼类繁衍的栖息地,主产大黄鱼(Larimichthyscrocea)、小黄鱼(Larimichthyspolyactis)、带鱼(Trichiurusjaponicus)等,是我国重要的海洋渔业资源分布区。然而在栖息地破坏、水域环境恶化及过度捕捞等因素的影响下,舟山近岸渔业资源出现了严重的衰退,主要表现为生物多样性降低、鱼类低龄化与小型化、渔获物营养层级下降等现象[1]。尽管当前国家已采取严格的措施来恢复渔业资源,如延长伏季休渔时间、实行渔船“双控”进一步压减捕捞努力量、严格限制网目尺寸等,但渔业资源总的衰退趋势并没有得到有效遏制[2],性成熟提前依旧未改观[3],加之捕捞对渔业个体大小选择性可能导致其生物学特征发生适应性演变,在缺少反向压力的情况下,生物学特征的恢复所需时间要远超过其生物量[4-5],因此有必要研究鱼类的幼鱼损害情况,弥补因损害幼鱼资源造成的渔业资源衰退。
幼鱼是指鱼的外形特点和栖息习性等已和成鱼一致,而性腺尚未成熟的阶段,是鱼类体长生长的快速期[6]。当前已有较多学者对幼鱼资源保护问题进行了研究,如张网作业对带鱼等渔获物幼鱼的损害状况分析[7-12]、定置作业对渔获群体组成特点变化的影响及幼鱼损害分析[13-14],以上研究重点突出了保护幼鱼对合理利用近海渔业资源的重要性。为进一步明晰舟山近岸海域渔业资源利用现状,促进幼鱼资源的合理养护,本文依据2015年11月(秋季)、2016年5月(春季)在舟山近岸海域进行的渔业资源拖网调查数据,分析了该海域的渔业资源量、渔获种类组成及其生物学特征等,重点分析该海域幼鱼资源情况,以期更好地掌握鱼类资源利用现状,强化幼鱼保护对舟山近岸海域渔业资源可持续利用的重要性,以期为舟山近岸海域的生态保护和渔业管理等提供基础资料。
1 材料与方法
1.1 数据来源
1.1.1 调查及采样方法
本次调查方法按照《海洋渔业资源调查规范》(SC/T 9403—2012)[15]和《建设项目对海洋生物资源影响评价技术规程》(SC/T 9110—2007)[16]进行,调查海域共设置80个站位(图1),调查范围为29°20′~31°00′N、121°40′~123°00′E,调查时间为2015年11月(秋季)、2016年5月(春季),所用调查船为群众单拖网渔船,船号为“浙普渔32207”,渔船主机功率为330 kW;调查网具为底拖网,网具规格为800目×80 mm,囊网网目尺寸为20 mm。每站拖曳时间为1 h,拖速为3.5 kn。每站拖网所获的渔获物全部取样装入样品袋,并进行编号、记录后,冰鲜保存,带回实验室分析、鉴定,统计渔获种类的尾数和重量,称重使用精确度为0.1 g的电子天平。
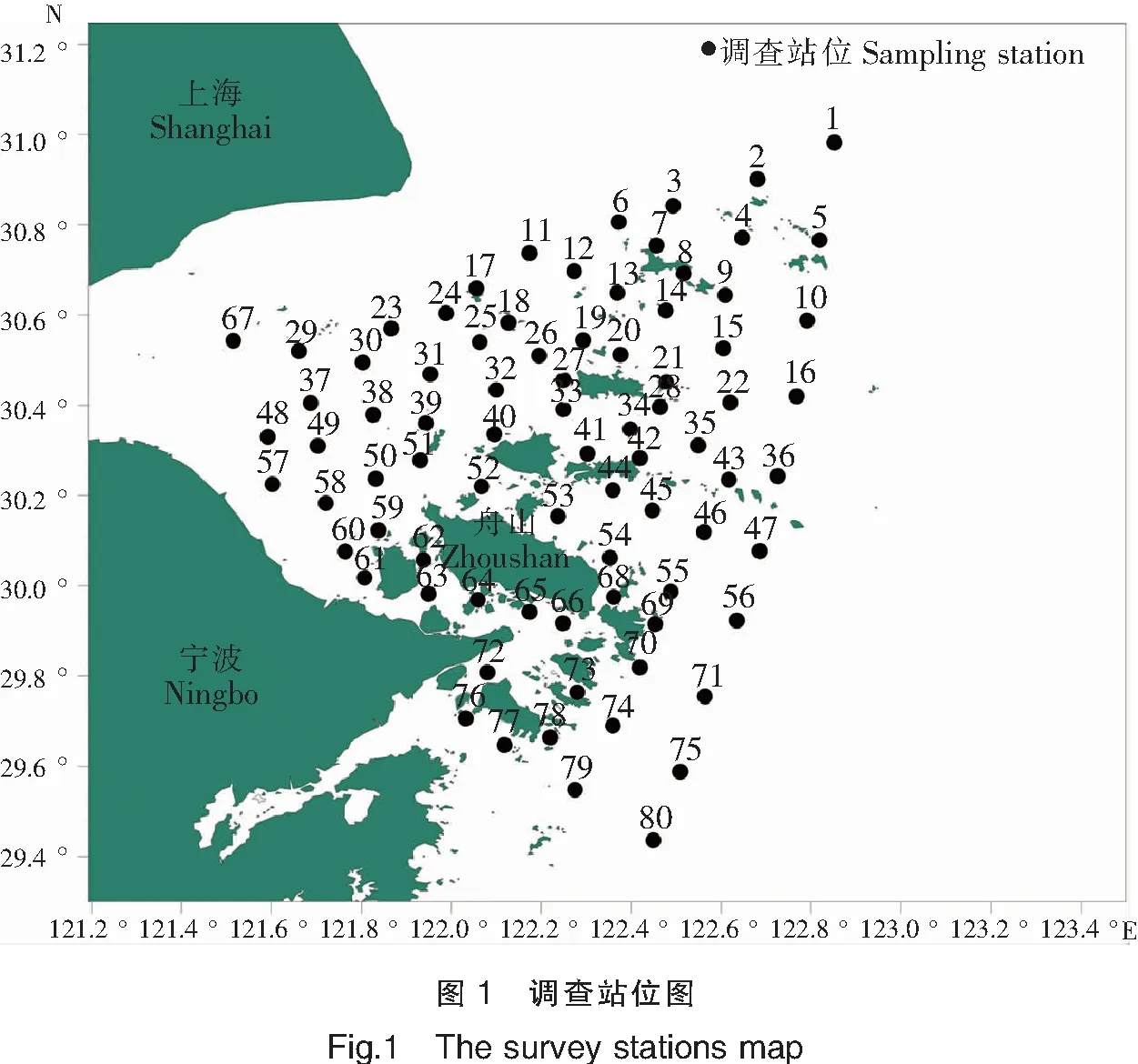
1.1.2 调查海区划分
舟山渔场及其临近海域是我国最大的近海渔场,依据渔民生产季节、作业工具及捕捞对象,可将舟山渔场划分为大戢渔场、洋山渔场、嵊山渔场、岱衢渔场、黄大洋渔场、中街山渔场及金塘渔场等[17],本研究参照历史上的生产渔场分布情况,结合不同海域的水文环境、渔业资源数量和种类、所处海域、临近岛屿等地理标记,将本次调查海域划分为10个区域,不同区域所包含的站位如表1所示。
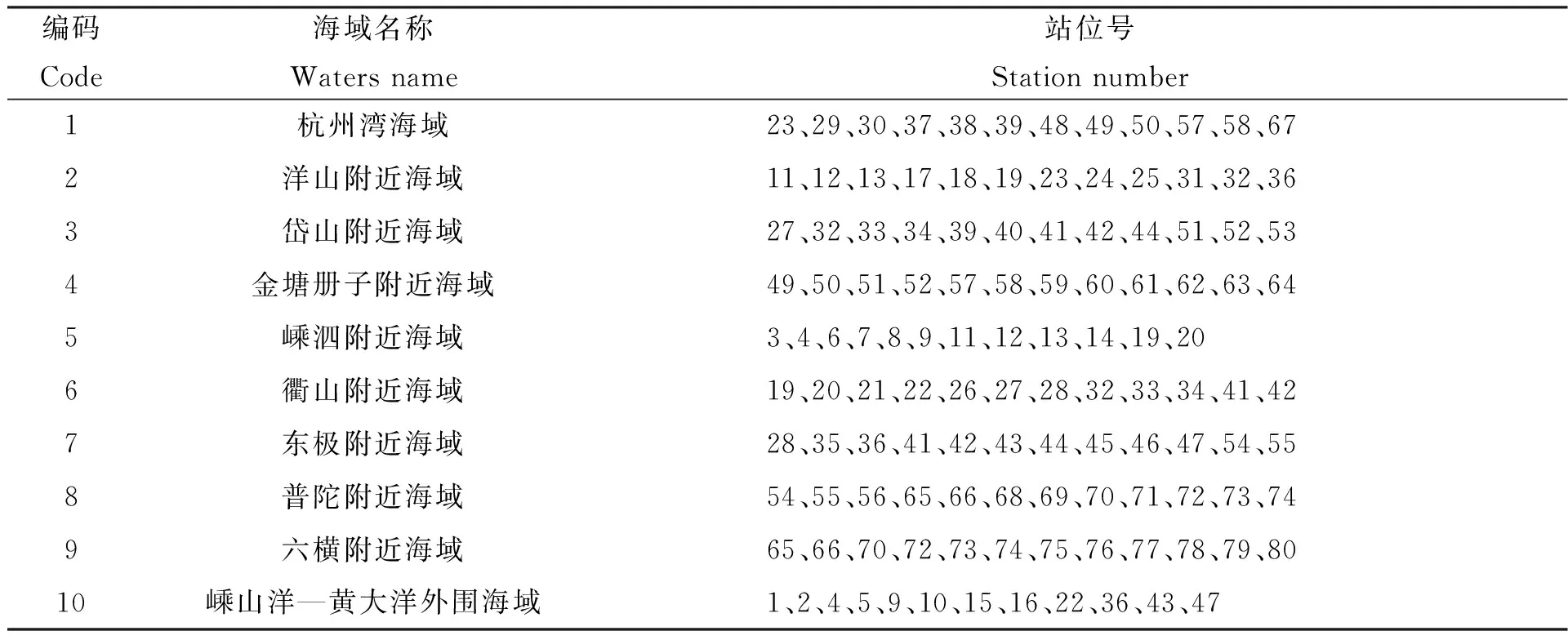
表1 调查区域划分
1.2 数据处理
1.2.1 主要鱼类的选定及其季节更替率
鱼类群落优势种测度采用相对重要性指数(IRI)[18],计算公式如下:
IRI=(N+W)×F
(1)
式(1)中:N为某鱼类个体数占鱼类总尾数的比例;W为某鱼类重量占鱼类总重量的比例;F为某鱼类出现的站位数占总站位数的百分比。鱼类优势种、常见种的划分参照俞存根等[19]的划分标准,将IRI>1 000的物种定为优势种,1 000>IRI>100为常见种,并以严利平等[20]的相关研究为参考,将IRI>100的种类(包括优势种和常见种)视为群落的主要种类。
本文对幼鱼的划分主要参照国家或行业标准规定的最小可捕标准,其中国家或行业标准有规定的种类参照标准进行划分,如2017年浙江省发布的《关于海洋渔业资源重点保护品种最小可捕过渡性规格的通告》、农业农村部2018年起实施的带鱼等15种重要经济鱼类最小可捕标准及幼鱼比例的管理规定等,其他没有规定的鱼类主要参考历史研究报道的最小性成熟体长或体重。
主要鱼类的季节性种类更替率(R)[21]计算公式如下:
R=(a+b-2c)/(a+b-c)×100%
(2)
式(2)中,a代表春季主要鱼类物种数;b代表秋季主要鱼类物种数;c为春秋季共有的主要鱼类物种数。
1.2.2 资源量估计
首先采用扫海面积法[22]对资源密度进行计算,具体公式如下:
Di=Ci/(aiq)
(3)
式(3)中:Di是第i站位的渔业生物的重量密度(kg·km-2);Ci是第i站位中每小时所捕渔业生物的重量(kg·h-1);ai是第i站位所用网具每小时扫海面积(km2·h-1),即网口横向扩展宽度与拖曳距离之积,拖曳距离等于拖速(km·h-1)×拖时(h);q为网具的捕获率,取0.5。
渔业资源量计算公式如下:
(4)
式(4)中:N为整个调查区域的现存资源量(t);Di为第i站位的资源密度(t/km2);A为调查区域的面积(km2);n为站位数。
1.2.3 平均拥挤度
平均拥挤度反映了鱼类的空间分布情况,该指标可以揭示不同鱼类的聚集强度[23]。计算公式如下[24]:
(5)
式(5)中,x为总调查站位的某鱼类平均渔获尾数;S2为方差。 某鱼种的平均拥挤度x*越大说明该鱼种聚集度强,在分布范围上集中度高[25]。
1.2.4 种间相遇机率
种间相遇机率(原称物种多样性指数)作为一种数学统计方法,常用于群落数量特征的分析,该法常见于于森林及草地群落不同种类个体分布机率的分析[26]。其公式为:
(6)
式(6)中,S为物种数;N为所有种的个体总数;Ni为第i种的个体数。
本文使用Surfer 11.0软件对调查海域站位经纬度数据进行可视化标注处理,相对重要性指数(IRI)、资源密度(Di)、资源量(N)、平均拥挤度(x*)及种间相遇机率(PIE)的计算在Microsoft Excel 2016中进行。
2 结果与分析
2.1 物种组成
根据本次调查所捕获的渔获物,春季分析鉴定出鱼类58种,隶属于11目34科49属;秋季分析鉴定出鱼类53种,隶属于11目25科41属。根据相对重要性指数(IRI)的计算结果(表2),调查海域春季优势种3种,分别为褐菖鲉(Sebastiscusmarmoratus)、棘头梅童鱼(Collichthyslucidus)和六丝钝尾鱼叚虎鱼(Amblychaeturichthyshexanema);秋季优势种1种,为龙头鱼(Harpodonnehereus)。
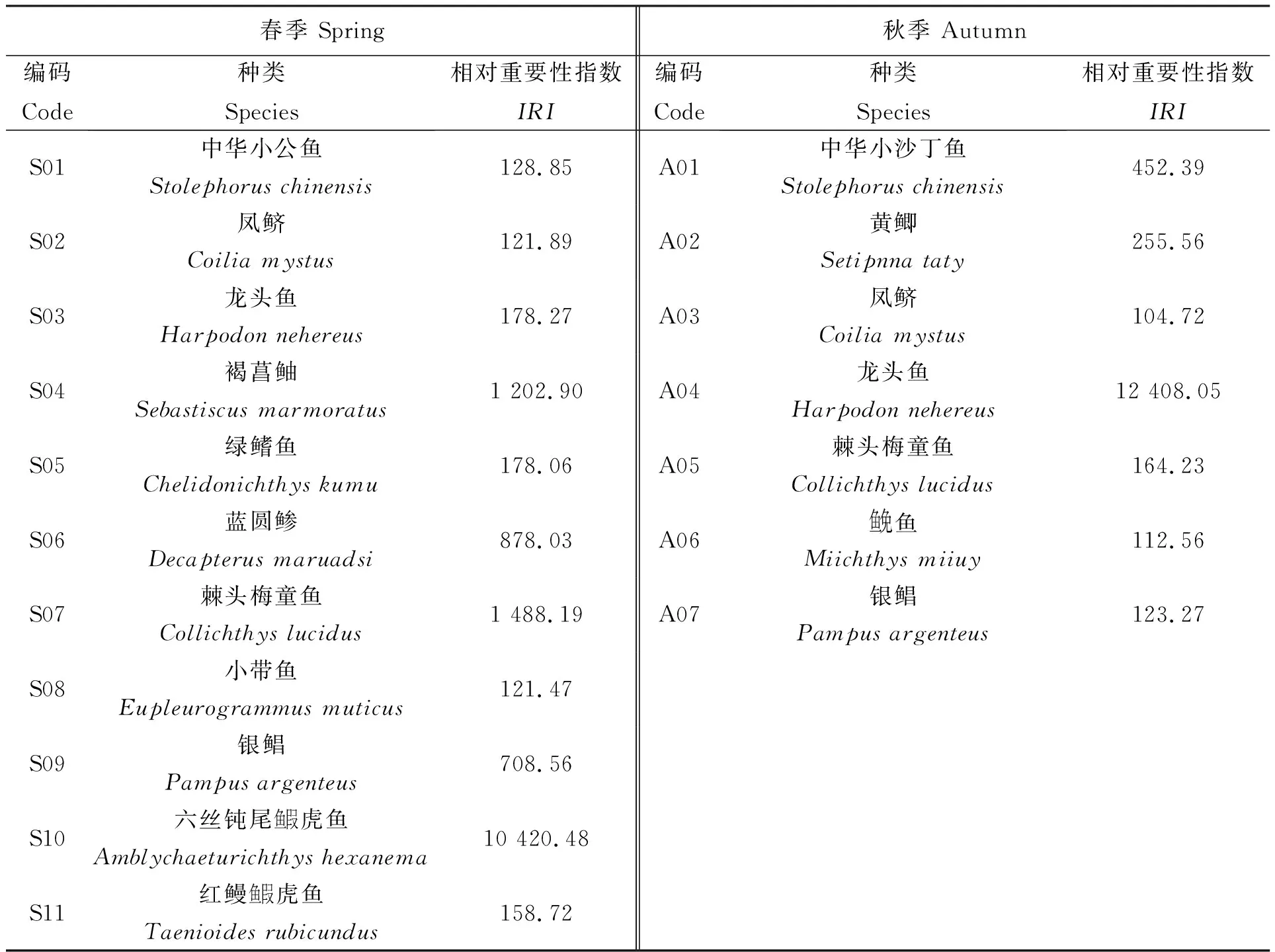
表2 春、秋季主要鱼类相对重要性指数
春季主要种有8种,分别为中华小公鱼(Stolephoruschinensis)、凤鲚(Coiliamystus)、龙头鱼、绿鳍鱼(Chelidonichthyskumu)、蓝圆鲹(Decapterusmaruadsi)、小带鱼(Eupleurogrammusmuticus)、银鲳(Pampusargenteus)和红鳗鱼叚虎鱼(Taenioidesrubicundus);秋季主要种有5种,分别为:中华小沙丁鱼(Stolephoruschinensis)、黄鲫(Setipnnataty)、凤鲚、棘头梅童鱼、鱼免鱼(Miichthysmiiuy)和银鲳。调查海域春秋季主要鱼类更替率为71.43%。
主要鱼类的幼鱼所占比例总体上春季高于秋季(表3),春季单鱼种幼鱼所占比例为100%的鱼类有8种,占春季主要鱼类总种类数的72.73%,其次银鲳的幼鱼占比也较高(86.67%);秋季幼鱼所占比例较高的仅鱼免鱼1种(88.24%),其次为龙头鱼(56.67%),其余种类如凤鲚、黄鲫等的幼鱼所占比例均较低。而主要鱼类的体重及体长平均值秋季均高于春季,生长优势较明显。
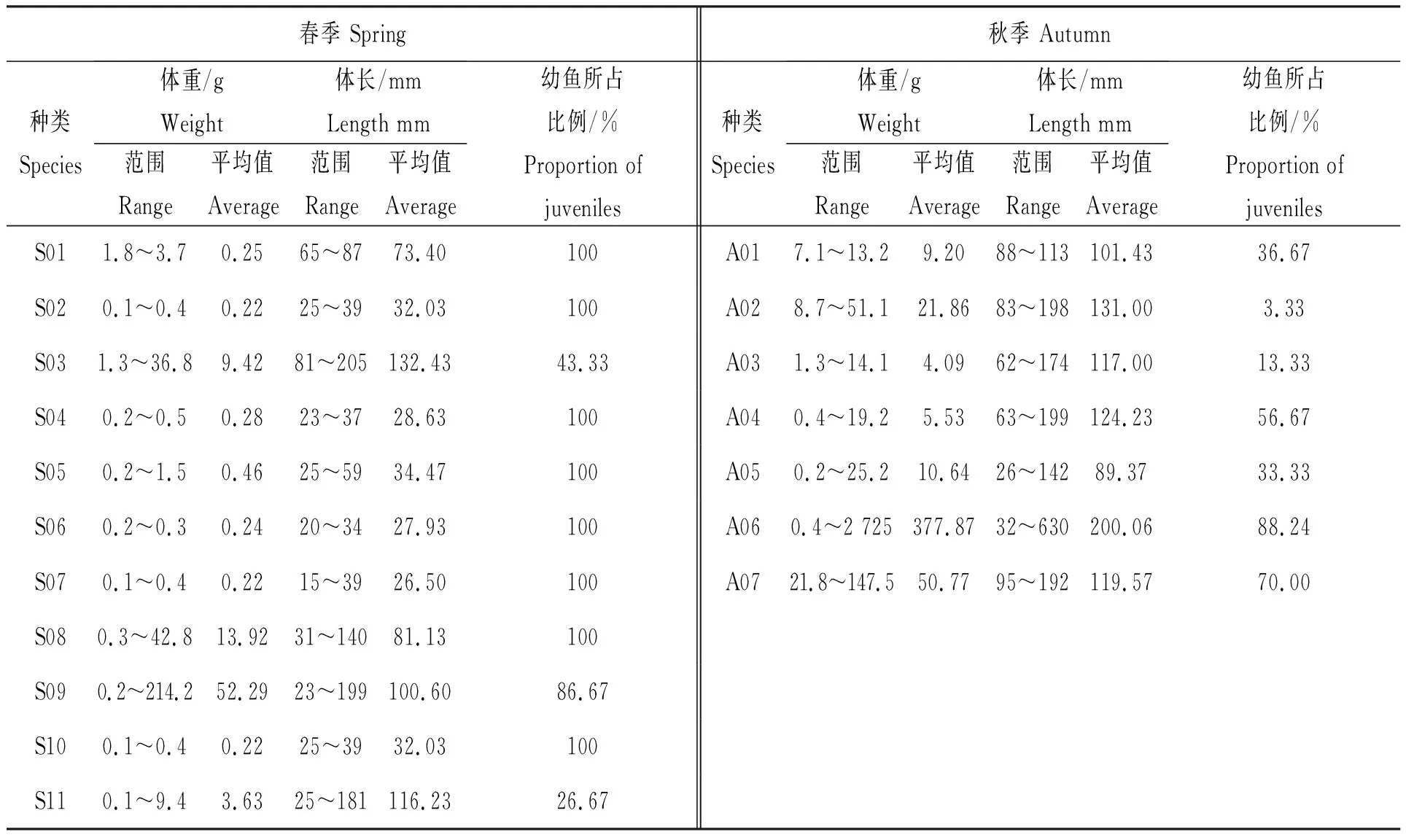
表3 春、秋季主要鱼类体重、体长分布及幼鱼所占比例
2.2 渔业资源重量密度与鱼类重量密度及其资源量估算
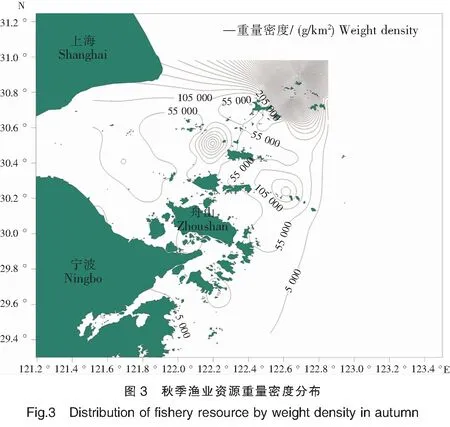
春季,以东极附近海域调查站位(表1)及嵊山洋—黄大洋外围海域调查站位的渔业资源重量密度最高,调查海域东部及东北部渔业资源的重量密度总体上高于122°E以西的杭州湾等海域(图2);秋季,整个调查海域渔业资源的重量密度与春季相比明显较高,重量密度较高的站位集中于嵊泗附近海域及嵊山洋—黄大洋外围海域的东北部,外侧海域总体上高于内侧海域(图3)。
利用扫海面积法估算不同区域春、秋季鱼类资源重量密度以及资源量(表4),结果显示,鱼类生物的重量密度秋季比春季大,嵊山洋—黄大洋外围海域春、秋季鱼类重量密度均为最大,其次为嵊泗附近海域、东极附近海域等依次递减,最小的为六横附近海域;此外,各海区鱼类资源量的评估结果与重量密度计算结果一致,海区间鱼类平均资源量差异较大,总体上距离岛屿海岸较远的海域鱼类资源量较多,鱼类资源量较大的如嵊山洋—黄大洋外围海域(485.26 t)、嵊泗附近海域(154.19 t)等,而靠近海岸受人类活动及海岸工程建设项目影响较大的海域鱼类资源量相对较低,如金塘册子附近海域(21.06 t)、六横附近海域(35.51 t)等;从季节差异上来看,鱼类重量密度及资源量秋季均大于春季,反映出鱼类资源状况的海区差异性和季节差异性。
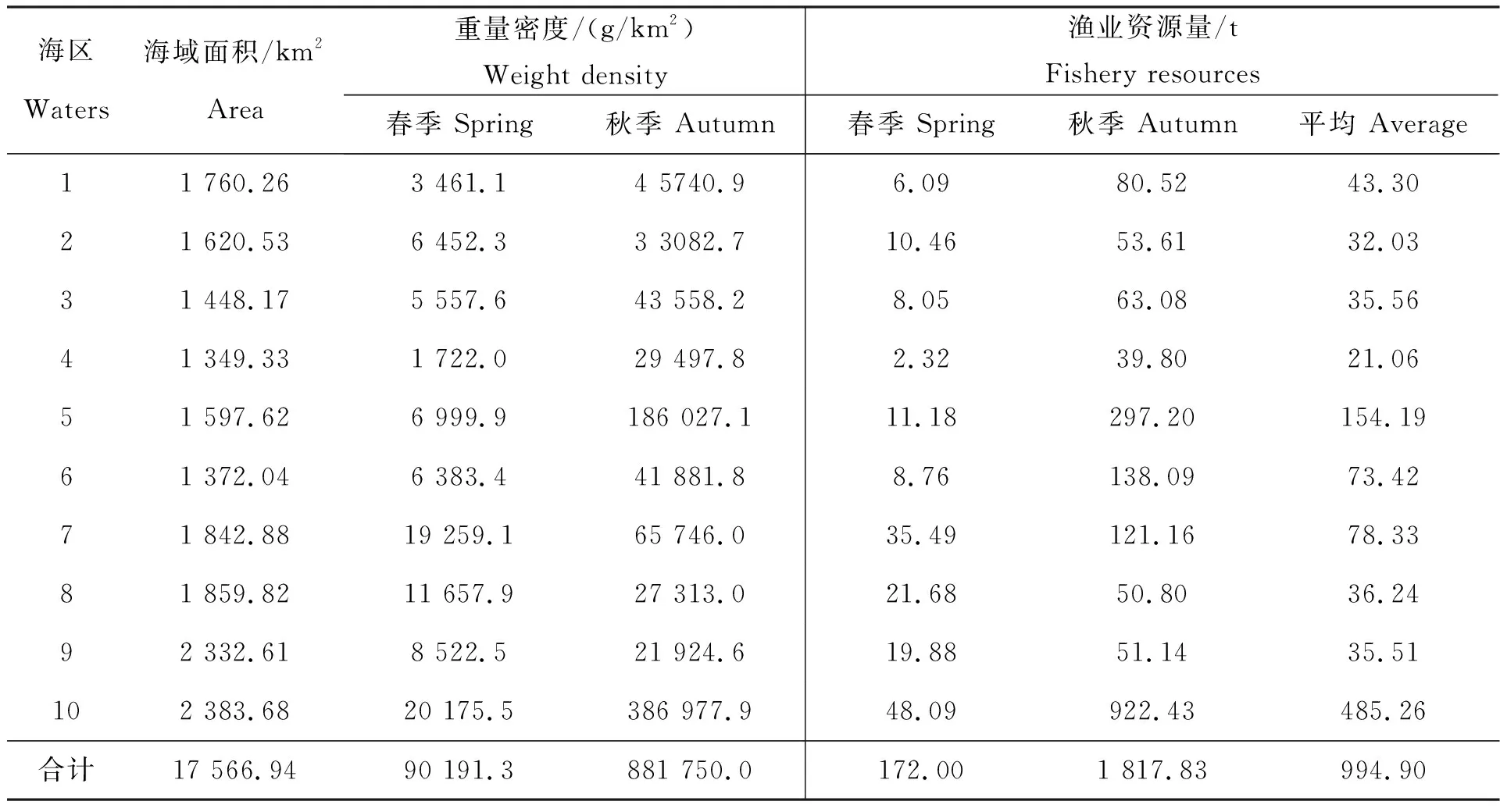
表4 春、秋季不同海区鱼类重量密度及资源量

2.3 平均拥挤度
平均拥挤度指数(x*)(表5)表明了春、秋两季主要鱼类聚集强度的差异性。春季,六丝钝尾鱼叚虎鱼的平均拥挤度(1 882.06)最高,聚集程度最强,其次为蓝圆鲹、棘头梅童鱼等,平均拥挤度最低的为红鳗鱼叚虎鱼(4.92);秋季,平均拥挤度以龙头鱼(4 230.85)为最高,物种的空间聚集程度远超其他鱼类,其次为中华小沙丁鱼、黄鲫等,平均拥挤度最低是鱼免鱼(44.15)。
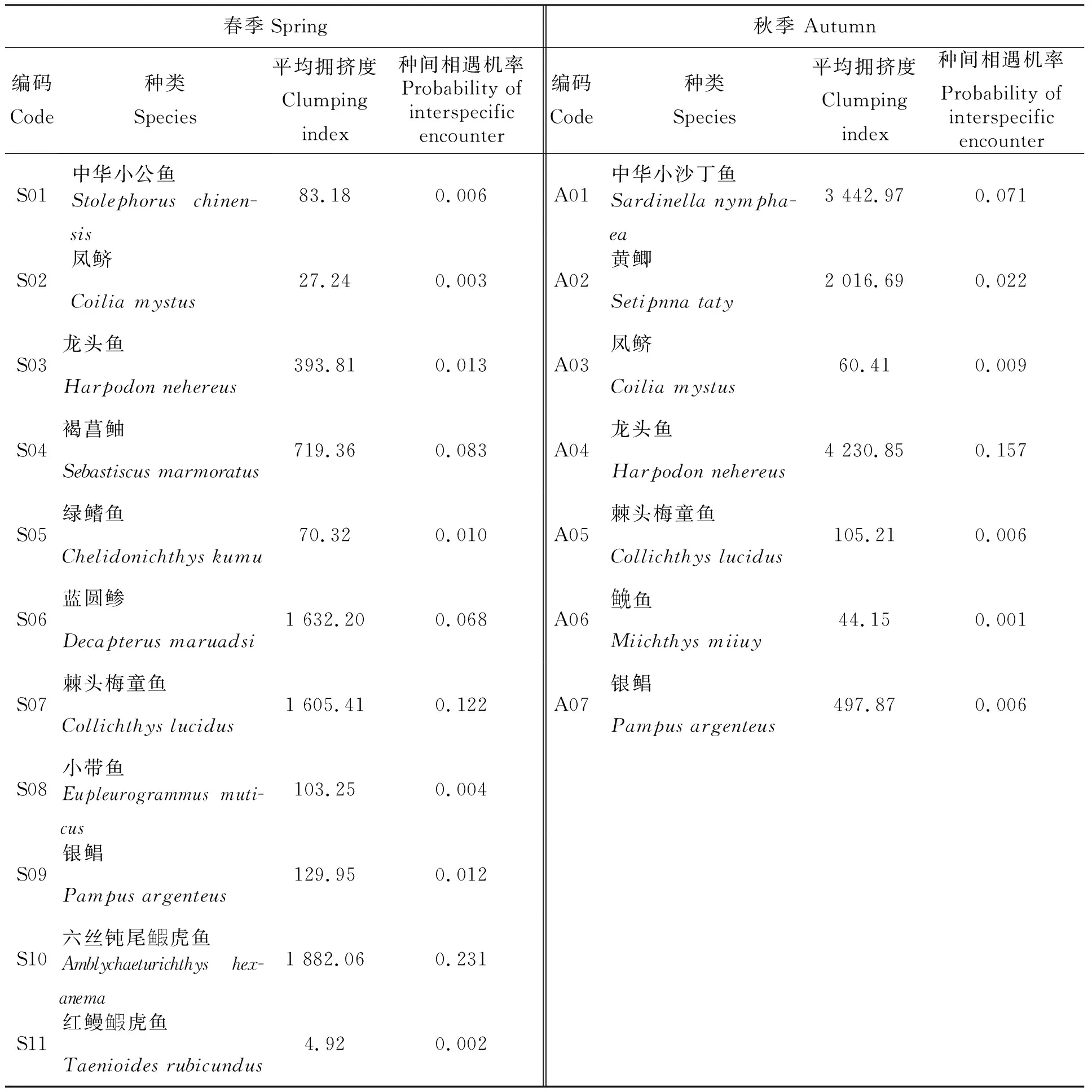
表5 春、秋季主要鱼类平均拥挤度及种间相遇机率
春季主要鱼类种间相遇机率总和为0.554,秋季主要鱼类种间相遇机率总和为0.272,春季群落内物种关联度较秋季更紧密。春季种间相遇机率最高的物种为六丝钝尾鱼叚虎鱼(0.231),该物种尾数占春季鱼类总尾数的63.76%,其出现的站位数占调查站位总数的98.75%;种间相遇机率最低的物种为红鳗鱼叚虎鱼(0.002),该物种尾数占春季鱼类总尾数的0.18%,其出现的站位数占调查站位总数的48.75%。秋季种间相遇机率最高的物种为龙头鱼(0.157),该物种尾数占秋季鱼类总尾数的80.52%,其出现的站位数占调查站位总数的100%;种间相遇机率最低的物种为鱼免鱼(0.001),该物种尾数占春季鱼类总尾数的0.09%,其出现的站位数占调查站位总数的13.75%。种间相遇机率与平均拥挤度计测结果基本一致。
3 讨论
3.1 渔业资源结构分析
本次调查中鱼类优势种与已有研究差别较大,如李建生等[27]20世纪90年代的研究表明,长江口临近海域优势种为带鱼、小黄鱼、白姑鱼(Argyrosomusargentatus)、黄鲫等,到21世纪初演变为龙头鱼、赤鼻棱鳀(Thrissakammakensis)、银鲳等中上层小型鱼类[28]。本次调查与俞存根等[17]2011年的舟山渔场调查也存在较大差异,特别是鱼类优势种从黄鲫、龙头鱼、海鳗(Muraenesoxcinereus)、大黄鱼、小黄鱼、棘头梅童鱼等经济价值较高的种类,演变为目前的褐菖鲉、棘头梅童鱼、六丝钝尾鱼叚虎鱼和龙头鱼4种。究其原因,一方面可能是因为过度捕捞兼捕了大量的经济鱼类幼鱼,引起渔业资源优势种衰退和群落结构演替;另一方面也与调查海域范围、站位设置以及所用网具不同有关,本次调查海域主要在舟山群岛的近岸海域,捕捞强度较大,同时海洋工程项目的实施使近岸海域水动力环境等发生改变,造成产卵场生境退化,成为“不育区”或者是鱼卵、仔稚鱼无法存活,导致渔业生态效益下降[29],生态系统的脆弱性更加明显,生物群落结构演替更加突出,探究可行的幼鱼资源生态损害补偿有助于合理完善海岸建设项目生态补偿机制。
相对重要性指数(IRI)反映了鱼类群落的优势度,春季优势种3种,群落的优势度较秋季相比更为集中,而秋季优势种仅龙头鱼1种,且IRI指数值(12 408.05)较大,远超其他鱼类,从渔业资源增殖角度出发,龙头鱼种群在繁衍补充上的优势度更加凸显,若考虑到整个调查海域群落结构的稳定性,这一优势种突出意味着其他鱼类生物多样性的降低[19],龙头鱼是我国近海渔业常见的捕捞对象[30],作为中下层肉食性鱼类,其主要的摄食类群为鱼类,大量吞食主要经济鱼类的稚、幼鱼,饵料生物中龙头鱼、小黄鱼、细条天竺鲷(Apogonichthyslineatus)、皮氏叫姑鱼(Johniusbelengerii)质量百分比较高,且夏秋季龙头鱼均为小黄鱼的主要摄食者[31],该群体数量虽较庞大,但其中56.67%的为龙头鱼幼鱼,多为当年生补充群体,一方面反映了当前该海域鱼类资源的现状,另一方面也揭示了该海域鱼类生物食物链的不稳定性。春、秋季主要鱼类的平均拥挤度及种间相遇机率计测结果表明,二者正相关性较强,即平均拥挤度较高的物种,种间相遇机率越高,反正则越低。这可能与二者对物种生态特征的衡量角度方面都注重物种的尾数及空间分布的集中程度有关。
3.2 幼鱼资源养护及增殖效果分析
当前海洋渔业资源衰退对渔业捕捞生产造成的刚性约束已经显现[32],而海洋渔业资源存量是从事捕捞生产的前提条件,渔民对渔业资源状况的认知(特别是幼鱼保护意识)对其捕捞生产具有决定性作用[33]。研究表明渔获量的增加一部分是靠大量捕捞幼鱼等手段获得,资源并没有明显好转[34]。长期以来张网、拖网等作业方式造成多数鱼类处于生长型过度捕捞,即对当年生幼鱼群体的大量捕获,导致大多数鱼未能达到最优化产量的捕捞状况,长此以往可能会转变成补充型过度捕捞[14],对资源恢复十分不利。目前休渔制度有效促进了东海区鱼类亲体和幼鱼补充群体的增加[35],对主要渔业资源起到了很好的暂养、增殖效果,为幼鱼群体生长提供有利条件,提高了补充群体的存活率,但过高的捕捞努力量使暂养的资源开捕后短期内被捕捞消耗,产卵亲体严重不足[36]。
以秋季主要鱼类资源量及幼鱼占比情况为例(表6),幼鱼成活率参照汪伟洋等[8]的研究取值50%,可见秋季主要鱼类的幼鱼资源量若能采取有效的养护措施,其一年后可提供较多的增殖补充资源量,特别是龙头鱼(A04)、鱼免鱼(A06)及银鲳(A07)的幼鱼资源量较大,若采取有效的幼鱼保护措施,其1龄鱼随着鱼体的增大,可增加海域内资源总量,其商品价值也将提高数倍。基于此,降低现存捕捞强度,强化幼鱼补充群体保护,有助于合理养护渔业资源,在完善伏季休渔制度的基础上,可采取相应措施形成一套科学合理的幼鱼资源养护及管理体系,在合理评估海域资源承载力的基础上,针对大黄鱼、小黄鱼、银鲳等常规品种,依据其原有生态栖息地开展增殖放流工程和人工鱼礁建设,实现幼鱼补充群体资源修复,增加渔业资源量。
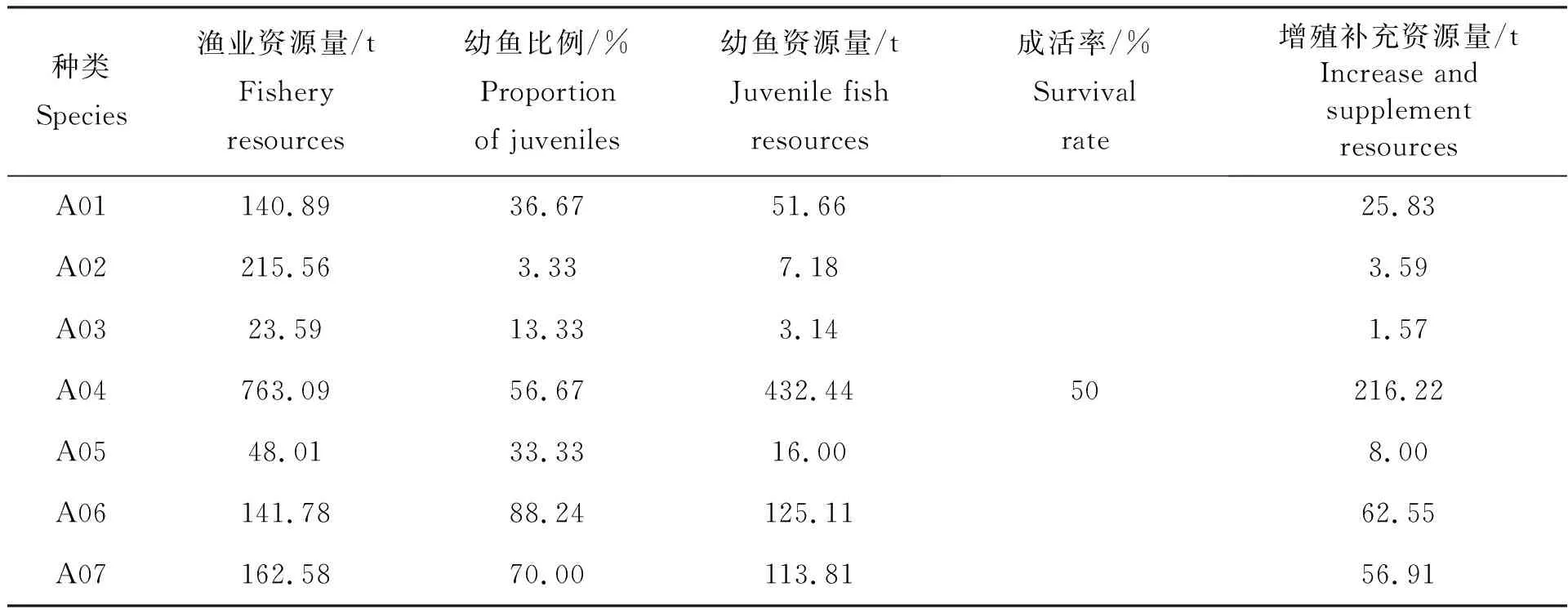
表6 秋季主要鱼类资源量及幼鱼占比情况
3.3 幼鱼保护与渔业管理
研究海域主要鱼类季节更替率较高,这可能与该海域春、秋两季鱼类洄游种类较多有关,主要鱼类组成季节性变化较大。而主要鱼类幼鱼所占比例春季较高的原因可能是因为春季是大多数鱼类繁殖产卵期,秋季物种体重、体长及幼鱼所占比例指标均优于春季,说明实施伏季休渔效果显著,而东海区鱼类如小黄鱼、银鲳等繁殖高峰期多为4月至6月,带鱼繁殖高峰期可至8月[19]。本研究春季主要鱼类幼鱼所占比例均较高且尾数较多,物种密集程度高,说明在该海域,大部分鱼类种群可能在春季的3月至4月即开始繁殖,生境场内已有大量幼鱼出现,该结论也与赵传絪[37]的研究结论相符,因此当前尚需探究科学可行的伏季休渔调整方案,提前伏季休渔开始时间,加大对禁渔前期产卵群体及幼鱼群体的保护,这一方面已有学者提出对现行伏季休渔制度实施时间进行调整[38-39],以保护春季产卵后补充群体的生长发育。春季六丝钝尾鱼叚虎鱼与秋季龙头鱼的平均拥挤度及种间相遇机率均为最高,反映了其物种资源量大、分布集中程度高、种群补充能力强的特征,一方面是由于两者当年生补充群体数量庞大,另一方面还可能与该海域适合其种群产卵繁育有关,推测长江口海域是龙头鱼的产卵场及索饵场,该结论与罗海舟等[39]的研究相符。其他经济价值较高的物种如凤鲚、银鲳及鱼免鱼等的平均拥挤度和种间相遇机率均不高,即这些物种在本海区内的分布较少,资源衰退较为严重。
本研究中5月幼鱼比例高于11月,即春季休渔期前有较多鱼类为快速生长的幼鱼,进一步说明了从2017年开始浙江省近岸定置张网休渔提前至5月的合理性,而秋季仍有大量幼鱼分布,即大量鱼类体长体重仍低于可捕标准,因此一方面沿岸渔场的生态养护亟需强化,舟山近岸海域为银鲳、小黄鱼等经济鱼类的产卵场和索饵场[40],修复这一海域的生态环境可提高渔业资源的再生能力;另一方面可探索将舟山沿岸渔场列为重要渔业水域保护区,进一步缩减近岸小型张网、刺网的捕捞强度,严格执行可捕标准,约束渔业生产者的捕捞行为,继续巩固“一打三整治”成果,并形成幼鱼保护长效机制。
当前浙江沿岸小型幼、杂鱼大多被出售给水产养殖企业作为鲜饲料投喂,而夏秋两季是养殖业的投饵旺季[38],在5—11月开展幼鱼保护工作,需强化对主要经济鱼类幼鱼的捕捞监管,加强渔业监管执法力度,一是加强伏季休渔期间的渔政管理,严格实施最小网目尺寸的监管措施等;二是完善新的渔业法规,针对不同作业方式制定不同的休渔方案及渔获物幼鱼比例规定,有针对性地制定繁殖季节幼鱼保护休渔制度及产卵场幼鱼养护制度,使幼鱼保护有法可依,促进该项举措逐步走上法制化管理、渔业资源合理性利用的良性循环之路;三是研发配合饵料以减轻水产养殖行业鲜活饲料投喂,增加捕捞网具幼鱼逃逸率等,亦需在海洋渔业资源保护与海洋捕捞业可持发展中优先考虑幼鱼资源保护,逐渐提高海洋渔业参与者的幼鱼保护意识。
猜你喜欢
当代水产(2022年8期)2022-09-20
当代水产(2022年7期)2022-09-20
养生保健指南(2018年4期)2018-04-11
价值工程(2017年8期)2017-03-25
法制与社会(2016年34期)2017-01-14
河北渔业(2016年12期)2017-01-11
养生保健指南(2016年12期)2017-01-06
河北渔业(2016年11期)2016-12-16
现代农业科技(2009年19期)2009-03-20
