
关中地区常绿阔叶植物叶片截留特征
2021-04-27 05:38:38孙雅婕刘建军
水土保持研究 2021年3期
孙雅婕, 刘建军
(西北农林科技大学 风景园林艺术学院, 陕西 杨凌 712100)
海绵城市的提出背景是日益严重的城市水资源问题。意图通过渗、滞、蓄、净、用、排的方式来缓解城市雨季内涝、旱季缺水的问题,提高城市对雨洪管理的自我调节能力[1]。林冠截留在促进雨水再分配和生态环境保护方面具有重要意义[2]。在森林林冠截留的基础上,部分学者关注园林植物林冠层对雨水的截留作用[3-5]。植物叶片与林冠截留能力之间的联系已被证实[6],并且与林冠截留的测定相比,单叶片水分截留测定更加便捷、灵活,因而探究叶片截留量对推断林冠截留具有意义。
叶片对水分的截留主要是通过表面截留及内部截留。叶片对水的亲和性不同,即叶片的润湿性不同,因此水分在叶片表面的展开形式不同。水分在亲水性叶片表面铺展为水膜,叶片与水分间的相互作用较强。水分在疏水性叶片表面通常以水珠的形态存在,容易受到风力、重力的影响而滴落[7]。不同植物叶片的微观几何结构、叶表面的化学组成不同,因此表面截留能力不同[8]。内部截留是指植物叶片利用角质层、毛孔或排水器来吸收水分,利用树冠截留水分的现象[9]。不同植物叶片结构特征、生理特性不同,内部截留速率不同,最大截留能力也不同。
部分学者利用浸泡法、喷水法、人工降雨法等多种方式对单叶片进行截留持水量测定[10-11]。然而,有关园林单叶截留能力的研究较少,且相关报道多集中于叶片形态因子,如叶面积、叶型、叶长、叶宽、粗糙度等[12-14],较少从植物生理功能进行探讨。叶片干物质含量(leaf dry matter content,LDMC)可以准确反映出植物生态行为差异及获取资源的能力,比叶面积(specific leaf weight,SLA)能够反映出不同植物为适应不同环境条件而产生的生长状况差异[15]。二者是已证描述叶片功能性状及其与生态系统关系的两个最优指标,可以解释90%的植物叶片与环境之间的相互作用[16]。为深入了解植物生理功能与环境适应潜力之间的内在联系,本研究在探究形态因子与单叶截留量的基础上,深入探讨LDMC,SLA与叶片截留持水的关系,以解释叶片性状在水文方面对环境的适应性。
西咸新区地处陕西关中地区,是国家第一批海绵城市试点城市之一[1],初步形成了符合低影响开发理念的设施体系,但在植物种类选取及配景上仍处于探索阶段[17]。海绵城市建设离不开绿地建设,海绵绿地的建设不仅要满足景观艺术化的需求,更应考虑植物的生态效益。选取水分截留效果较好的植物,形成一个个“小海绵”,可以促进城市对雨水的调节作用。已有研究证明,常绿植物的雨水截留量较落叶植物强[18]。与针叶树种相比,常绿阔叶树种形态多样、色彩丰富,更有利于营造具有观赏美感的植物景观。关中地区的常绿阔叶树种不少,最常见的有:广玉兰、石楠、桂花、枇杷、南天竹等[19]。对常绿阔叶植物加以合理选择及应用,有助于打造“景观美”与“生态美”兼顾的植物景观群落。因此,本研究依据植物生长状况,选取关中地区16种常见的常绿阔叶植物作为对象,利用浸泡法模拟降雨截留过程,研究不同植物叶片的截留能力,及其与叶性状因子之间的关系。为营造水分截留效果较好的园林植物种类的选取提供参考,以期利用园林植物的截留作用来缓解城市中水资源问题,促进海绵城市建设。
1 试验材料与方法
1.1 试验材料
试验所采集的叶片皆为晴天干燥的同一时间段采摘。每个树种选取3株生长态势良好的中龄树,在其树冠的不同方位处采样,随机摘取大小不一的完整成熟叶片,叶片较大树种每棵取10片,叶片较小树种每棵取20片,共540个叶片样本。记录不同植物类型的生活习性及叶面特征见表1。

表1 被试植物生活习性及叶面特征
1.2 试验方法
已有研究证明,植物叶片吸水量与浸水时间为自然对数关系,极短时间内叶片吸水量几乎可以忽略不计,吸水1 h内为快速增加期,截留速率最大,1 h后缓慢增长至饱和状态,4 h达到截留量峰值后不再变化[20]。因此,叶片进入水中5 s后可将内部截留量忽略不计,增加水量为叶片表面截留水(Is)。测定吸水过程中截留速率变化的重要时间拐点——1 h后截留量(It)。为方便LDMC的测定,本研究测定浸水24 h后截留量来探究不同植物叶片内部对实际降雨的最大截留能力(Imax)。叶片截留量皆采用浸泡称重法测定,计算如式(1),(2),(3) ,具体操作为:利用电子天平先测量每个样品的鲜重M1(g),再用镊子将单个样品完全浸没于装有自来水的圆桶或圆盆中,若叶片表面有较大气泡将其破坏,浸没5 s后迅速取出称重记为M2(g),随后将叶片完全浸入水中充分吸水,分别于吸水时间1 h后和24 h后进行称重记为M3(g)和M4(g)。每次称量前,将叶片取出水后悬垂至无明显水滴滴落再进行测定。完成24 h浸水测定后的叶片放入85℃±5℃的烘箱中至质量恒定,称重记为M5(g)。
记录叶片形态因子为生长型、叶质、叶平滑度、叶毛、叶面积、叶厚、叶长、叶最大宽。其中,生长型按乔木、小乔木、灌木分别记为1,2,3;叶质按厚革质、革质、纸质分别记为1,2,3;叶面光滑度按叶皱、微皱、上表面光滑、光滑分别记为1,2,3,4;叶毛按照无毛、叶背密生毛、两面被毛分别记为1,2,3。叶片面积、长、宽、厚利用LI-3000C叶面积仪、电子游标卡尺测定;记录叶片功能因子为SLA,LDMC,具体计算如式(4),(5) 。
IS=M2-M1
(1)
It=M3-M1
(2)
Imax=M4-M1
(3)
SLA=LA/10M5
(4)

(5)
式中:SLA表示比叶面积(m2/kg);LDMC表示叶片干物质含量(%);Is表示5 s(表面)截留量(g);It表示1 h截留量(g);Imax表示24 h(内部)最大截留量(g);LA表示叶面积(cm2);M1表示叶片鲜重(g);M2表示浸水5 s后重(g);M3表示浸水1 h后重(g);M4表示浸水24 h后重(g);M5表示叶片烘干重(g)。
利用Excel 2019进行数据整理计算,SPSS 23.0统计分析软件对数据进行单因素方差分析、相关性分析、主成分分析(PCA)、回归分析以及聚类分析。
2 结果与分析
2.1 不同植物叶片水分截留能力差异
由图1可知,同一植物在5 s,1 h,24 h的截留量依次递增,且不同植物间增加量具有明显差异。广玉兰、皱叶荚蒾、枇杷的叶片截留量显著高于其他植物。其中,在不同时间段广玉兰的截留量始终为最大。与枇杷相比,皱叶荚蒾的Imax较大,但Is,It值较小,由此可见短时间内,枇杷的截留效果比皱叶荚蒾强。除上述三者外的其他植物,不同植物的叶片截留量差异较小,且同一植物Is,It,Imax间的增量较小,即在短时间内基本达到最大截留能力,截留水量基本达到饱和状态。24 h内,部分植物截留水量间差异性发生改变(表2)。如:蚊母树与十大功劳间、北海道黄杨和枸骨间、南天竹和枸骨间、珊瑚树与枸骨间的Is,It差异显著,Imax差异不显著,即表面润湿性不同,叶片内部截留速率不同,但24 h内部最大截留能力差异不显著。枸骨与红叶石楠间的Is,Imax差异显著,It差异不显著,即与红叶石楠相比,枸骨叶片表面的亲水性更好,24 h内的截留能力更大。但1 h内,枸骨与红叶石楠的截留速率差异不显著。皱叶荚蒾和箬竹间Is,It差异不显著,但Imax差异显著。即1 h内叶片内部截留速率差异不大,但24 h叶片内部最大截留量大小有差异。由于本试验从1 h到24 h的时间跨度较大,因此无法详尽得知二者间截留量差异变大的具体时间节点。除上述外,24 h内其余植物间的差异性没有发生改变,总体表现为大叶片与小叶片间截留量差异显著,叶型相似的植物叶片间截留量的差异不显著。
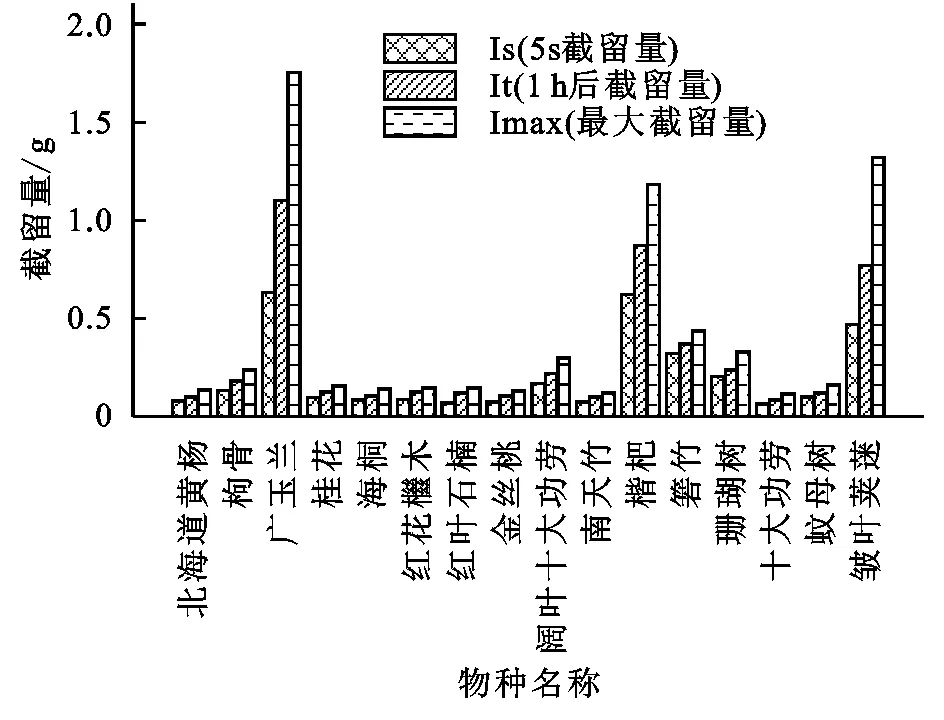
图1 16种植物叶片截留量
2.2 叶片性状因子与截留能力之间的关系
It,Imax皆为叶片内部截留能力的特征值,二者间相关性极显著(p<0.01,r=0.963)。因此,仅对Imax与叶片性状因子进行相关性检验,以探究叶片在实际降雨中内部截留的最大潜力。叶片表面截留量、内部截留量与叶片性状因子之间的相关性检验见表3。Is与叶面积、最大宽、叶长、平均叶厚、叶毛呈极显著正相关(p<0.01),Is与生长型、叶质、叶片光滑度呈极显著负相关(p<0.01)。即厚革质、叶皱、两面被毛、叶形态较大的乔木截留能力最佳,这与图1的结果相吻合。SLA与Is间呈极显著正相关(p<0.01),LDMC不影响叶片短时间内的表面截留能力。总体上叶片功能因子对叶片表面截留量的影响程度小于叶片形态因子。除SLA与Imax间呈显著正相关(p<0.05)外,Imax与叶面积、最大宽、叶长、平均叶厚、叶毛呈极显著正相关(p<0.01),与生长型、叶质、叶面光滑度、LDMC呈极显著负相关(p<0.01)。即叶片的形态因子、功能因子皆对叶片内部最大截留能力有影响,且形态因子中叶片大小的影响更大。与叶片表面截留量相比,叶片内部截留量受叶片功能因子的影响更明显。

表2 植物叶片截留量及差异性

表3 主要叶片性状因子与截留能力的相关性分析
为进一步探究植物叶片性状因子对水分截留能力影响程度的大小,对其与Is,Imax进行PCA分析。由表4可知,累积贡献率的76.936%来自前3个主成分,即前3个因子模型可以解释76.936%的试验数据,包含了大多数的信息,且前3个因子特征根皆大于1,因此本试验提取前3个因子来判断叶片性状对截留率的影响。由表5可知,第1主成分中叶面积、最大宽、平均叶厚、叶长荷载分别为0.895,0.869,0.784,0.741,反映了叶面的形状特征。第2主成分中LDMC的荷载为0.756,反映了植物叶片对养分的保持能力。第3主成分中SLA的荷载为0.874,反映了叶片光合、蒸腾等生理特征以及对不同环境的适应能力。

表4 PCA中总方差解释
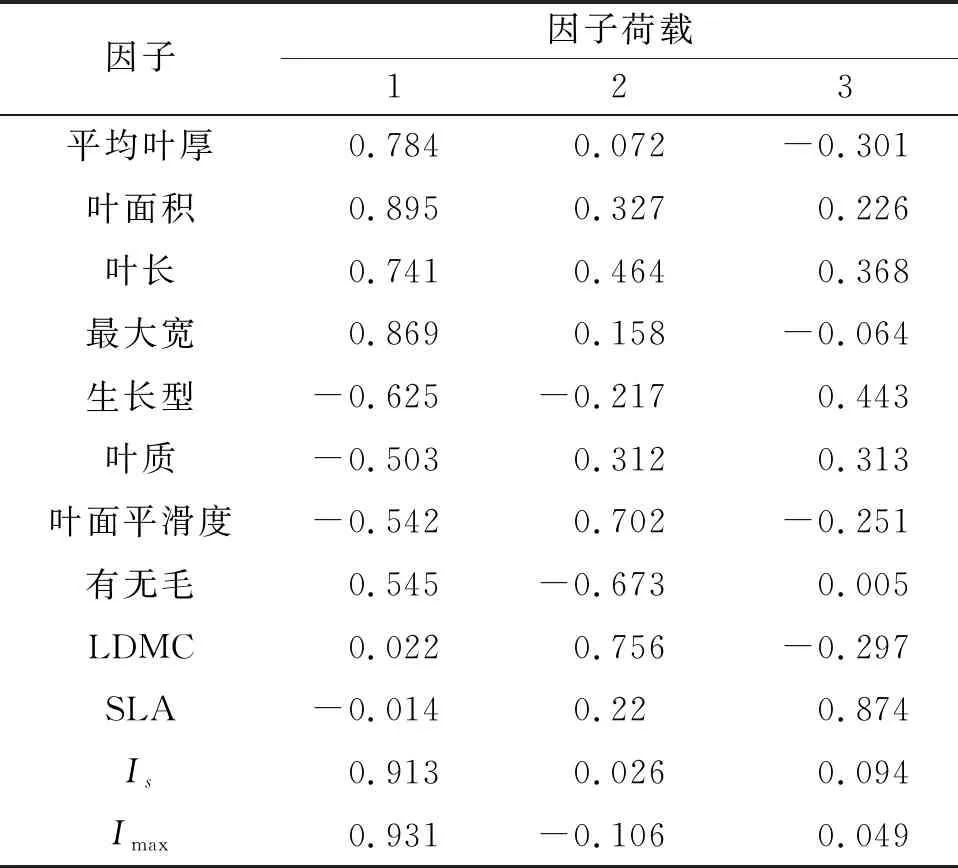
表5 各因子荷载
综合相关性分析及PCA分析结果可知,对叶片截留能力具有影响的主要因子有叶片大小特征(叶面积、最大宽、叶厚、叶长),叶片功能特征(SLA,LDMC),且叶片形态因子影响较大。基于叶片形态因子及叶片截留量的实测数据进行函数拟合。由初步拟合结果看,叶片形态因子与截留量间关系对线性函数、对数函数、二次函数、三次函数、复合函数、幂函数皆具有统计学意义(p<0.01),依据相关系数大小,采用拟合效果较好的复合函数(表6)。叶片表面截留量与内部截留量皆呈现随叶面积、平均叶厚、叶长、叶最大宽增大而增大的变化趋势,但增长速度不同,总体为先缓后快的增长趋势。叶面积<75 cm2,平均叶厚<1 mm、叶长<25 cm、叶最大宽<6 cm时,叶片截留量增长速度较慢,随后增长速度较快。

表6 主要性状因子与叶片截留量的回归方程
基于16种植物叶片的截留量及筛选出来的叶片性状主要影响因子为属性,采用欧氏距离为度量进行系统聚类,得到16种植物叶片性状与截留能力共同影响下的聚类树状图(图2)。取阈值为9,则可将16种植物分为4大类。叶片形态最大、LDMC较小、SLA中等、高截留功能的乔木:广玉兰。叶片形态较大、LDMC较小、SLA中等、较高截留功能的灌木或小乔木:箬竹、枇杷。叶片形态较大、LDMC较小、SLA较大、较低截留功能的灌木:皱叶荚蒾、珊瑚树。叶片形态较小、LDMC较大、SLA较小、低截留功能的灌木或小乔木:十大功劳、海桐、南天竹、北海道黄杨、金丝桃、红花檵木、阔叶十大功劳、枸骨、桂花、蚊母树、红叶石楠。
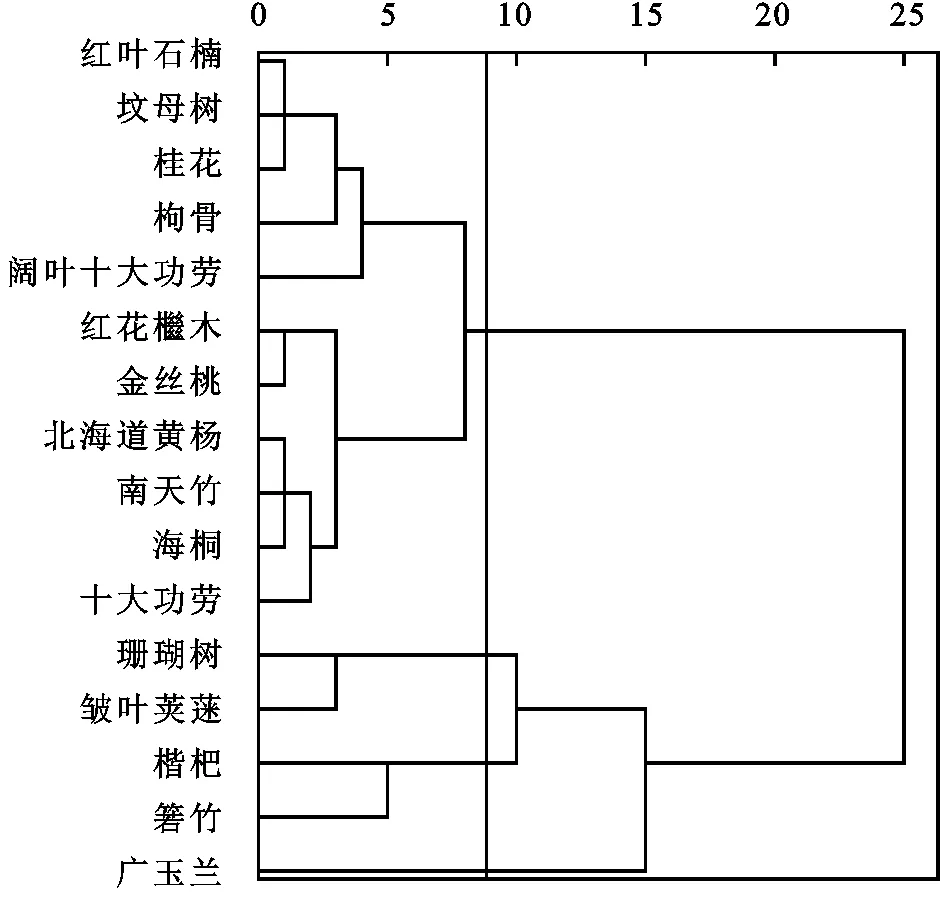
图2 截留能力-叶片性状聚类分析
3 讨论与结论
(1) 本研究中对叶片截留能力影响最大的叶片性状因子有叶面积、叶长、叶宽、叶厚、SLA及LDMC。有研究证明叶面积大小[10]、叶长[14]、叶宽[21]对叶片截留能力的影响显著,本研究中广玉兰、枇杷、箬竹、皱叶荚蒾的单叶截留量较大,与已有研究的结果相同。叶片形态越大,则与水分的接触面越大,因此截留量越大。有关叶厚对截留能力影响的研究较少。叶片纸质的箬竹虽叶片厚度较小但单叶截留量较大,这可能是因为箬竹的单叶面积在16种植物中仅次于广玉兰,属于叶面积较大植物。除箬竹外,总体而言,本研究中叶片厚度与截留能力为显著正相关,符合植物通过增加叶厚来适应干旱强光的环境,进而达到降低蒸腾作用,减少组织内部水分散失,提高水分利用率的现象[22]。
除本研究中PCA筛选出的主要叶片形态因子外,叶毛在前人的结论中被认为也是影响叶片截留持水量的重要因子[13-14]。不同的绒毛类型影响着水滴在叶表皮的存在形式,进而使叶片的湿润性也有所差异[23]。仅以有无来评断叶毛对截留量影响的局限性,以及被试物种的差异可能是本研究与他人研究结果不同的原因所在。
(2) 植物叶片功能因子与水分截留的耦合作用,反映了植物适应环境条件的自我调控机制。水分作为光合作用的重要影响因素之一,不仅直接参与光合作用过程,更是通过影响叶绿素合成等间接影响叶片的光合能力[24]。叶片含水量降低,则光合速率增大,水分利用率增大[25]。植物叶片光合作用能力直接影响干物质积累,光合作用越强,LDMC越小,植物叶片的组织密度较低,减小了水分向表面扩散的距离及阻力[26],内部水分散失的增大增加了植物对水分的需求,叶片截留持水能力增加,因而LDMC与Imax间呈负相关。就SLA而言,代表着一定干物质投资所展开的捕光表面积数量,是反映植物相对生长速率的因子[27],这可能是本研究中SLA与截留量相关关系较弱的原因。叶片内部生物量分配是对环境的响应。SLA较高则植物单位生物量的面积较大,叶片吸收水分的表面积较大,因此有利于叶片吸水[20]。SLA较低的植物为降低蒸腾作用,避免过度失水而形成面积较小、较厚或组织密度较大的叶片[28]。因此,为验证SLA与叶片截留能力的作用,进一步探究SLA与被试16种植物的单叶截留率以及平均叶厚的关系,结果显示SLA与单叶截留率呈明显的正相关,与平均叶厚呈明显的负相关,符合较小的SLA可以降低内部水分丧失,提高水分利用率的现象。虽然高SLA的植物叶片对于雨水的截留能力较强,但高SLA的植物生命周期相对较短且对不良环境的适应性更差[29]。这可能是本研究中高SLA树种(皱叶荚蒾、珊瑚树)的单叶截留量反而小于中等SLA树种(广玉兰、箬竹、枇杷)的单叶截留量的原因。
(3) 叶片对雨水截留的利用表现为两方面。其一为叶片利用表面润湿性,使水分在叶表面铺展,即叶片表面截留水分。再通过蒸腾蒸发作用使水分返回生态系统,从而实现对水分的再分配,达到增加湿度、促进大气水循环的生态作用。其二是叶片表面的微形态结构及其内部的亲水化合物可以帮助叶片吸收水分,不仅可以截留水分,也可以改变叶片本身的水势,提高叶片的含水率[30]。再通过茎叶间的水势差,使多余水分由叶片进入树种、茎干甚至根部土壤,实现水分从大气到植物根际的逆向运移过程。叶片的内部截留量对植株整体的生理过程及环境适应性产生影响[9]。基于此,本研究探究了叶片性状因子与表面截留量、内部截留量的关系,综合二者来表现叶片性状因子与叶片截留能力的关系。
(4) 通过对关中地区16种常绿植物叶片性状及截留能力的定性定量分析,得出叶片形态较大、LDMC较小、SLA中等水平的植物截留能力最佳,即广玉兰、箬竹、枇杷等。但本研究中截留量测定的时间跨度较大,因此未能准确反映植物叶片截留速率变化的具体时刻差异。后续研究中可细化时间间隔,并进行一年四季的测定,以得到较为完整的叶片截留变化态势。此外,本研究探讨的叶片形态特征仅为宏观形态,可以从叶片微观结构层面深入探究叶片内部不同结构类型与水分截留的具体关系。
猜你喜欢
科普童话·百科探秘(2023年5期)2023-06-19 04:18:30
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02 01:48:06
世界科学技术-中医药现代化(2020年2期)2020-07-25 02:06:08
新疆农业科学(2020年1期)2020-02-14 03:22:46
特别健康(2018年9期)2018-09-26 05:45:42
临床医药文献杂志(电子版)(2017年11期)2017-05-17 04:47:56
青苹果·教育研究版(2016年7期)2016-12-12 05:19:52
中学生物学(2016年8期)2016-01-18 09:08:21
中国麻业科学(2015年5期)2015-12-28 06:22:11
高中生学习·高三版(2014年3期)2014-04-29 06:12:54
