
改变凋落物输入对川西亚高山天然次生林土壤呼吸的影响
2021-04-25 06:26李亚非胡宗达刘兴良罗明霞欧定华吴德勇
生态学报 2021年7期
李亚非,胡宗达,*,余 昊,胡 璟,刘兴良,罗明霞,欧定华,吴德勇
1 四川农业大学资源学院, 成都 611130
2 湖南农业大学资源环境学院, 长沙 410128
3 四川省林业科学研究院, 成都 610081
森林土壤作为碳储存的重要组分,在森林生态系统中扮演着十分重要的角色。土壤碳排放是连接土壤碳库和大气CO2重要纽带,每年向大气排放的约50 Pg C来自凋落物和土壤有机质的分解[1]。凋落物作为影响土壤呼吸的一个重要因子,分解速率因其质量和数量不同而异[2],对土壤呼吸速率的影响也就不尽相同[3-4]。在全球尺度上,凋落物对土壤呼吸的平均贡献率约为20%—30%[5],而温带地区森林植被地表凋落物的贡献则为15%—31%[6-9]。有研究显示,凋落物量与土壤呼吸速率正相关或不相关[10-11],凋落物的去除和添加可降低或增加土壤呼吸速率[7,12-13],但增减幅度与凋落物输入量并不完全同步[13-15],一方面源于地上凋落物输入量增加可能提升现存有机质的分解速率[16]或对其分解速率没有显著影响[17];另一方面由于去除和添加凋落物改变了表层土壤碳源的供给水平和土壤的水热条件[14,18]。可见凋落物输入量对土壤碳排放的影响较为复杂。此外,不同林型的土壤温湿度、地上生物量、根系周转、土壤有机碳氮及其活性组分储量的不同,其土壤呼吸的响应也不尽相同[19-21]。
次生林作为川西亚高山林区的主要森林类型之一,在水源涵养、气候调节、水土保持等方面起着重要作用。近年来,人们对川西亚高山森林土壤呼吸进行了大量研究[22-25],发现土壤呼吸速率主要受控于温度的变化,认为全球气候变暖可能会导致该区域森林土壤呼吸速率增加,但不同森林类型的碳排放量高低各异[26-27],主要由于凋落物量、地形以及微气候等的差异所致[9,27-28]。因此,有必要进一步探究该区域土壤碳排放对不同林型的响应及其影响因素。有研究表明,次生针阔混交林和次生桦木林凋落物储量分别为4.10 t/hm2和3.52 t/hm2,凋落物分解速率较快有利于土壤养分的补充[29],表明地上凋落物可能是该林区土壤呼吸的重要来源。然而关于凋落物对土壤呼吸影响的研究主要针对粗枝云杉(Piceaasperata)人工林[30],对天然次生林尤其是相同母质及气候背景下地表凋落物对土壤呼吸影响的相关量化研究至今未见报道。鉴此,本研究在川西亚高山米亚罗林区,选择20世纪60年代采伐后经自然更新恢复形成的天然次生林,监测3种处理(对照、添加和去除地表凋落物)下的土壤呼吸速率及其相关环境因子,拟解决以下科学问题:(1)分析生长季内去除和加倍地表凋落物对土壤呼吸的影响程度;(2)探究控制土壤呼吸的主要影响因素;(3)评估凋落物对天然次生林土壤呼吸的贡献率。本研究有助于了解环境条件相似背景下天然次生林凋落物对土壤呼吸及其温度敏感性的影响程度,也可为天然次生林土壤碳库管理提供参考。
1 研究区概况
研究区位于四川省阿坝藏族羌族自治州理县米亚罗林区,海拔范围2200—5500 m,属于青藏高原—四川盆地的过渡地带。气候受高原地形影响,具有典型的高山气候特征。年均温度6—12℃,1月份平均气温-8℃,7月份平均气温12.6℃,年均降雨量700—1400 mm,集中于5—10月的生长季,森林土壤类型以山地棕壤为主。
实验样地位于理县夹壁乡下辖的夹壁沟天然次生林林区(102°50′20″ E, 31°37′15″ N),海拔范围3113—3172 m,土壤母质和气候背景相似。林型主要是青榨槭(Acerdavidii)+红桦(Betulaalbo-sinensis)+陕甘花楸(SorbuskoehneanaSchneid)次生阔叶林(简称阔叶林)、红桦+青榨槭+岷江冷杉次生针阔混交林(针阔混交林)和岷江冷杉(针叶林)等3种天然次生林。主要物种有青榨槭(Acerdavidii)、红桦(Betulaalbo-sinensis)、岷江冷杉(Abiesfaxoniana)、青荚叶(Helwingiajaponica)、陕甘花楸(SorbuskoehneanaSchneid)、野樱桃(Cerasusszechuanica)、忍冬(Lonicerajaponica)、鞘柄菝葜(Smilaxstans)、苔草(Carexdoniana)、山酢浆草(Oxalisacetosella)、卵叶韭(Alliumovalifolium)、掌叶橐吾(Ligulariaprzewalskii)、圆穗蓼(Polygonummacrophyllum)、冷水花(Pileanotata)等。本研究选取针叶林、针阔混交林和阔叶林3种林型为试验样地,样地基本概况见表1。
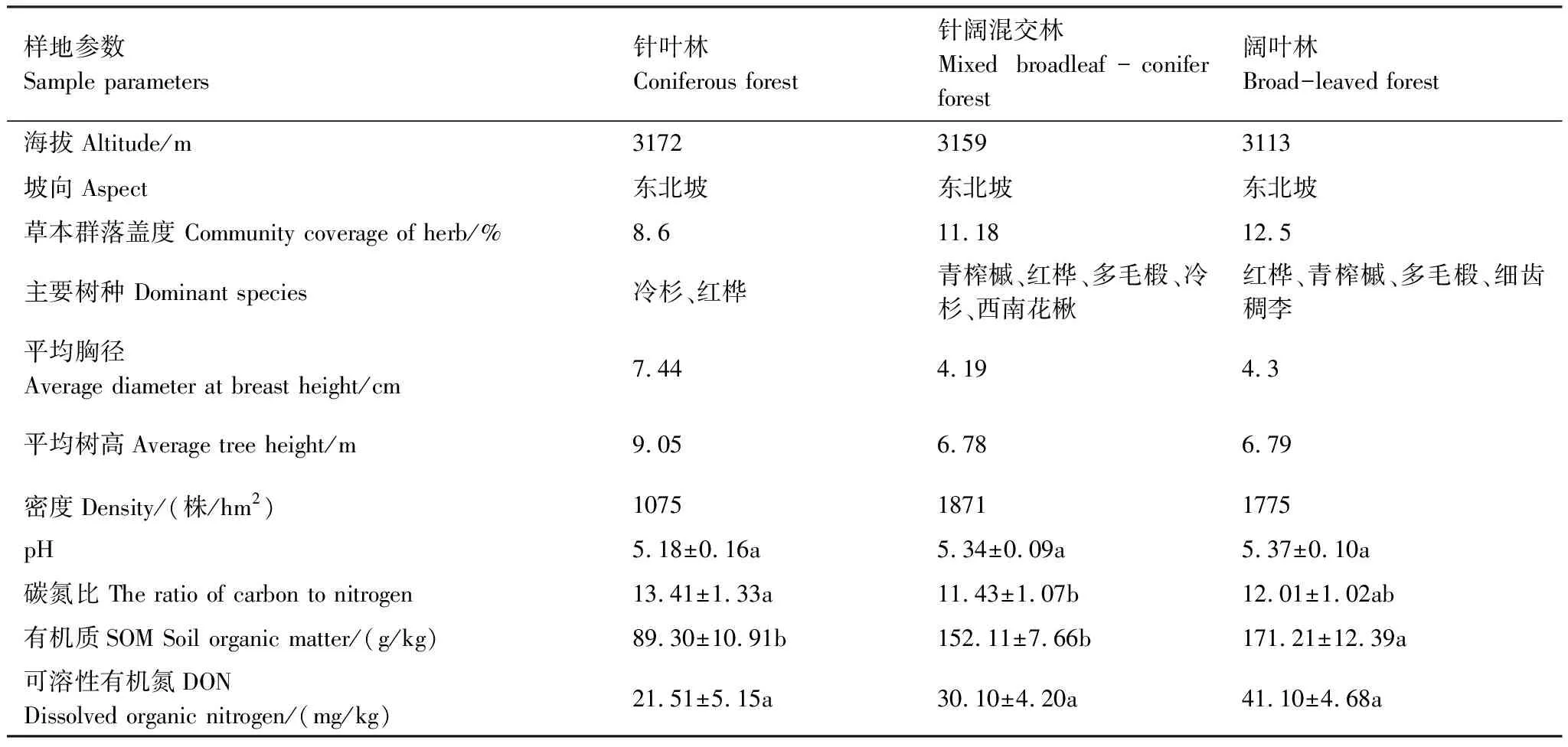
表1 样地基本概况
2 研究方法
2.1 试验样地设置
2018年8月,在针叶林、针阔混交林和阔叶林 3种林型中,按照相邻原则(即依据试验样地具体情况,样地间隔≥20 m;林型间距>20 m),用罗盘仪(DQL-1)分别设置3个投影为20 m×20 m的标准样方,在每个样方内随机设置3种处理,每种处理为1 m×1 m的小样方,即:(1)对照处理(CK),不做任何处理,保持原状;(2)去除凋落物(RL),清除小样方内全部地表凋落物;(3)加倍凋落物(DL),将去除凋落物处理小样方内的凋落物均匀添加到加倍凋落物的小样方中。每种处理3个重复,每个林型各设置27个小样方,并在去除凋落物小样方的上方0.5—0.8 m高处放置一个1 m×1 m的尼龙网,以阻止凋落物掉入小样方内。
2.2 土壤呼吸与土壤温湿度测定
每个小样方内埋设一个PVC环,其中去除掉落物处理的PVC环为直径20 cm×高8 cm;对照和加倍凋落物处理的PVC环为直径20 cm×高12 cm。所有PVC环平行地表插入土壤5 cm,监测期间PVC环的位置保持不变。于2019年生长季(5月—10月)利用LI-8100A土壤碳通量自动测量系统(LI-COR, NE, USA)测定土壤呼吸速率(Rs),每月测定2次(测定时间间隔约2周),每次测定尽量选取晴朗天气进行观测,测量时间为9:00—14:00。在测定土壤呼吸的同时,分别采用LI-8100A自带的温度探针与水分传感器(ML2x)同步测定土壤5 cm处的土壤温度(T)和湿度(体积含水量,W),每个观测点重复测定2次取平均值。此外,观测期内,因2019年8月下旬去往研究区域的道路因突发泥石流滑坡导致交通中断,故8月只在中上旬测定一次。
2.3 物种调查与土壤理化性质测定
利用生态学群落样方调查法,对每个标准样方进行调查,主要记录和测量各样方内乔木(胸径≥1.5 cm)的物种名称、盖度、数量、高度、群落盖度等,同时按照“梅花5点法”布设5个1 m×1 m小样方,记录草本植物名称、株数、高度等指标。此外,将20 m×20 m样地按照网格法划分为4个10 m×10 m的亚样方,用不锈钢土钻在每个亚样方内按照“梅花5点法”钻取0—20 cm表层土壤(共计20钻土样),充分混合后,按四分法取一个混合样装袋带回实验室,去除土壤中的石砾、根系和杂物,过2 mm土壤筛,每个土壤样品分为2份备用。
土壤有机质(SOM)含量采用重铬酸钾氧化-外加热法测定[31];土壤pH值采用电位法测定[31];土壤全氮(TN)含量采用采用硫酸-催化剂消解-流动分析仪法(AA3连续流动分析仪CFA,德国);土壤可溶性有机氮(DON)含量=可溶性总氮含量(DTN,紫外分光光度计测定)-可溶性无机氮含量(DIN,AA3连续流动分析仪测定)。
2.4 数据处理与分析
选用Margalef丰富度指数、Shannon指数、Simpson优势度指数、Pielou均匀度指数计算乔木层和草本层的物种多样性[32]。
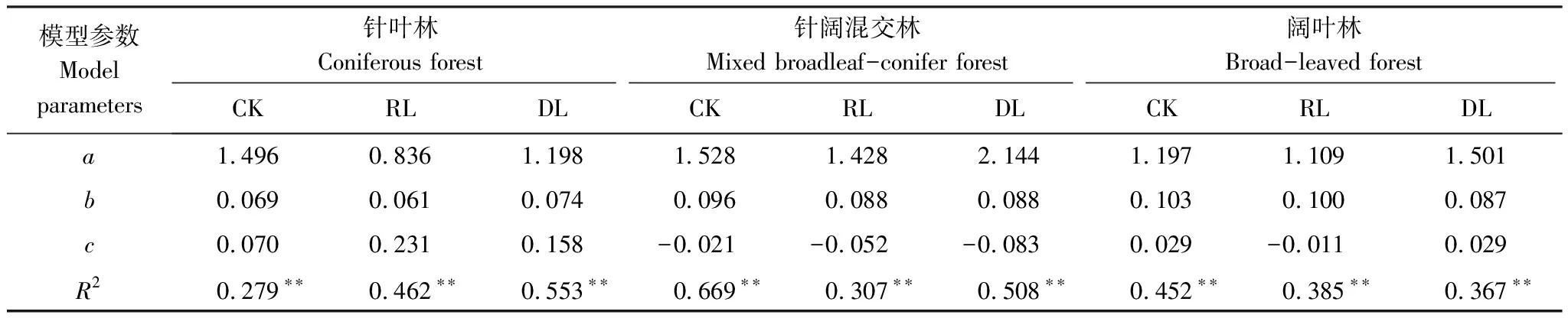
采用指数模型Rs=aebT、线性模型Rs=aW+b与双因素模型Rs=aebTWc分析观测期内土壤呼吸与土壤温度和湿度的关系,式中Rs为土壤呼吸速率(μmol m-2s-1),T为5 cm处土壤温度(℃),W为5 cm处土壤体积含水率(%),a、b、c为拟合参数。
温度敏感系数(Q10):Q10=e10b,式中b为土壤呼吸与温度拟合参数。
凋落物贡献率:Sl=(R-RE)/R×100%,式中Sl为凋落物贡献率,R为对照土壤呼吸速率平均值,RE为去除凋落物的土壤呼吸速率平均值。
利用单因素方差分析(One way ANONA)比较不同林型及处理间平均土壤呼吸速率与土壤温湿度的差异性;利用重复测量方差分析(Repeated measure ANONA)检验林型与凋落物处理的交互效应;用非线性回归(Regression analysis)分析土壤呼吸速率与土壤理化性质、物种多样性指数的相关性。数据统计分析采用SPSS 26.0,用Sigmaplot 14.0软件制图。
3 结果与分析
3.1 土壤温湿度变化
针叶林、针阔混交林和阔叶林3种林型中(图1),去除凋落物(RL)、加倍凋落物(DL)和对照(CK)的土壤温湿度均表现为先升高后下降的变化趋势(P<0.05)。从表2可知,同一林型中,3种处理间5 cm土壤温湿度差异性均不显著(P>0.05)。不同林型间相同处理的土壤温度亦无显著差异(P>0.05);而土壤湿度显著性差异表现不一,其中RL处理在不同林型之间未表现出显著差异性,DL处理在针叶林(19.55%)与针阔混交林(21.36%)间差异显著(P<0.05),CK在针阔混交林(23.37%)与阔叶林(19.96%)间差异显著(P<0.05)。
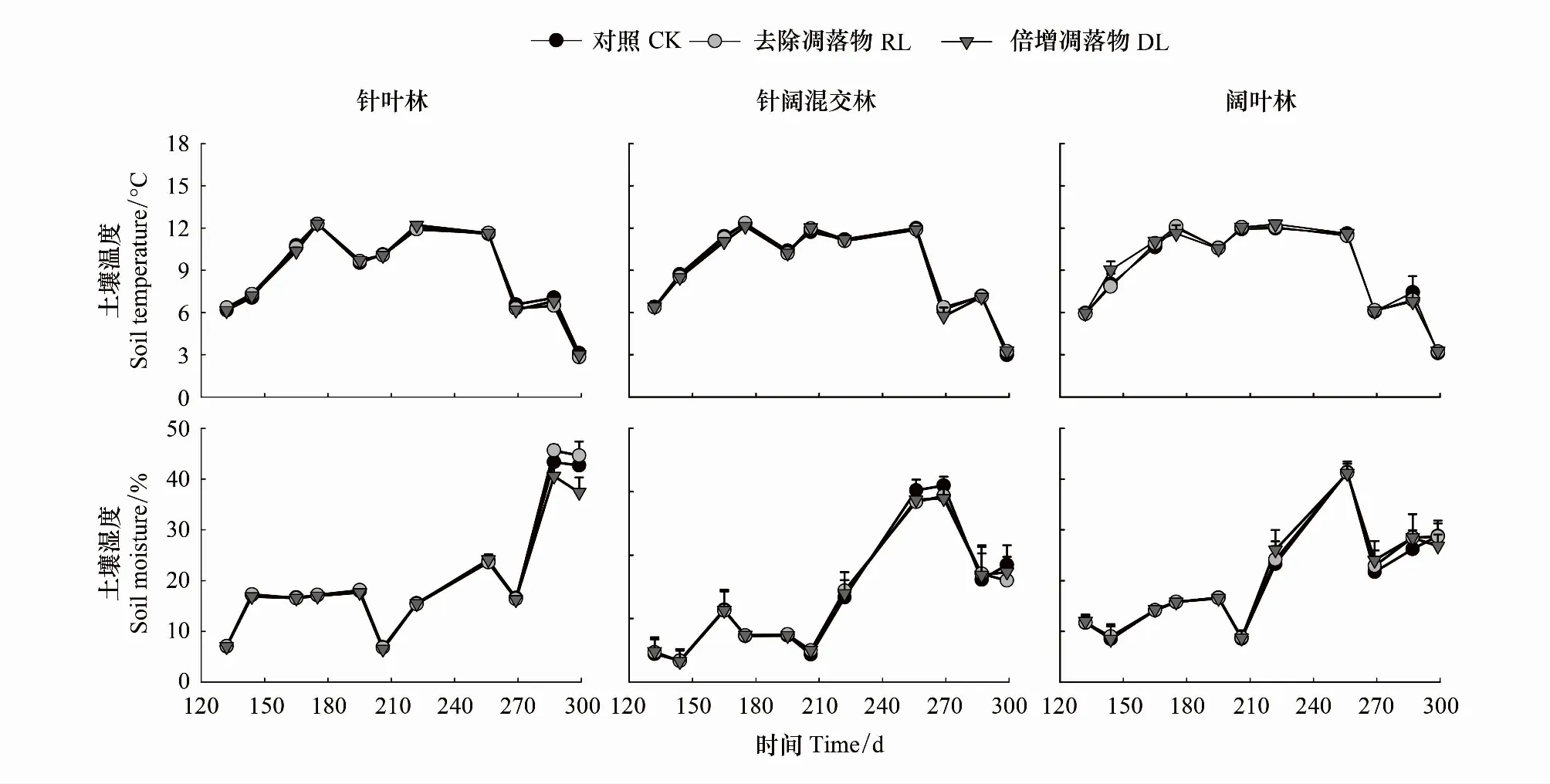
图1 川西亚高山天然次生针叶林、针阔混交林和阔叶林不同处理土壤温度和湿度动态变化
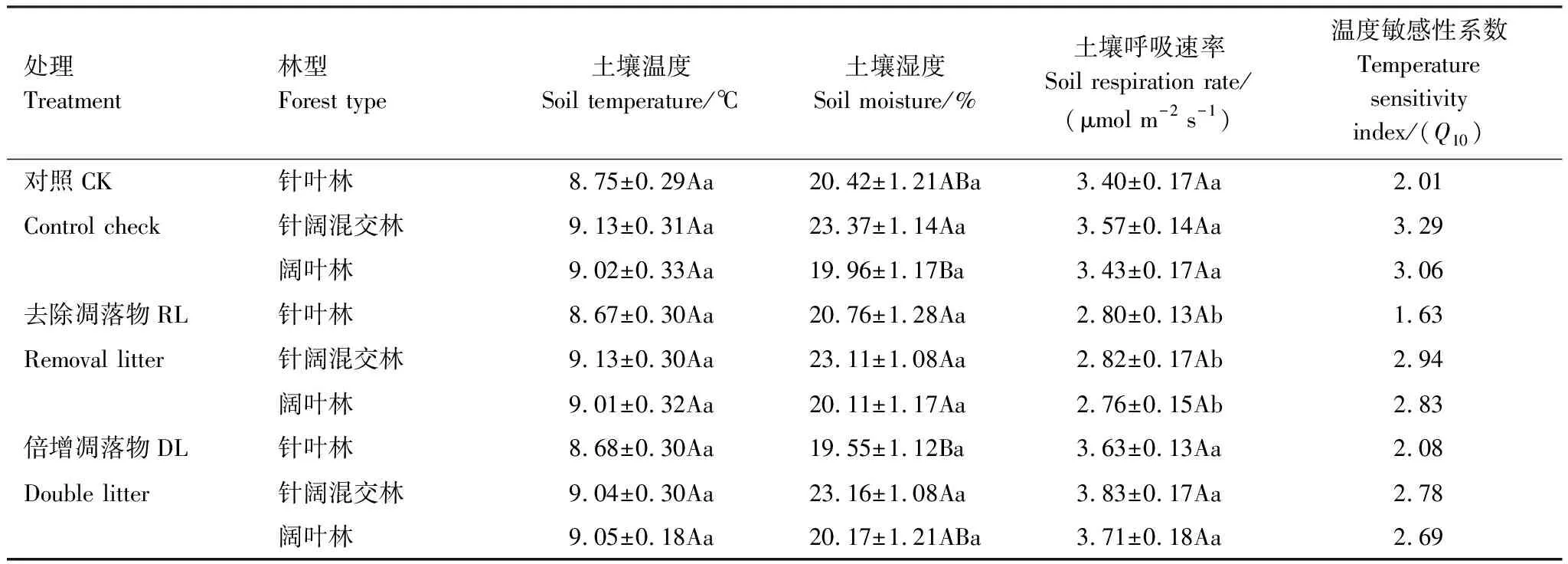
表2 土壤温湿度、呼吸速率均值与温度敏感性系数
3.2 去除和添加凋落物对土壤呼吸的影响
3种林型不同处理的土壤呼吸速率均与温度季节变化趋势基本一致(图2)。由表2可知,在3种次生林中,CK土壤呼吸速率与DL差异均不显著,而与RL均有显著差异(P<0.05),针叶林、针阔混交林、阔叶林土壤呼吸速率均表现为RL
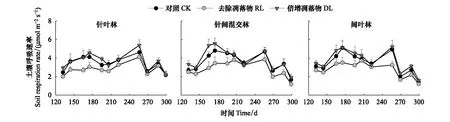
图2 针叶林、针阔混交林和阔叶林不同凋落物处理土壤呼吸动态变化
针叶林中CK与DL平均土壤呼吸速率分别为3.40 μmol m-2s-1与3.63 μmol m-2s-1,RL较CK降低0.60 μmol m-2s-1;针阔混交林中CK与DL平均土壤呼吸速率分别为3.57、3.83 μmol m-2s-1,RL较CK降低0.75 μmol m-2s-1;阔叶林中CK与DL平均土壤呼吸速率分别为3.43、3.71 μmol m-2s-1,RL较CK降低0.67 μmol m-2s-1(表2)。去除凋落物后土壤呼吸速率在针叶林、针阔混交林和阔叶林中分别降低了17.65%、21.01%、19.83%,而加倍凋落物的土壤呼吸速率则分别增加了6.76%、7.28%和8.16%。
重复测量方差分析(表3)可知,时间、凋落物处理对土壤呼吸均表现出显著的影响(P<0.05),而林型的影响不显著(P>0.05);林型与凋落物处理对土壤呼吸不存在显著的交互作用(P>0.05)。
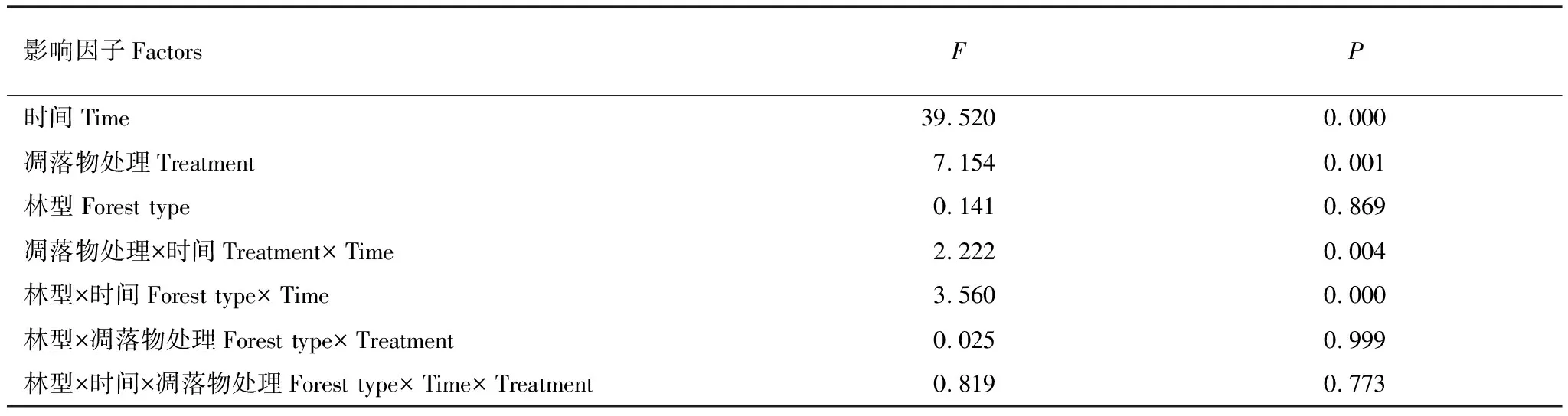
表3 土壤呼吸的重复测量方差分析
3.3 去除和加倍凋落物土壤呼吸速率与水热因子的关系
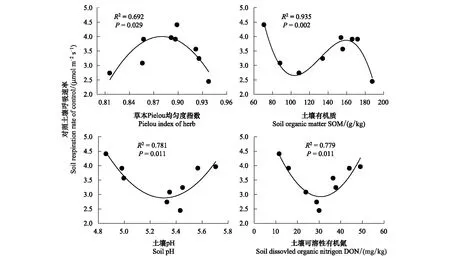
3种林型不同凋落物处理土壤呼吸速率与土壤温度呈极显著指数相关(P<0.01),土壤温度可解释土壤呼吸速率变化的31.80%—58.84%(图3),与土壤湿度相关性不显著(P>0.05)。针叶林、针阔混交林、阔叶林的CK土壤呼吸Q10值分别为2.01、3.29、3.06,RL的Q10值较CK分别降低0.38、0.35、0.23,DL较CK分别增加0.07、降低0.51、降低0.37,针叶林的Q10值表现为RL 图3 土壤呼吸速率与温度的关系 土壤呼吸与温湿度的双因素模型分析(表4)表明,土壤温湿度共同解释针叶林中CK、RL、DL土壤呼吸季节变化的27.94%、46.19%、55.32%;针阔混交林中CK、RL、DL土壤呼吸季节变化的66.93%、30.74%、50.84%;阔叶林中CK、RL、DL土壤呼吸季节变化的45.16%、38.51%、36.68%,其中针叶林的CK,针阔混交林的RL、DL,阔叶林的CK、RL、DL温湿度共同解释率要低于对应的单因素模型解释率(图3),说明研究区域生长季节降水量充沛条件下,土壤温度是影响其碳排放的关键因素。 表4 土壤温湿度与土壤呼吸速率回归模型 通过土壤理化性质及物种多样性指数与土壤呼吸速率的非线性回归分析(图4),发现天然次生林土壤呼吸速率与pH、SOM、DON及草本Pielou均匀度指数显著相关仅出现在CK处理,这可能是因为凋落物的去除和加倍在短期内并未引起土壤温湿度以及土壤底物的明显变化。SOM与CK土壤呼吸速率呈极显著的3次曲线相关(P<0.01),可解释土壤呼吸速率变异的93.48%,土壤SOM含量低于99.20 g/kg或高于171.47 g/kg时,土壤呼吸速率呈下降趋势,在99.20—171.47 g/kg则表现为上升趋势,说明土壤有机质含量过高或过低可能会限制土壤碳排放;pH、DON和草本Pielou均匀度指数与CK土壤呼吸速率呈显著的二次曲线相关(P<0.05),解释率分别为78.08%、77.94%、69.25%,pH值低于5.30、DON含量低于30.50 mg/kg、草本Pielou均匀度指数大于0.88时,土壤呼吸速率表现为下降趋势,反之呈上升趋势。 图4 对照土壤呼吸速率与土壤理化性质、物种多样性的关系 森林地表凋落物数量和质量引起土壤微环境变化而影响到土壤微生物种类和数量[33],导致土壤有机质分解速率快慢程度不一[34],进而影响到土壤碳排放[35-36]。有研究表明,去除凋落物一方面减少了土壤有机碳的输入,另一方面附着在凋落物表层的土壤微生物也随之被去除,造成土壤呼吸速率降低[37]。本研究发现,去除凋落物使3种次生林土壤呼吸速率降低了17.65%—21.01%(P<0.05),降幅高于该区域人工云杉林(14.21%)[9]和次生桦木林(17%)[30],但低于温带油松次生林(18.8%—47.0%)[38],高于法国温带山毛榉林(8.0%)[39],与美国温带老针叶林(19.0%)相近[7],主要是因为不同林分物种组成结构不同,凋落物数量和质量存在差异,其周转速率快慢各异所致[29,40]。本研究结果显示,凋落物贡献率表现为针叶林(17.65%)<阔叶林(19.83%)<针阔混交林(21.01%),与冯华敏等[41]的研究结果(针叶林20.6%<针阔混交林21.1%)相似;针叶林地表凋落物具有较高含量的木质素、酚类等物质使得分解速率较低[42],导致其土壤呼吸贡献率小于阔叶林,与邓琦等[43]对鼎湖山森林的研究结果(阔叶林27.82%<针阔混交林33.29%<马尾松林36.35%)和韩海燕等[44]对三峡库区森林的研究结果(阔叶林11%<针叶林24%=针阔混交林24%)并不相同,说明凋落物的贡献率具有明显的空间变异性,可能与不同的林分结构、气候状况和立地条件等有关。 新鲜凋落物输入一般会带来微生物可利用的有效碳源[45],凋落物的增加会刺激有机质分解,但土壤呼吸以及土壤微生物活性对其响应时限不同,造成土壤“激发效应”响应的持续时间不尽相同[46],进而对土壤呼吸影响程度亦不同[47]。本研究中,加倍凋落物使3种次生林土壤呼吸速率比对照增加了6.76%—8.16%,其增幅小于去除凋落物后土壤呼吸速率的降幅,凋落物的加倍处理并未引起土壤呼吸成倍增加,与已有研究结果[14,48]相似,究其原因:一方面凋落物的去除减少了土壤有效性养分供给而影响到微生物数量及其活性,引发土壤呼吸速率降低[37,49];另一方面凋落物层厚度增加产生对土壤CO2排放的屏蔽作用与凋落物自身分解释放CO2的多少存在一定平衡关系[18,50]。 土壤呼吸的Q10值常受到土壤生物、底物有效性、温湿度以及实验方法等差异的影响而表现出不确定性[51]。本研究中,3种林型不同处理的Q10值变幅为1.63—3.29,位于我国森林土壤呼吸Q10值范围(1.10—5.18)内[52],其中次生针叶林对照Q10值(2.01)低于该研究区域的粗枝云杉人工林(4.09)[9]和岷江冷杉原始林(4.71)[27];次生针阔混交林对照Q10值(3.29)低于粗枝云杉阔叶混交林(3.56),高于红桦-岷江冷杉天然次生林(3.06)[27]。不同处理下的Q10值均表现为针叶林<阔叶林<针阔混交林,其中针阔混交林和阔叶林Q10值均表现为对照高于去除和添加凋落物(表2),这与已有研究结果一致[53],表明凋落物输入方式的改变会降低土壤呼吸温度敏感性,其原因可能是凋落物的添加和去除缩小了地表日温差导致温度敏感性降低[54];但在次生针叶林中,加倍凋落物则提高了温度敏感性,这可能是因为土壤呼吸温度敏感性变化较为复杂,并非仅受单一因素制约,而是受土壤温湿度、物种特性、底物供应等多因素协同作用影响[1,27,35]。 不同林型土壤理化性质差异导致土壤微生物群落结构及活性、土壤酶活性、植物根系发育等不同,进而影响土壤呼吸速率的变化[55],而物种多样性的高低也是造成土壤呼吸出现差异的重要因素之一[56-57]。本研究发现,土壤pH、有机质、可溶性有机氮与草本Pielou均匀度指数是影响天然次生林对照土壤呼吸的主要因素。有机质是微生物的能量来源[58],微生物较高的底物亲和力与有机质的矿化密不可分,此外有机质也会对土壤通气性产生重要影响[59],从而间接影响土壤CO2排放;土壤pH可通过影响土壤基质、养分利用进而干扰微生物与酶的活性[60],使土壤呼吸发生变化。刘丽等[61]在对酸性土壤的研究中,发现pH值显著影响到土壤细菌、真菌群落结构与多样性,本研究中土壤pH变幅为5.18—5.37,亦有可能影响其土壤微生物群落,进而使土壤呼吸出现明显变化;土壤有机质与可溶性氮组分一般可通过干湿沉降、凋落物输入、根系和微生物代谢、根系与有机体的更替等途径进入土壤[62],可能会从多方面影响土壤呼吸作用。此外,土壤呼吸作为土壤理化性质的具体体现,也是植物生长繁育状况的重要表征,据Murphy等[57]的研究结果,树种构成复杂的森林较单种栽培其林木生物量更大,生长速度更快,他们认为更高的物种多样性有利于提高植物生产力[63],增加土壤碳输入,并通过影响根系呼吸或根际碳的供给水平影响微生物呼吸,进而提高土壤呼吸速率。 相关研究表明,土壤呼吸速率与土壤pH、有机质和可溶性有机氮显著线性正相关[27,64],且随植物群落的均匀度指数和丰富度指数的增大而增加[65]。本研究发现,天然次生林对照处理中,土壤呼吸速率与相关因子均呈显著的非线性相关,说明生境相似情况下,物种的群落分布及底物有效性是影响土壤呼吸的主要因素,但其对土壤呼吸的影响存在阈值,主要源于微生物与植物都有最适宜自身活动的群落结构和土壤养分含量,当土壤理化性质与植物多样性指数越接近适宜范围,土壤微生物与植物的活动就越旺盛,碳释放量就越高。此外,去除和添加凋落物后,天然次生林土壤呼吸与土壤理化性质、物种多样性指数均无显著相关性,可见土壤呼吸对土壤理化性质和植物多样性的响应较为敏感,凋落物输入方式对两者关联性的具体影响有待进一步的长期观测。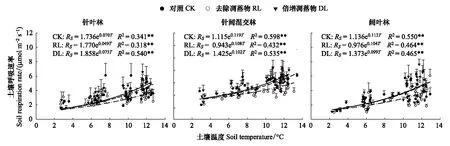
3.4 土壤呼吸与土壤理化性质、物种多样性的关系
4 讨论
4.1 凋落物去除和添加对土壤呼吸的影响
4.2 凋落物去除和添加对温度敏感性的影响
4.3 土壤理化性质和物种多样性对土壤呼吸的影响
猜你喜欢
现代农村科技(2022年5期)2022-11-18
农村农业农民·A版(2020年8期)2020-09-06
绿色中国(2019年18期)2020-01-04
现代农业研究(2018年4期)2018-08-11
中国绿色画报(2018年5期)2018-08-07
现代农业研究(2017年11期)2018-01-12
农家科技下旬刊(2017年8期)2017-11-13
中国农业文摘-农业工程(2017年2期)2017-01-12
科技创新导报(2016年21期)2016-12-17
农业与技术(2016年6期)2016-05-14
