
兰州市南山和北山 3种乔木叶片生态化学计量特征的对比研究
2021-04-25 08:24:40张小平罗子渝曹建军冯明铭赵慧君李空明李光栋
生态学报 2021年6期
魏 晨,张小平,*,罗子渝,曹建军,冯明铭,赵慧君,李空明,李光栋
1 西北师范大学地理与环境科学学院, 兰州 730070 2 甘肃省白龙江林业管理局林业科学研究所, 兰州 730070
甘肃省兰州市位于黄土高原最西端,以半干旱气候为主,生态脆弱,水土流失较为严重。为缓解水土流失,保障兰州市生态安全,20世纪初兰州市在南北两山进行了生态工程建设。侧柏(Platycladusorientalis)、圆柏(Sabinachinensis)和云杉(Piceaasperata)以其适应性强、抗严寒、耐干旱的特点成为我国西北黄土高原及中部干旱半干旱地区生态林建设的重要树种,在黄土高原开发和治理中有重要的作用[1]。有关这3种乔木的生态适应性与水分利用效率[2- 3]、固碳释氧量及净化空气[4- 6]、生理适应性及形态变化[7- 10]和育种[11- 12]等已有大量研究。但截至目前,对三者叶片生态化学计量的对比研究仍比较缺乏,以致无法全面了解其环境适应策略,并有效选择适合南北两山生境条件的人工林树种。
生态化学计量学(Ecological stoichiometry) 结合了生态学和化学计量学的基本原理,是研究生态系统能量和多重化学元素(主要是C、N、P)平衡的科学[13- 14]。自1958年Redfield[15]提出生态化学计量概念以来,有关生态化学计量特征的研究在国内外迅速开展,内容跨越生态系统各个层次(如种群、群落等)且尺度较大[16- 18]。叶片作为植物光合作用的主要器官,既是植物生长状况的“敏感器”,也是能量与物质的“交换所”[19]。叶片化学计量可构建一个集成养分框架(integrative nutrient framework),这一框架能将从细胞或有机体到全球尺度上的生物地球化学循环联系起来,从而反映植物养分利用效率和其所处特定生长条件等关联的适应性策略[20]。然而,由于跨区域或大尺度上研究通常无法有效解释微生境对叶片化学计量产生的影响[21],在区域中观尺度上开展植物叶片生态化学计量特征研究显得尤为重要。
以兰州市人工林(主要树种为圆柏、侧柏和云杉)为例,探讨3种乔木叶片化学计量特征之间的异同,以期揭示不同乔木的环境适应策略,从而为丰富区域性植物叶片化学计量研究,以及该市南北两山人工林的合理经营等提供决策依据。
1 材料与方法
1.1 研究区概况
兰州市南北两山是典型造林区,地处黄土高原西部的丘陵沟壑区,干旱草原地带的最北端,海拔1560—171 m[22]。属温带半干旱大陆性季风气候,昼夜温差大,年平均气温5—9℃,干燥少雨,年降水量250—360 mm, 且主要集中在6—9月[1]。土壤为黄土母质上发育起来的灰钙土,土层深厚,有机质含量低,保水保肥能力极差。植被类型属于荒漠与草原的过渡带,主要乔木有侧柏(Platycladusorientalis)、国槐(Sophorajaponica)、云杉(Piceaasperata)、圆柏(Sabinachinensis)、桃树(Amygdaluspersica)、银杏(GinkgobilobaL.)和榆树(UlmuspumilaL.)等,其中主要优势种为圆柏、侧柏和云杉。
1.2 实验设计与方法
2018年10月,于兰州市南北两山,对人工林中圆柏、侧柏及云杉的叶片及土壤进行采样。南山选取1个采样点 (半阳坡),地理坐标为(36.03°N, 103.81°E),海拔为1754.6 m;北山选取2个采样点 (半阳坡和半阴坡各一个),地理坐标分别为 (36.08°N,103.78°E)和(36.07°N,103.81°E),海拔分别为1674.1 m和1630.7 m。所选采样点的立地条件基本一致(均在兰州市地段内,坡度都为相对平缓的台地),包含3种优势种乔木(以配对取样),且林相较为整齐。
叶片采样:每个样点内选取3个样地(10 m×10 m),选取长势良好的个体,采集每棵植株向阳中上部位健康成熟的叶片,装入信封袋。经烘干和磨碎后,装入锡箔纸并标记。
土壤采样:每个样地内,沿对角线设置3个样方(两端和中点),用直径为35 mm的土钻分层(0—10、10—20、20—40 cm) 取样,样品装入自封袋,编号后,带回实验室处理。
1.3 样品测定
采用酸度计测定土壤pH,烘干法测定土壤含水率,重铬酸钾-硫酸外加热法测定叶片和土壤有机碳(SOC),凯氏法测定叶片和土壤全氮(STN),钼锑抗比色法测定叶片和土壤全磷(STP)[23]。土壤基本理化性质及其化学计量特征如表1所示。

表1 土壤基本理化性质及其化学计量特征
1.4 统计与分析
采用 SPSS 22.0 (SPSS Inc.Chicago, USA)统计软件进行数据分析。单因素方差分析用于比较不同土层土壤含水率、pH、SOC、STN、STP含量及其化学计量比和不同树种叶片C、N、P含量及其计量比的差异。Pearson相关分析用于分析叶片C、N、P含量及计量比的相关性。冗余分析用于探究影响叶片C、N、P含量及其化学计量比的主要土壤因子,以及它们各自与土壤因子的关系。变异系数为标准差/平均值×100%。数值用平均值±标准差表示,显著性水平为P<0.05。运用 CANOCO 4.5软件进行冗余分析及作图。
2 结果
2.1 3种乔木叶片C、N、P含量及其计量比的变异特征
云杉、圆柏叶片C含量 (497.64、496.02 g/kg)显著高于侧柏叶片 (484.12 g/kg) (表2);侧柏叶片N、P含量 (17.70、1.66 g/kg)显著高于云杉叶片 (15.62、1.40 g/kg)和圆柏叶片 (14.04、1.42 g/kg) (表2)。3种乔木叶片C的变异系数分别为2.07%、2.25%、2.26%,叶片N的分别为5.64%、16.05%、23.04%,叶片P的分别为14.79%、31.33%、35%。叶片C、N、P含量的变异系数大小均为圆柏<侧柏<云杉。
云杉、圆柏叶片C∶N (33.32、35.42)、C∶P (389.57、360.52)都显著高于侧柏叶片 (28.17、306.07);云杉叶片N∶P (11.88)显著高于圆柏叶片 (10.20)和侧柏叶片 (10.98),且后两者叶片N∶P无显著差异 (表2)。3种乔木叶片C∶N的变异系数分别为4.98%、17.68%、20.29%,叶片C∶P的分别为3.56%、3.65%、5.84%,叶片N∶P的变异系数分别为19.22%、26.96%、33.83%。叶片C∶N、C∶P、N∶P变异系数的大小均为圆柏<侧柏<云杉。
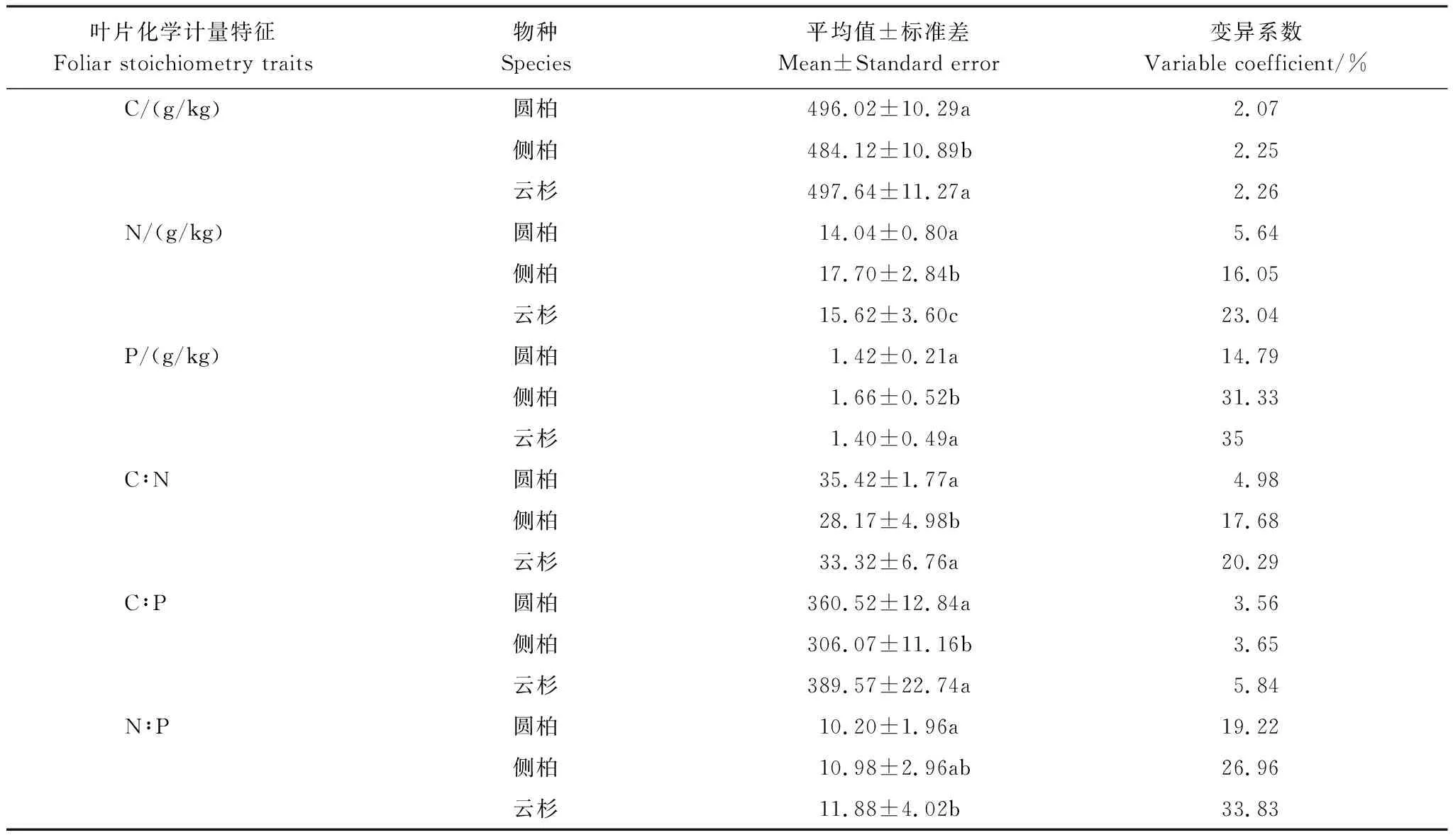
表2 圆柏、侧柏、云杉叶片化学计量特征及变异系数
2.2 3种乔木叶片化学计量的关系
圆柏叶片C含量与N含量正相关,侧柏叶片C含量与N含量负相关、与C∶N、C∶P正相关;侧柏、云杉叶片N含量与P含量正相关与C∶N、C∶P负相关,圆柏叶片N含量与C∶N负相关;3种乔木叶片P含量与C∶P、N∶P负相关(C∶P和N∶P正相关),侧柏和云杉叶片C∶N与C∶P正相关(表3)。
2.3影响3种乔木叶片化学计量的主要土壤因子
DCA分析结果表明,云杉、侧柏、圆柏叶片化学计量的最大梯度长度分别为0.328、0.306、0.140(均<3),故采用线性模型RDA进行排序分析。云杉叶片C、N、P化学计量第Ⅰ轴和第Ⅱ轴的解释量分别为32%和0.6%,前两轴的解释量之和为32.6%,对叶片C、N、P化学计量和影响因子关系的累计解释量高达98%;侧柏叶片C、N、P化学计量第Ⅰ轴和第Ⅱ轴的解释量分别为41.8%和0.3%,前两轴的解释量之和为42.1%,对叶片C、N、P化学计量和影响因子关系的累计解释量为99.1%;圆柏叶片C、N、P化学计量第Ⅰ轴和第Ⅱ轴的解释量分别为5.6%和3.9%,前两轴的解释量之和为9.9%,对叶片C、N、P化学计量和影响因子关系的累计解释量为56.3%(表4)。这说明3种乔木叶片C、N、P化学计量与土壤因子的关系可由前两轴解释,但均以第Ⅰ轴为主(表4)。

表3 3种乔木叶片元素含量及其化学计量比的相关系数

表4 基于RDA分析的叶片C、N、P化学计量与土壤因子的关系
3种乔木SOC∶STN的连线均最长,说明SOC∶STN对其叶片化学计量起主导作用。3种乔木叶片N都与SOC、SOC∶STN、SOC∶STP正相关,圆柏叶片C与SOC、SOC∶STN、pH正相关,云杉叶片N,侧柏、云杉叶片P和STP负相关但与SOC∶STP、STN∶STP正相关;3种乔木叶片C∶N与SOC∶STN、SOC∶STP负相关,侧柏叶片C∶P和云杉叶片C∶P、N∶P与STP正相关(如图1)。

图1 云杉、侧柏、圆柏叶片C、N、P 化学计量与土壤因子关系的冗余分析二维排序图Fig.1 Redundant analysis of the relationships between foliar C, N, P stoichiometry of three major trees and soil factors SOC∶土壤碳含量, Soil organic carbon;STN∶土壤氮含量, Soil total nitrogen;STP:土壤磷含量, Soil total phosphorus
3 讨论
3.1 3种乔木叶片C、N、P含量异同的原因
C、N、P是构成植物体最基本的化学元素,其中叶片C是组成植物体内干物质最主要的元素,N是蛋白质、核酸、叶绿素、脂质的组成部分,与光合作用紧密相关,而P对核糖体的产生至关重要,也是RNA、DNA、ATP的重要组成部分[20,24]。研究区内圆柏、侧柏和云杉叶片C含量均高于全球和全国陆生植物叶片C平均含量[13,17]、祁连山排露沟流域云杉叶片C含量[25]、华北石质山地侧柏人工林叶片的C含量[26]和云南普洱地区乔木叶片C含量[27](表5)。说明三者叶片的有机化合物含量较高,对外界不利环境的抵御能力较强[28],因为兰州市地处黄土高原西部,光照充足,光合速率较高且营养代谢旺盛,有利于有机物的有效积累[29]。云杉、圆柏叶片C含量显著高于侧柏叶片(表2),表明云杉、圆柏碳同化能力优于侧柏,因此在兰州南北山可适当增加云杉、圆柏的种植数量。这与该树种较强的水分获取和光合作用能力有关,与Yang等[30]的研究结果一致,但与郝鑫杰等[4](呼和浩特市人工林)和张亚民等[31](郑州市常见乔木树种)的研究结果不一致。其中,前者发现圆柏的碳同化能力优于云杉和侧柏,而后者发现圆柏的固碳释氧量较低。因此,生境差异是物种间碳同化能力不同的重要原因之一[32]。
3种乔木叶片N含量均低于全球、全国植物叶片N含量,这可能与土壤氮难以转化为植物可直接吸收利用的土壤有效氮有关[33],但具体机制还需进一步研究。三者叶片P含量低于全球植物叶片P含量[18](表5),与乔木叶片P含量较低的特性一致[34],因为全球植物叶片包含草本和灌木植物,而其叶片的N, P吸收效率通常较高[30,35]。侧柏叶片N、P含量分别显著高于云杉、圆柏叶片N、P含量(表2),说明侧柏的资源竞争力较强。这与马仁义等[36]对侧柏和祁连圆柏的研究结果相反,因此他们认为圆柏可替代侧柏作为绿化树种。造成这一结果差异的可能原因,与物种自身的遗传特性及不同区域养分利用策略的分异有关[37- 38]。另外,物种不同,地表凋落物、地下根系及其根系分泌物也不同,从而影响叶片N、P含量,但其机理有待进一步研究。
3种乔木叶片C、N、P变异系数均为圆柏<侧柏<云杉(表2),但C含量的变异最小(均小于3%),P含量的变异均大于N含量的变异,这与许雪赟等[39]对青藏高原火绒草及贺和亮等[40]对青藏高原窄叶鲜卑花的研究结果一致,说明草本、灌木和乔木叶片元素含量的变异基本一致。叶片C元素含量变异较小的原因在于:C主要通过光合作用转化,是构成植物骨架的基本结构物质,为植物生理活动提供能源,需求量较大,且受环境的影响较小[39-40];叶片P含量变异大于N含量变异的原因在于:与P含量相比,植物体内N含量的阈值范围一般较小[41]。

表5 叶片C、N、P含量及其化学计量比与其他研究地区对比
3.2 3种乔木叶片化学计量比异同的原因
植物叶片C∶N、C∶P代表植物吸收N、P元素时能同化C元素的能力,反映了植物的生长速率和养分利用效率[14]。生长速率假说认为叶片C∶N、C∶P越低,植物生长速率越快[42]。本研究中,圆柏、侧柏和云杉叶片C∶N和C∶P均值均高于全球尺度陆生植物叶片C∶N和C∶P[18](表5),说明它们叶片C元素同化能力较高,N、P元素吸收利用效率较低,且生长缓慢,这是植物适应养分贫瘠生境(表1)的一种选择性适应策略[43]。圆柏和云杉叶片N、P含量均低于侧柏叶片,导致其叶片C∶N和C∶P均高于侧柏叶片(表2)。该结果与张光德等[44]的研究结果不一致:他们发现圆柏叶片C∶N高于云杉,而云杉叶片C∶P高于圆柏。这可能因区域差异使物种对养分的选择不同而造成[45- 46]。圆柏与云杉叶片C∶N、C∶P无显著差异,与杨文高等[47]的研究结果一致,说明二者的生长速率差异较小,因为在相同的生境中它们采取了同一营养策略[27]。另外,这一无差异的结果也可能与植物化学计量的内稳态有关,即:在长期进化过程中,随着外界环境的变化,植物能保持其化学组成的相对恒定性[14]。
研究表明,叶片C∶N、C∶P能有效反映竞争和防御策略之间的平衡。N、P含量较高,而C∶N、C∶P较低时,植物采取竞争策略;反之,采取强有力的防御策略[48- 49]。本研究中,圆柏和云杉叶片C含量、C∶N、C∶P均高于侧柏(表2),说明在兰州市南北两山人工林系统中,为了维持自身的生长发育,云杉和圆柏采取了防御策略,侧柏采取了竞争倾向策略,以通过营养生态位的分化,实现物种的共存[50- 51]。植物叶片N∶P作为判断生境对植物所需养分的供应状况及植物生长速率的重要指标,决定植物在特定环境下的生长策略[38,52]。Koerselman[52]通过施肥实验对湿地生态系统进行了研究,发现植物叶片N∶P<14时,生长受N限制;叶片N∶P>16时,受P限制;14≤N∶P≤16时,为N、P共同限制。本研究中,云杉叶片N∶P显与侧柏叶片无差异(表2),在一定程度上反映了同一地区不同物种对养分利用及对外界环境适应与进化方面的趋同性[51]。三者叶片N∶P均小于14(表2),说明这3种树种生长都受N限制,与白雪娟等[53]、赵一娉等[54]对黄土丘陵区主要的人工林树种 (如刺槐、侧柏)的研究结果一致。云杉的叶片计量比的变异系数最小(表2),说明其养分含量较为稳定,抵抗外界干扰能力强[16],这可能与植物的生长发育和生理代谢有关。叶片为了维持正常的生理功能,其含量及计量比保持恒定,也是植物遗传或适应环境的结果[13]。
3.3 叶片化学计量自相关的原因分析
圆柏叶片C含量与N含量正相关(表3),与McGroddy等[55]的研究结果一致。因为叶片C主要来源于光合作用,而光合作用速率受叶片养分影响,通常高N含量会促进光合作用,进而有利于C的积累[49]。侧柏叶片C含量与N含量负相关(表3),符合高等陆生植物C、N等计量关系的普遍规律。这一规律把植物体中的C比作溶液中的溶剂,把N元素比作溶液的溶质,即C对N有“稀释作用”[14]。云杉叶片C含量与N含量不相关(表3),这可能与研究区水分缺乏有关(表1)。水分缺乏使 C 的同化与N的吸收过程不同步,即C和N的呈解耦趋势[56]。侧柏、云杉叶片N含量与P含量正相关,与黄小波等[27]的研究结果一致,这是因为叶片在建成过程中对土壤中N、P元素的吸收具有一定的比例关系,且通常为同向关系[27,57]。
3种乔木叶片N含量与C∶N、C∶P负相关,表明叶片C比叶片N、P更加稳定[58],而其叶片P与C∶N、C∶P、N∶P负相关,说明叶片在固C过程中对N、P利用效率的权衡策略不同[59]。三者叶片C∶N、C∶P、N∶P两两正相关,说明植物在吸收营养元素时具有一定的比例关系,也体现了叶片属性之间的经济策略[60]。
3.4 土壤因子对3种乔木叶片化学计量的影响
冗余分析表明,SOC∶STN主导3种乔木叶片的化学计量变化(图1),与杨文高等[47]的研究结果一致,说明研究区树木的生长主要受N调控。圆柏叶片C与SOC正相关的原因在于叶片将部分通过光合作用固定的C以枯落物的形式补偿给土壤[61],而与pH正相关的可能原因是:叶片在光合作用过程中所需水分较多,最终导致浅层土壤pH增大[62]。三者叶片N及侧柏叶片P与SOC正相关,主要是因为SOC来源于地表凋落物及根系凋亡分解的积累,而植物N、P含量在植物与土壤养分循环过程中较为稳定[63- 64]。云杉叶片N、P和侧柏叶片P与STP负相关,与章广琦等[65]的研究结果一致,表明随着植物从土壤中吸收N、P和叶片中单位质量N、P含量的增加,滞留在土壤中的STP含量会越来越少[39]。圆柏叶片P与STP不相关,可能因圆柏叶片受多种因素(生物、气候、土壤养分浓度)的影响,使STP对叶片P的影响被其他因子的作用抵消[66]。
3种乔木计量比与土壤因子的关系(图1)表明,植物体内各器官元素分配时相互协调,且N、P吸收效率与土壤养分紧密相关,因为植物通过凋落物和根系凋亡形成土壤有机质, 而土壤有机质分解矿化为植物正常代谢活动提供必要的营养元素[47,67]。另外,三者叶片计量比与土壤因子关系的不同,说明植物对土壤营养元素的吸收和利用是一个极其复杂的过程,即使在同一地区,不同物种(生物学特性不同)对同一种养分的需求及对环境的适应方式也不同[68- 69]。
4 结论
对兰州市南北两山3种乔木(云杉、圆柏、侧柏)叶片生态化学计量特征进行了分析,发现云杉叶片固碳能力及养分稳定性均强于圆柏和侧柏。因此,适当增加其种植数量,从而有利于碳的固持和平衡。
在养分贫瘠的环境中,3种乔木的竞争策略不同,其中云杉和圆柏采取了防御策略,而侧柏采取了竞争倾向策略。土壤营养元素对3种乔木叶片化学计量均有显著的影响,且其生长均受N限制。通过人工添加氮肥,可促进研究区乔木的生长。
叶片化学计量具有区域依赖性特点。本研究取样范围较小,研究结果是否适合于整个南北两山地区,还需进一步的论证。另外,研究区的人工林多为混交林,除了本研究中选取的树种外,其他树种的化学计量也有待研究,以最终找到一个适合大多数乔木树种的管理手段,如优先树种和施肥元素的选择等。
猜你喜欢
花卉(2024年1期)2024-01-16 11:33:50
林业科学研究(2023年4期)2023-04-29 00:44:03
新农民(2022年22期)2022-11-07 23:38:06
星星·诗歌原创(2021年3期)2021-06-01 09:13:23
现代农村科技(2020年7期)2020-12-19 03:12:08
新疆大学学报(自然科学版)(中英文)(2020年2期)2020-07-25 01:40:40
热带农业工程(2019年5期)2019-12-22 15:38:04
启迪与智慧·教育版(2018年12期)2018-02-26 12:24:06
山西林业科技(2017年4期)2018-01-23 05:45:05
河北林业科技(2016年5期)2016-11-08 03:12:49
