
半干旱区不同围封年限对草地土壤和微生物碳氮含量的影响
2021-04-16 07:44:18张强,邓军,毛瑾,朵莹,程杰,郭梁
水土保持通报 2021年1期
张 强, 邓 军, 毛 瑾, 朵 莹, 程 杰, 郭 梁
1.西北农林科技大学 草业与草原学院, 陕西 杨凌 712100; 2.宁夏云雾山国家级自然保护区管理局, 宁夏 固原 756000; 3.国家林业和草原局西北调查规划设计院, 陕西 西安 710048;4.西北农林科技大学 水土保持研究所, 陕西 杨凌 712100; 5.中国科学院 水利部 水土保持研究所, 陕西 杨凌 712100)
围栏封育作为一种有效且简便易行的促进退化草地自然恢复的方法,通过人为降低或完全排除家畜对草地生态系统影响,使草地生态系统在自身弹性下得以恢复和重建,在干旱半干旱区,特别是黄土高原植被恢复重建中广泛应用[1]。但有关退化草地适宜的围封面积和围封年限等关键问题却一直存在较大争议[2]。相比大量的以植被物种多样性和生产力等地上指标变化来确定最佳封育年限研究[3-5],植被恢复过程中土壤养分、土壤微生物量碳和氮含量变化趋势明显,其作为评价土壤肥力、土壤质量早期变化及植被恢复演替的有效性亟待加强[6]。
生态化学计量学通过分析多重元素(主要是C,N,P)质量平衡对生态交互作用的影响来探讨生态系统结构与功能[7]。土壤C∶N是表征土壤碳氮元素动态平衡的指标,其往往被用来检测土壤质量,能够调控土壤微生物对有机物的分解作用及碳的释放与吸收[8]。目前,生态化学计量学研究主要集中在植物叶片及土壤养分方面,而分析围封年限对根际土养分影响的报道较少。因此,本研究选取中国西部半干旱地区草原植被恢复的成功典范—宁夏云雾山国家级自然保护区的试验区,对不同封育年限样地中根际土壤和微生物量碳、氮含量及其化学计量特征进行研究,探讨其对封育年限的响应规律,以期为半干旱草地生态系统物质循环研究以及生态系统养分限制判定等提供依据,并为确定合理围封年限提供科学参考。
1 材料与方法
1.1 研究区概况
选取宁夏回族自治区固原市的云雾山国家级自然保护区试验区为研究点(106°21′—106°27′E,36°10′—36°17′N)。保护区自1982年以来一直定位监测退化草原恢复情况,至目前已近40 a,积累了丰富的草地群落变化资料。研究区地处黄土高原西南部,具有典型的半干旱气候特征,年平均降水量为425 mm,其中7—9月降水量占全年60%以上;年平均气温7 ℃,1月份温度最低,平均最低气温-8.2 ℃,8月平均最高气温25.2°。土壤主要类型为山地灰褐土。目前研究区物种数为313种,优势物种包括长芒草(Stipabungeana)、大针茅(Stipagrandis)、甘青针茅(Stipaprzewalskyi)、百里香(Thymusmongolicus)、白莲蒿(Artemisiasacrorum)以及星毛委陵菜(Potentillaacaulis)等。
1.2 试验设计
2017年8月底,对不同封育年限的草地生态系统进行了全面的群落调查及土壤样品采集,分别为放牧样地(GE0)、封育10 a样地(GE10)、封育25 a(GE25)和35 a(GE35)样地。其中,放牧样地为全年连续放牧(4只羊/hm2)。每个样地随机设置有代表性的试验小区3个,小区面积50 m×100 m,每个小区间隔80~100 m。在每个小区内随机选取10个采样点进行土壤样品采集,齐地去除植物地上部分后,用直径10 cm的根钻采集0—20 cm深度土样,将根系与土壤一齐取出,利用抖落法[9]采集根际土壤,用毛刷收集入无菌自封袋后将10个样点土样均匀混合为一个土壤样品。过2 mm筛后将每份土样分为两部分:一部分在4 ℃条件下保存,用于微生物量碳、微生物量氮含量测定,另一部分阴干后在实验室测定土壤养分有关指标。
1.3 测定项目与方法
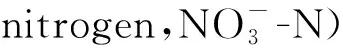
土壤微生物量碳(microbial biomass carbon,MBC)和微生物量氮(microbial biomass nitrogen,MBN)的测定采用氯仿熏蒸浸提法[13]。即称取25 g相当于烘干土重的新鲜土壤在25.8 ℃下用CHCl3熏蒸24 h,然后将100 ml 0.5 mol/L K2SO4加入土壤中,震荡1 h(200 r/min)后过滤去除熏蒸剂,同时另外提取25 g未熏蒸土壤。使用Elementar Liqui TOCII分析仪测定提取物的有机碳和氮含量。
1.4 数据统计分析
采用Excel 2016进行数据整理,共采集土样120个,表中数据为平均值±标准差。采用SPSS 25.0软件对不同围封年限的土壤数据进行单因素方差分析(one-way ANOVA)检验并运用LSD法进行多重比较,显著性水平为0.05。采用Pearson相关分析法对不同围封年限下土壤和微生物量C,N含量及其化学计量的相关性进行检验。
2 结果与分析
2.1 不同封育年限对土壤及微生物量碳、氮含量的影响

表1 云雾山自然保护区不同封育年限下土壤及微生物量碳、氮含量变化
2.2 土壤及微生物量碳、氮化学计量关系对封育年限的响应
土壤碳氮比(C∶N)为SOC与TN比值。由图1可知,研究区放牧和不同封育年限草地土壤C∶N的变化范围为7.72~9.16,平均值为8.31,变异系数为9%,不同封育年限土壤C∶N较为稳定,放牧样地最低,GE25样地最高,但未达显著差异水平。封育年限对微生物量C∶N则有显著影响,随封育年限延长,微生物量C∶N逐渐下降,GE35最低。qMBC为MBC与SOC的比值,qMBN为MBN与TN的比值,qMBC和qMBN常被用作评价土壤肥力和土壤质量变化的有效指标,本研究LSD分析表明封育年限对qMBC影响差异不显著;qMBN在GE10时最低,GE35最高,显著高于其他处理。化学计量不平衡性是指土壤碳氮比与微生物量碳氮比的比值,其分析结果表明,GE0,GE10和GE25之间差异均不显著,GE35最高,且与前3个阶段差异显著。硝态氮∶铵态氮的变化规律与微生物量C∶N较为相似,随封育年限增加,其值呈下降趋势。
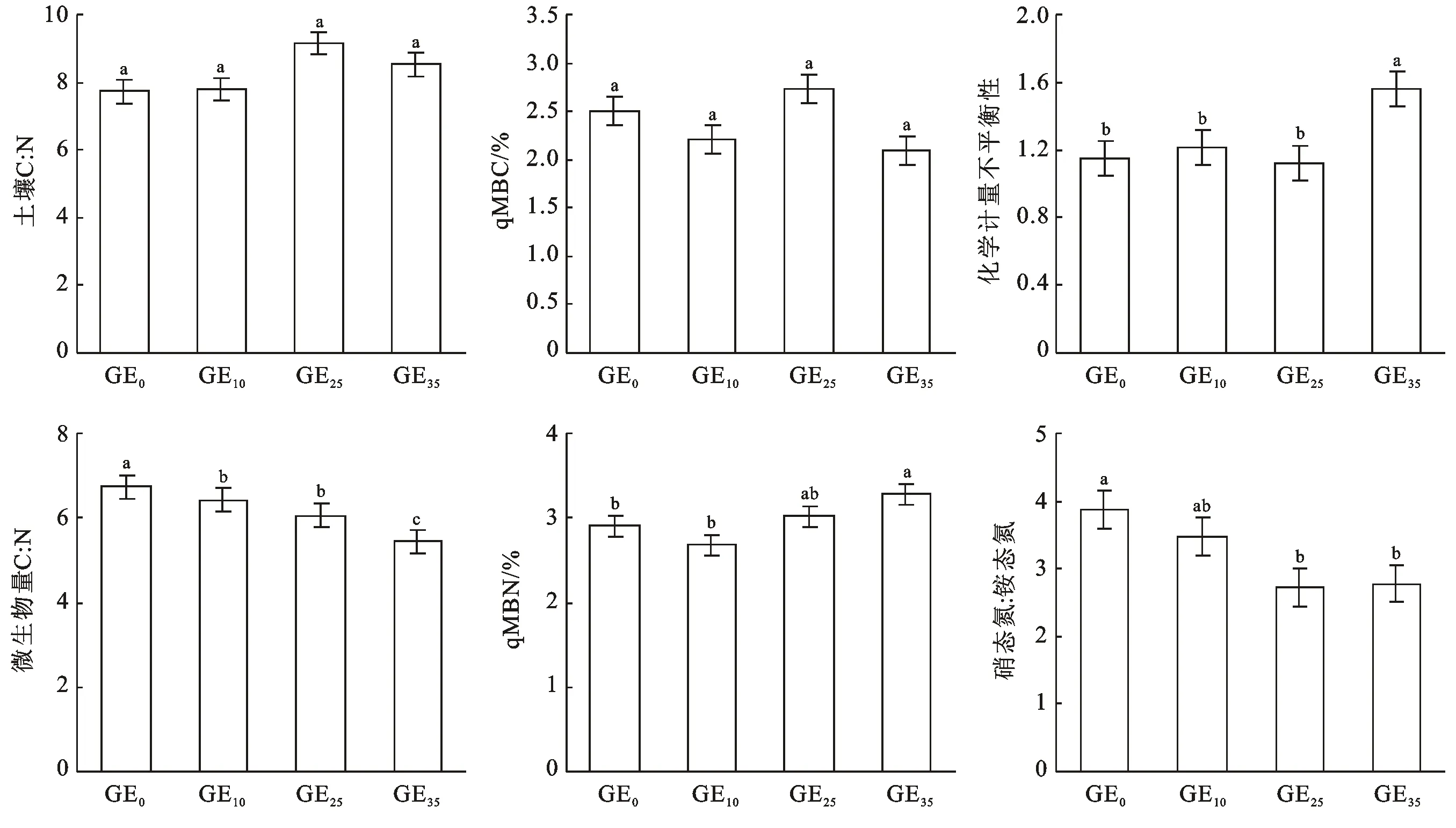
注:①图中不同小写字母表示差异显著; ②GE0,GE10,GE25,GE35分别表示放牧地,围封10 a,围封25 a,围封35 a。
2.3 土壤及微生物量碳氮和化学计量特征的耦合关系
相关性分析结果(表2)表明土壤有机碳与总氮和微生物量氮以及MBC∶MBN具有显著相关性(p<0.05),与土壤C∶N,微生物量碳和qMBC之间具有极显著相关性(p<0.01),特别是与qMBC的相关系数达到0.8,而与qMBN相关性较弱。土壤总氮与微生物量碳、微生物量氮和qMBC之间均存在极显著的线性相关,但土壤C∶N却与微生物量碳、氮和qMBC,qMBN以及MBC:MBN之间均无显著相关性。微生物量碳与微生物量氮之间具有极显著相关性,此外,qMBN与MBC:MBN之间也存在显著的相关关系。
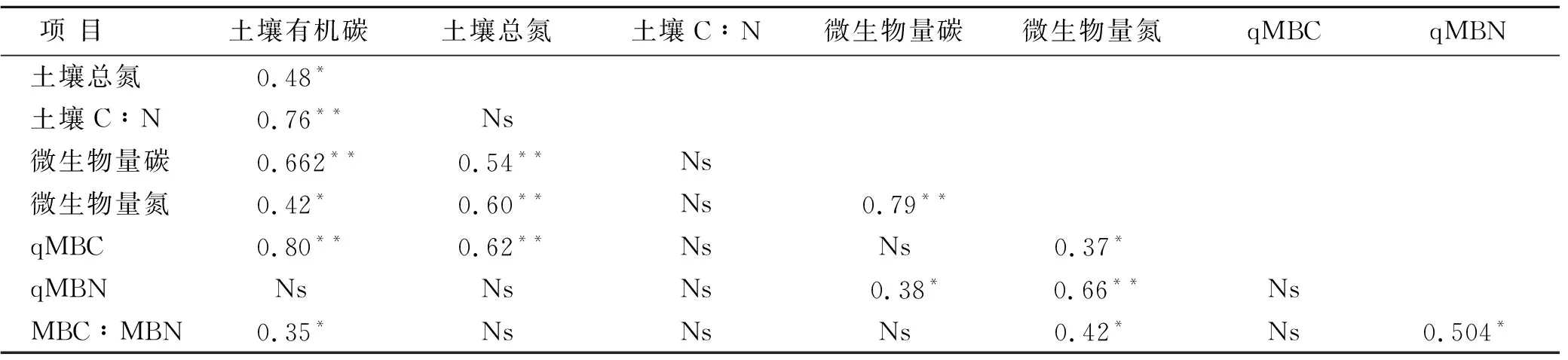
表2 土壤和微生物量C,N含量及化学计量的相关性
3 讨 论
3.1 土壤碳、氮及其化学计量关系随封育年限变化规律
研究表明,在植被演替过程中磷含量相对稳定[14],而土壤有机碳和氮含量作为除磷之外表征土壤质量状况的主要生态因子,其含量对地上植被生长发育影响较大,因此在封育植被演替过程中,探讨土壤碳、氮及其化学计量关系变化对于准确评价封育后植被恢复情况具有重要意义。本研究中,封育后土壤碳、氮含量较放牧地相比,均显著升高,这与前人研究类似[15-16]。土壤氮和有机碳主要来源于生物死亡后分解形成的稳定有机质[17],围封后家畜(主要是绵羊)采食植物被限制,从而极大增加了地上生物量的积累,进而使围封样地碳、氮含量较放牧地相比显著提高[18]。值得注意的是,尽管围封后土壤有机碳和总氮含量均高于放牧地,但二者值却在GE25达到峰值后急剧下降,这可能与演替进程中地上群落变化有关,之前研究表明物种多样性的峰值通常出现在演替中后期[19],Zhang等[20]研究发现半干旱草原封育25 a后地上物种多样性和地上生物量最高,随后下降,与本研究中土壤有机碳和总氮含量变化趋势一致。因此,演替后期地上生物量减少导致土壤有机质输入量降低是造成土壤有机碳和总氮含量在GE25后急剧下降的主要原因。土壤铵态氮和硝态氮可被植物较快吸收用于自身生长发育,所以本研究中围封地和放牧地相比,土壤铵态氮与硝态氮含量变化不大,甚至随着封育演替后期植物对养分需求的增大,硝态氮含量有所下降。
土壤生态化学计量作为表征土壤内部养分循环状况的重要指标,对于揭示元素的可获得性和平衡机制及生态恢复过程具有重要意义[21]。本研究中土壤C∶N随围封年限变化保持稳定,与周正虎和王传宽[17]整合分析全球生态系统演替过程中土壤碳、氮动态变化结果相一致,说明土壤碳、氮含量在生态系统演替过程中具有同步性,这也解释了土壤碳、氮在封育时间上具有显著的相关性。然而,也有一些研究发现土壤C∶N在演替中是可变的,例如张增可等[22]对海岛森林不同演替阶段土壤和植物碳、氮、磷化学计量特征的研究指出土壤C∶N随演替进行呈增加趋势。造成这种差异的原因可能是不同生态系统所提供的养分含量和质量不同,与林地地上部分和根系分解可提供给土壤丰富养分相比,地上生物量和根系相对较低的草地向土壤中添加的C和N较少。研究进一步指出,有机体往往具有较稳定的C∶N比例,即使在植物枯枝落叶进入土壤后也存在紧密的C-N耦合[23]。Zeng等[24]对云雾山草原保护区的研究表明,在30 a的封育禁牧期间,凋落物C∶N和根系C∶N没有明显变化。因此,稳定的C∶N可能与碳、氮含量紧密耦合的地上凋落物和植物根系相关。同时,稳定的C∶N比也表明,放牧禁牧对该半干旱退化草地土壤C,N平衡影响不大。
3.2 土壤微生物量及其化学计量关系随封育年限变化规律
土壤微生物是土壤有机质和养分循环转化的动力,是土壤养分转化过程中一个重要的活性库或源,能够有效促进土壤养分有效性和其在陆地生态系统的循环[25]。面对土壤碳、氮资源变化,微生物会通过调节自身生物量碳和氮以适应土壤资源的变异[26]。本研究显示,围封后微生物量碳、氮含量与土壤碳、氮含量的变化趋势一致,均呈现先增加至GE25达到最高后再下降趋势,但因其变化程度大小不同,引起微生物量C∶N随封育时间的延长而显著下降,这一点与土壤C∶N差异较大。本研究的相关性分析也显示,土壤C∶N与微生物量C∶N之间不存在显著相关关系。Xu等[27]对陆地生态系统研究也未发现微生物C∶N和土壤C∶N具有相关关系,支持了本研究的结论。微生物量C∶N产生这种变化的原因可能与微生物群落结构的改变有关,Mooshammer等[28]通过整合分析微生物对其资源的适应机制后提出微生物可以通过调整群落结构以非稳态的方式使其生物量组成适应其资源。Nicolas等[29]研究发现微生物量C∶N与真菌∶细菌的比值成正相关,表明随围封时间的延长,研究区真菌:细菌比值有所下降,亟待开展不同封育年限草地微生物群落特征研究。
3.3 化学计量不平衡性与微生物熵随封育年限变化规律
化学计量不平衡性值的高低可以表征资源质量水平和微生物的生长效率,化学计量不平衡性越高就表示资源质量越差,微生物的生长效率也就越低[28]。本研究发现,长期封育(GE35)能够显著影响化学计量不平衡性,相较放牧地和其他封育年限的化学计量不平衡性,其值在GE35显著增高,这是微生物调整其比例适应演替后期的结果,表明过长时间封育会降低资源质量水平及有效性。对于封育后的草地,其微生物的许多特性并不能完全由微生物量展现出来,因此,除分析其绝对量之外,微生物熵(即微生物量C,N分别与土壤C,N的比例)的变化能够较好地反映土壤中活性有机碳、氮所占的比例,从微生物学的角度揭示土壤肥力的差异。Zhou等[30]发现qMBC会随土壤C∶N增加而减少,而本研究却发现与放牧地相比,围封处理对qMBC和qMBN无显著影响,这可能是封育后草地受外界条件干扰较少,微环境相对稳定所致。
4 结 论
围封能够显著影响土壤有机碳和全氮的含量。随着封育年限的延长,其含量均呈现出先增加后下降的趋势,二者值都在GE25时达到峰值。但封育对C∶N影响却不大,草地在封育后的演替中,土壤有机碳和总氮之间有显著的相关性,二者具有同步性。
围封对土壤微生物量碳、氮及其化学计量特征也具有显著的影响,微生物量碳和氮对封育时间的响应与土壤有氮和有机碳对封育时间响应趋势相一致。封育后,微生物量C∶N显著下降至GE35最低。GE10,GE25与放牧地相比qMBN和化学计量不平衡性不显著,但GE35样地此值却显著增加,qMBN与微生物量C具有极显著的相关关系。qMBC对围封时间无显著响应。
适当的围封时间(GE25)对于草地恢复具有积极作用,但围封时间过久(GE35)就会对草地恢复产生负面影响,造成资源浪费,因此退化草地在适当围封后应该解封放牧或适当刈去地表植物进行结构调整。
猜你喜欢
天津农林科技(2022年2期)2022-04-19 10:48:14
河南畜牧兽医(2022年3期)2022-04-13 09:12:58
现代园艺(2021年23期)2021-12-01 07:47:44
现代畜牧科技(2021年4期)2021-07-21 06:13:08
绿色科技(2021年10期)2021-06-23 03:13:06
林业勘查设计(2020年1期)2021-01-18 02:40:48
新农业(2020年18期)2021-01-07 02:17:08
草业学报(2019年2期)2019-02-25 01:57:16
中成药(2017年6期)2017-06-13 07:30:35
动物营养学报(2015年3期)2016-01-07 11:06:10
