
农田景观和管理方式对稻田系统植物多样性的复合影响
2021-04-12 07:41:42杨梅刘念阎艳吴芸紫刘章勇
生态科学 2021年1期
杨梅, 刘念, 阎艳,吴芸紫, 刘章勇, *
农田景观和管理方式对稻田系统植物多样性的复合影响
杨梅1, 刘念1, 阎艳1,吴芸紫2, 刘章勇1, *
1. 长江大学农学院, 湿地生态与农业利用教育部工程研究中心, 荆州 434025 2. 湖北省荆州市农业科学研究院, 荆州 434000
农田生物多样性对于维持农田生态系统服务功能和农业可持续发展具有重要意义。为揭示农田景观特征和管理方式对农田生态系统植物群落和物种多样性的影响, 以有机稻田(ON)、常规化防稻田(天然林脚下, CN)、常规化防稻田(距天然林>10km, CF)系统为研究对象, 调查了田内、田埂和沟渠生境的植物物种组成和多样性特征, 并对三种类型稻田系统共125个样方进行DCA排序分析。结果表明: (1)有机稻田的物种丰富度显著高于两种常规化防稻田, 其中, ON植物由51科101属109种组成, CN植物由35科69属74种组成, CF植物由23科49属54种组成; (2)ON的Simpson指数和Shannon-Weiner多样性指数与CN均没有显著性差异, 但显著高于CF; (3)无论是有机稻田还是常规化防稻田, 物种丰富度、Simpson指数和Shannon-Weiner指数均以田埂最高, 沟渠次之, 田内最低; (4)总体而言, 相同生境下, ON各多样性指数(除外)均高于CN, 但无显著差异, 而与CF差异显著。此外, DCA排序图显示, 相同生境下, ON和CN的植物群落相似性较大, 而CN和CF的群落相似性小。上述结果共同表明, 农田植物物种多样性维持受农田管理方式和农田景观特征的复合影响, 低集约化农田景观对于维持农田生态系统植物物种多样性和群落组成具有更重要作用。
有机稻田; 常规化防稻田; 农田景观; 植物多样性; 生境
0 前言
作为生物多样性的重要组成部分, 农田生物多样性的保持对于养分积累与循环、土壤结构和肥力、空气过滤、传粉、增加天敌数量等均有重要作用[1]。在现代农业生态系统中, 农业集约化不仅简化了农业景观格局, 降低其空间异质性和时间异质性, 同时集约化的农事管理和大量农业化学品(如除草剂和杀虫剂)的投入, 造成生境质量急剧降低, 不利于农田生物多样性维持和农业可持续发展[2-4]。大量证据表明, 农业集约化是许多农田鸟类[5]、杂草[6]、传粉昆虫[7-8]、地表节肢动物[9]以及土壤动物和土壤微生物[10-11]等物种丰富度及多度下降的主要原因。如何减轻农业集约化对环境和生物多样性保护的负面影响, 已成为广泛受关注的问题[12-14]。作为重要的可持续发展的农业系统, 有机农业被视为实现农业景观生物多样性和生态系统服务功能的重要途径[15]。在过去近20年, 国外许多学者以现代常规化防农业生产系统作为对照, 开展了有机农业对农田生物多样性影响的研究[7-8,15-17]。国内在此领域的研究相对较少[18], 且主要集中在有机管理方式对农田动物的影响方面[19-22,], 但是, 对植物群落组成和植物物种多样性的影响的报道仍较少[23]。
与常规化防农业相比, 有机农业是一种在生产过程中不使用任何化学合成的农药、化肥、生长调节剂等, 也不采用基因工程获得的生物及其产物的农业生产体系[24]。作为对生物和环境友好的农田管理方式, 实施有机农业成为恢复农田生物多样性的一项重要措施。但是, 仅在局部尺度上改变农田管理方式以促进农田生物多样性的方法并未获得广泛支持, 原因可能是受到较大空间尺度上景观结构(landscape structure)特征的复杂影响[4, 25-28]。Brittain等[29]研究发现, 景观单一的有机农场中, 访花昆虫的多样性并没有明显提高; Rey Benayas 和 Bullock[30]的研究表明, 相较于异质性较高的农田景观, 有机农业对物种丰富度的积极影响更可能出现在高度集约化的农田景观中。低集约化的农田景观, 如农田周围的森林、灌木丛、池塘、杂草带等, 其多样化的生境为昆虫或其它动物提供更为多样的栖息环境和适宜的小气候环境, 对农田生物多样性维持可能具有重要作用[31-34], 且农田生态系统中植物物种组成和多样性将直接影响农田动物多样性(如农业害虫天敌和传粉动物)[35]和土壤微生物多样性[28]。尽管如此, 较大尺度的景观结构和局部尺度的农田管理方式对于植物多样性的复合影响的案例仍较少, 还有待进一步研究。
江西省宜春市万载县从1999年开始进行有机农业生产和有机产品的出口贸易, 2011年被确定为首批“国家有机产品认证示范创建区”, 同时也是全国唯一的以有机农业为产业特色的国家现代农业示范区。目前, 该区域有机农业规模达182.87 km2, 其中有机认证耕地面积为55.34 km2, 有机认证耕地中种植规模较大的作物为水稻39.95 km2, 生产的有机稻出口欧盟。有机稻田分布在由天然林包围的平洼地或梯田中, 具有复杂的农业景观; 同时, 该区域具有两种不同景观背景的常规化防稻田, 分别位于天然林脚下和距离天然林较远(>10 km)(图1)。这种距天然林“近”或“远”的分类方法可以作为农田景观背景的两种极端类型[7]。因此, 该区域为研究不同农田管理方式和农业景观对植物多样性的复合影响提供了理想的研究调查对象。研究拟调查并比较不同农田景观背景下, 常规化防稻田和有机稻田系统的植物群落分布格局、物种组成和物种多样性特征, 以评估农田管理方式和景观特征对农田植物群落的影响。该研究可为理解农田生物多样性维持机制和保护农田生物多样性提供科学依据。
1 材料和方法
1.1 研究区概况
研究区万载县位于江西省西北部(N27°59′37″— 28°27′48″, E113°59′13″—114°36′11″), 多山环抱, 丘陵广布。土壤类型以山地红壤为主, 土层深厚, PH值为4.5—5.0。该地区属亚热带湿润气候, 四季分明, 气候温和, 雨量充沛, 光热资源充足, 4—10月间为作物生长季, 平均日照时数在100 h以上, 7—8月为最多, 平均日照时数在200 h以上, 无霜期256 d, 全年平均气温16.9℃—18.2℃; 年平均降雨量为1742.5 mm, 但各地雨量分配不均, 呈北多南少的降水分布。降水的季节性差异较大, 由于季风影响, 4—6月降水最多, 占全年降水量的42%左右。
有机稻田(图1a, organic paddy near the natural habitats, ON)3个样地分别位于万载县仙源镇株木村和茭湖乡茭湖村、上峰村, 其有机稻种植历史16年。两种常规化防稻田(conventional paddy), 分别位于天然林附近(图1b, conventional paddy near the natural habitats, CN)和距离天然林较远(图1c, conventional paddy far from the natural habitats, CF)。其中, 近天然林常规化防稻田(CN)的3个样地位于三兴乡, 样地之间相距1—2 km, 常规化防种植历史6—8年; 远天然林常规化防稻田(CF)的3个样地均位于白良乡, 样地之间相距约2km, 常规化防种植连续8—10年。ON和CN周围的天然林以壳斗科青岗栎、钩栗、石栎和樟科香樟为主, 伴生有马尾松、枫香、毛竹等, 灌木层主要有檵木、白栎、杜鹃、盐肤木、乌饭、胡枝子和油茶等。上述三种类型稻田的种植模式主要为中稻—紫云英(冬闲田), 其中, CN和CF的田间病虫害根据当地情况及时预防各类病虫害的发生, 通过喷施除草剂去除杂草, 其它田间管理措施同当地大田生产, 且调查样地在前半个月内都未进行人工除草或喷洒除草剂。
1.2 调查方法
本研究选取ON、CN和CF三种类型稻田各3个样地。农田生态系统包括作物生境和其周围的半自然生境, 如田埂、沟渠等。因此, 每个样地调查生境包括田内(interior)、田埂(field bund)和沟渠(drainage ditch),每种生境类型随机选取5个具有代表性的样方进行调查, 其中沟渠的调查范围包括沟渠边坡和沟渠内。根据草本植物调查的样方标准进行调查[36], 设定样方面积为1 m2(1 m×1 m或0.5 m×2 m), 累计样方数量为125个(其中, ON和CF各45个样方, CN有35个样方)。各稻田系统植物群落调查时间为2016年5月, 该季节为草本植物生长的茂盛期, 同时是水稻生长苗期。调查并记录每个样方内的维管植物种类、盖度和频度, 对于在野外调查期间难以辨认的物种, 要求在原生境下拍照, 并制作标本, 带回室内以作进一步物种鉴定, 物种的科、属、种名称以《中国植物志》[37]为依据。
1.3 数据分析
物种多样性测度采用ɑ多样性的测度方法[38-39]: 物种丰富度指数()采用物种数(); 物种多样性分析采用Shannon-Weiner多样性指数(); 均匀度分析采用Pielou指数(); 优势度分析采用Simpson指数()。其中, 物种丰富度是一个群落中的物种数目, 它是基于物种的存在与否, 而不是物种的相对多度的一个多样性测度。相关参数计算公式如下:
物种丰富度指数():=

重要值():= ( 种i相对频度+种i相对盖度) / 2
式中: S为每一样地中的植物物种总数目, N为所有物种总个体数, Pi为第i种的相对重要值; 相对频度= (某物种频度/总物种频度之和) × 100%; 相对盖度= (某物种盖度/总物种盖度之和) × 100%。将各稻田类型的3个样地中的所有样方数据作为基础数据, 由上述公式分别计算ON、CN和CF的多样性指数; 将各稻田类型的3个样地中不同生境的各样方数据作为基础数据, 由上述公式分别计算ON、CN和CF各生境的多样性指数。

注: a.天然林包围的有机稻田(ON); b.天然林脚下的常规化防稻田(CN); c.距天然林较远(>10km)的常规化防稻田(CF)。
Figure 1The landscape of three types of paddy fields
群落排序分析是研究植物群落生态关系的重要方法, 可以揭示不同类型植物群落结构差异及其与环境的关系。只使用物种组成数据时采用非限制性排序方法, 如, 去趋势对应分析(Detrended Correspondence Analysis, DCA)应用较广泛[40]。构建群落-物种矩阵, 对9类样地、125个样方的植物群落进行排序分析, 物种在每个样方中的有或无以0—1表示。稀有种的存在会影响排序结果, 使最终解释与实际有所偏差[41]。因此, 排序之前进行了剔除稀有种(仅在一个样方中出现过的物种)的处理。
利用 Excel 2013 软件进行基础数据处理并计算各样地植物群落的物种多样性; 运用单因素方差分析和LSD对不同类型稻田的多样性各指标进行显著性检验和多重比较, 用< 0.05或者<0.01表示; 并采用Canoco 5.0软件对125个样方进行群落排序。
2 结果与分析
2.1 植物物种组成、物种丰富度和生活型分析
有机稻田(ON)的3个样地共有109种植物, 分属于51科101属, 种类较多的有菊科(13属14种) 和禾本科(12属12种), 共占物种总数的23.9%, 其次是莎草科(4属7种)、大戟科(4属4种)、蓼科(4属4种)和唇形科(4属4种)。近天然林常规化防稻田(CN)的3个样地共有74种植物, 分属于35科69属, 其中种类较多的有禾本科(14属14种)、菊科(9属9种)和莎草科(3属6种)。远天然林常规化防稻田(CF)的3个样地共有54种植物, 分属于23科49属, 其中种类较多的有禾本科(14属15种)、菊科(4属5种)和莎草科(2属4种)。与ON和CN相比, CF的物种数量明显下降(=0.0001), 存在于ON和CN的28种物种在CF中都未出现, 如蒲儿根()、断节莎()、糯米团()、慈姑()、半边莲()、垂盆草()、地耳草()、粟米草()、槐叶萍()等。
三种稻田系统不同生境的植物物种丰富度和种类差异较大。ON、CN和CF中均以田埂植物物种数为最多, 分别为70、57和40; 其次为沟渠, 物种数分别为63、33和32; 田内物种数最少, 分别为30、18和13。ON田埂植物总计70种, 三个样地平均值为36.0; 沟渠植物总计63种, 各样地平均值为35.0; 田内植物总计30种, 各样地平均值为16.3。CN田埂植物总计57种, 三个样地平均值为34.0; 沟渠植物总计33种, 各样地平均值为31.0, 田内植物总计18种, 各样地平均值为13.0。CF田内植物有13种, 各样地有植物8—10种, 平均值为9.0; 田埂共有40种植物, 各样地有植物18—27种, 平均值为23.0; 沟渠共有32种植物, 平均值为16.6(表1)。此外, 对ON、CN和CF相同生境下的物种数进行比较, ON的田内、田埂、沟渠的物种数量都极显著高于CN(=0.0003;=0.0001;=0.0001)和CF(=0.0001), 且CN各生境的物种数也显著高于CF(=0.0001;=0.0001;=0.0498)(图2)。
从生活型角度, ON的一、二年生植物共有53种, 占物种总数的48.6%, 多年生植物共有56种, 占物种总数的51.4%; CN的一、二年生植物共有43种, 占物种总数的58.1%; 而CF的一、二年生植物共有35种, 占物种总数的64.8%。与两种常规化防稻田相比, ON的多年生植物物种数量显著增多(= 0.0001)(图2)。
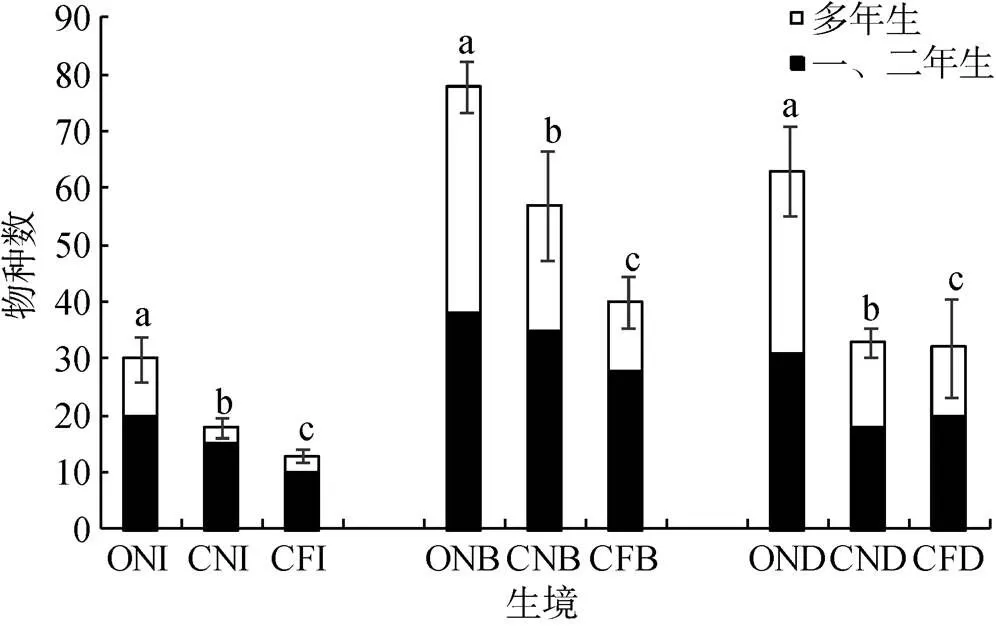
注: ONI、ONB、OND分别代表有机稻田(ON)田内、田埂、沟渠, CNI、CNB、CND分别代表近天然林常规化防稻田(CN)的田内、田埂、沟渠, CFI、CFB、CFD分别代表远天然林常规化防稻田(CF)的田内、田埂、沟渠。不同小写字母表示同种生境下不同稻田植物物种数差异显著(P<0.05)。
Figure 2 Number of plant species under different habitats in organic paddy (ON), conventional paddy near the natural habitats (CN) and conventional paddy far from the natural habitats (CF)
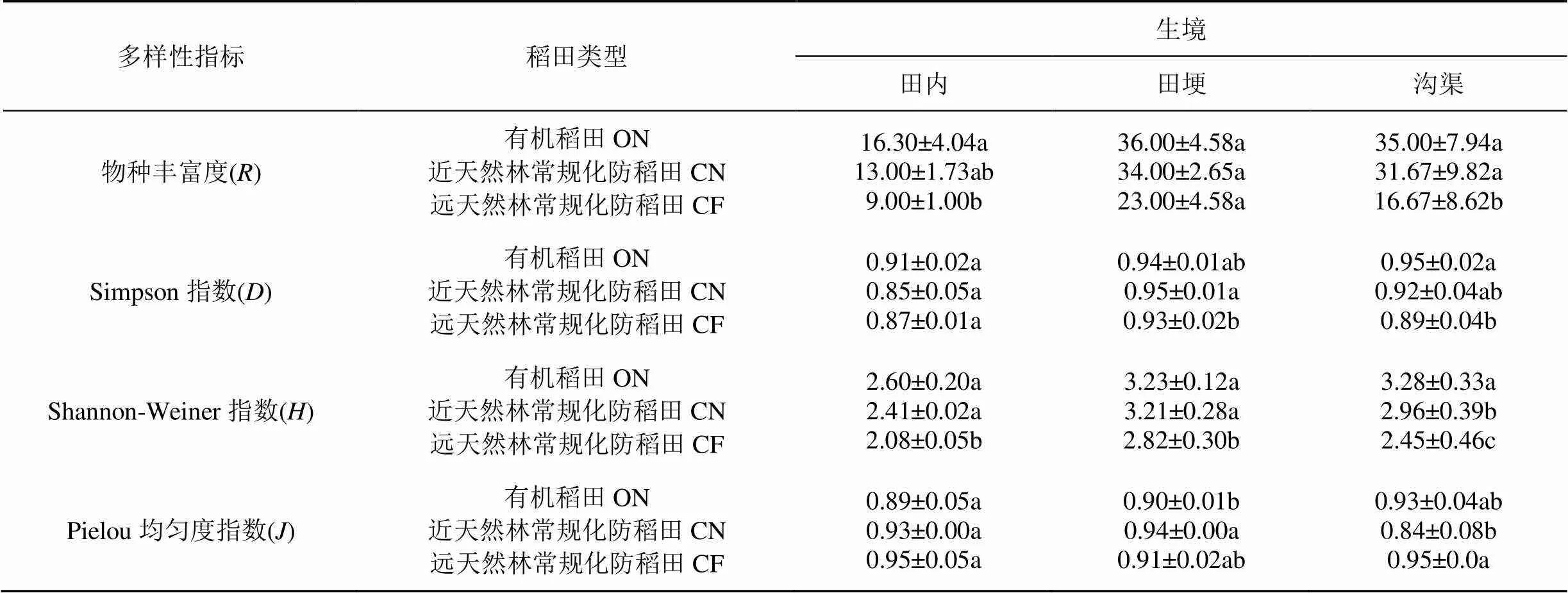
表1 有机稻田和常规化防稻田系统不同生境的物种丰富度及物种多样性比较(平均值±标准误)
同一多样性指数同列不同小写字母表示同一生境下不同类型稻田间差异显著(<0.05)。
2.2 稻田系统及不同生境间物种多样性比较
将有机稻田(ON)、近天然林常规化防稻田(CN)和远天然林常规化防稻田(CF)系统的各多样性指数进行比较(图3)。ON和CN的Simpson指数(0.96 vs. 0.96)、Shannon-Weiner指数(3.69 vs. 3.51)均没有显著性差异(=0.8982;=0.3416), Pielou均匀度()指数具有极显著性差异(0.88 vs. 0.93)(=0.0071); ON和CF的Simpson指数(0.96 vs. 0.93)和Shannon- Weiner指数(3.69 vs. 3.02)均有极显著性差异(= 0.0142;=0.009), Pielou均匀度指数(0.89 vs. 0.88)没有显著性差异(=0.0052); 而两种常规化防稻田CN和CF的Simpson指数(0.96 vs. 0.93)、Shannon- Weiner指数(3.69 vs. 3.02)均有显著性差异(= 0.0159,= 0.0326), Pielou均匀度指数(0.93 vs. 0.88) 具有极显著性差异(=0.0043)。

注: D、H、J分别代表Simpson优势度指数、Shannon-Weiner指数和Pielou均匀度指数; 不同小写字母表示不同稻田系统同一种多样性指数差异在0.05水平上显著。
Figure 3 Comparison of plant species diversity of ON、CN、CF
各稻田系统不同生境之间多样性指数差异较大。在常规化防稻田CN和CF中, 物种丰富度、Simpson指数和Shannon-Weiner指数均以田埂最高, 沟渠次之, 田内最低; 而在ON中, 田埂和沟渠的物种丰富度、Simpson指数、Shannon-Weiner指数和Pielou均匀度指数均高于田内, 且其中田埂和沟渠的物种丰富度(=0.0059;=0.0075)和Shannon- Weiner指数(=0.0178;=0.0209)均显著高于田内(表1)。
此外, 对各稻田系统相同生境的多样性指数进行比较(表1)。远天然林常规化防稻田(CF)的物种丰富度、Simpson指数和Shannon-Weiner指数均显著低于同生境下的有机稻田(ON), 而Pielou均匀度指数高于ON, 但无显著差异(田内:=0.1276; 田埂:=0.1871; 沟渠:=0.6464)。CN和ON相比, 物种丰富度、Simpson指数(田埂除外)和Shannon-Weiner指数均低于同生境下的ON, 但无显著差异; 相反, CN的Pielou均匀度指数均高于同生境下的ON(沟渠除外)。CN和CF相比, CN的物种丰富度、Simpson指数(田内除外)和Shannon-Weiner指数均显著高于同生境下的CF; 而CF的Pielou均匀度指数均高于同生境下的CN(除田埂外), CN和CF的沟渠之间(=0.0496)差异显著, 田内(=0.9654)和田埂(= 0.0771)无显著差异(表1)。
2.3 不同生境稻田植物群落的排序分析
对ON、CN和CF三种稻田系统不同生境共125个样方的植物群落进行去趋势对应分析(DCA排序)结果见图4。样方DCA图第1轴基本反映了生境的变化对群落物种组成的影响, 从左到右, 依次为田内、沟渠和田埂, 第2轴基本反映了群落沿稻田类型的变化梯度。DCA图中各个样点间距离表示样方内群落相似程度。距离越近, 则说明物种组成越相似; 若出现重叠, 则说明物种组成极为相似。第1排序轴和第2排序轴分别解释了53.49%和38.63%的差异, 说明第1轴在排序轴中占主导地位。图4显示, CFI、CFD、CFB处于排序图中间位置, 并出现重叠; CNI、ONI和CFI位于排序图左下方, 且CNI和ONI重叠区域较大, 两类样地相似性强; 相反, CNI和CFI、CNB和CFB不重叠或重叠区域小, 分别显示出较小的群落相似性。除CFD外, CN和ON的非作物生境(田埂和沟渠)样地, 包括CNB、ONB、CND和OND主要分布于排序图右方, 其中CNB和ONB, 以及ONB和OND重叠大, 分别显示较大的群落相似性。但是, 相较于ON, CF和CN相同生境的样地亦有重叠, 如CFB和CNB, 而CFI和CNI、CFD和CND分开, 表明农田景观特征促进二者产生分化。相反, CNI和ONI重叠, 说明相较于农田管理方式, 景观特征在农田植物群落结构中具有更重要的作用。
3 讨论
植物群落调查强度、有机管理时间和农田景观背景都可能对植物群落物种组成和多样性特征的结果产生影响。部分研究有一年两次、分别在夏秋进行植物调查, 以获取研究区域的植物名录全貌[42]。本研究在5月份进行的一次植物调查虽然不能反映整个年度植物多样性的状况, 但也是相对重要时期的反映。农田野生植物主要为草本植物, 春末夏初为草本植物生长的茂盛期, 类似的进行一次植物调查来衡量生物多样性在其它研究中广泛采用[14,23,36,43-44]。例如, Hyvönen[45]对芬兰南部的一个农场进行了连续6年的杂草群落研究, 以探讨氮肥施用和除草剂对植物物种多样性的影响, 其调查时间为每年8月; Pallett等[6]研究英国南部的一个农场由常规化防管理转为有机管理对农田植物物种丰富度的影响时, 进行了连续18年、每年一次的植物群落调查。此外, 本研究中, 无论有机稻田还是两种常规化防稻田, 其管理方式都相应的连续了多年(6—16年), 因此调查结果能无偏的反映农田管理方式和景观背景对农田植物多样性特征的影响。

ONI、ONB、OND分别代表有机稻田(ON)田内、田埂、沟渠, CNI、CNB、CND分别代表近天然林常规化防稻田(CN)的田内、田埂、沟渠, CFI、CFB、CFD分别代表远天然林常规化防稻田(CF)的田内、田埂、沟渠。
Figure 4 Two-dimensional DCA ordination diagram of 125 quadrats of plant community
农田管理方式能直接影响农田环境, 改变土壤营养的数量和质量, 并进一步使不同种类植物的生长发育受到不同程度的影响, 表现出植物群落物种组成和多样性特征差异。本研究发现, 有机稻田ON的物种丰富度要显著高于两种常规化防稻田CN和CF(109 vs.74; 109 vs.54), 植物物种数分别提高了47.3%和100.2%。与常规化防稻田相比, 有机稻田中, 与水稻伴生的田间杂草, 如千金子()、稗()等禾本科植物和异型莎草()、水蜈蚣()等莎草科植物数量大幅度减少, 而其它植物如鸭舌草()、慈姑、槐叶萍等数量剧增。此外, 有机稻田系统出现大量野生植物, 如蒲儿根、断节莎、糯米团、野荸荠()、白毛夏枯草)、紫堇()、地耳草、半夏()、合掌消()、油茶()等, 并促进稻田生态系统逐步形成草-灌复合群落。该结果表明不使用除草剂和化肥可以提高稻田植物物种数, 这可能是杂草长期适应土壤条件及与作物竞争的结果[46-47]。Hyvönen[45]的研究也表明, 连续6年的有机管理方式下, 农田植物物种平均数目提高了50%左右, 且除草剂对物种多样性和物种组成的影响远大于化肥的使用。Callauch[48]研究了相邻的有机农田与常规化防农田的杂草群落, 发现有机农田的杂草种类是常规化防农田的 2—3倍, 许多研究得到相似的结论[15]。Pallett等[6]对常规化防农田转为有机管理后的农田植物群落进行了连续18年的调查, 发现有机管理实施5年后, 植物物种丰富度达到稳定的水平, 为之前常规化防农田的3倍。而Heinke[49]的研究发现, 与常规化防农田相比, 有机农田的植物种类甚至高达10倍。同时, 植物生活型谱也发生了变化, 在常规化防稻田CN和CF中, 与水稻伴生的主要是一、二年生短生活周期植物, 如稗、千金子、鹅观草()和满江红(), 而在有机稻田中, 多年生植物增加, 这可能是因为化肥、农药和除草剂等化学用品停止施用和人工干扰变弱后, 一年生杂草植物得到了控制。同时, 与常规化防农田相比, 有机农田更能支持稀有物种[50]。例如, ON中出现了萝藦()、络石()、半夏、合掌消、地耳草等。Kay等[51]对濒危杂草的植物群落调查发现, 21种稀有物种中, 有机农田有11种, 常规化防农田只有3种, 8种在两类农田中都有, 但在有机农田更常见。
尽管如此, 也有研究表明有机农田杂草丰富度与常规化防农田没有差异[31], 甚至低于常规化防农田, 可能是受较大空间尺度上景观特征的影响[52], 但对于农药和化肥使用较少的农田, 有机农田的杂草种类要高于常规化防农田[45]。本研究中, 有机稻田ON和常规化防稻田CN的Simpson指数(=0.8982)和Shannon-Weiner指数(=0.3416)均没有显著性差异, 而与常规化防稻田CF具有极显著性差异(= 0.0142,=0.009), 且有机稻田ON和常规化防稻田CN中存在的28个物种在常规化防稻田CF中不存在。DCA排序图显示常规化防稻田CN和有机稻田相同生境的群落组成相似(CNI和ONI, CNB和ONB, CND和OND); 相反, 常规化防稻田CN和CF相同生境的群落组成差异更大(CNI和CFI, CNB和CFB, CND和CFD)。该结果表明距离天然林100m以内的有机稻田和常规化防稻田CN均受天然林的影响, 具有更复杂的景观特征, 表现出较高的物种多样性和物种丰富度, 而距离天然林较远的常规化防稻田CF没有或较少天然林的影响, 植物物种种类更少和多样性更低。
此外, 无论是常规化防稻田还是有机稻田, 物种丰富度和多样性的差异主要存在于田内、田埂和沟渠不同生境之间, 相对于作物生境, 半自然生境的田埂和沟渠的物种丰富度和多样性更高。有机稻田田内、田埂、沟渠的物种数分别为30、78和63, 显著高于两种常规化防稻田CN(18、57和33)和CF(13、40和32), 表明有机农田管理方式对于稻田植物物种丰富度和物种多样性有积极作用。同时, 农田景观结构中的非作物生境在生物多样性维持上具有重要意义[53]。
农田生物多样性受农田景观背景、空间尺度和农田管理方式等的影响[25, 54-55]。本研究通过比较不同景观特征下的有机稻田和常规化防稻田系统植物群落组成、物种丰富度和多样性特征差异, 评估了大尺度上的景观特征和小尺度上的农田管理方式对农田植物多样性维持的相对重要性。研究发现, 有机稻田的物种丰富度显著高于常规化防稻田; 且相同的复杂景观特征下, 有机稻田系统的多样性指数与天然林脚下常规化防稻田系统没有显著性差异, 但是, 均显著高于景观单一的集约化稻田。上述结果共同表明农田植物群落结构和物种多样性维持受农田生产管理方式和农田景观特征的复合影响, 低集约化农田景观对于维持常规化防农田的生物多样性有重要作用。
[1] MACE G M, NORRIS K, FITTER A H. Biodiversity and ecosystem services: a multilayered relationship[J]. Trends in Ecology and Evolution, 2012, 27(1): 19–26.
[2] 尹飞, 毛任钊, 傅伯杰, 等. 农田生态系统服务功能及其形成机制[J]. 应用生态学报, 2006, 17(5): 929–934.
[3] BENTON T G, VICKERY J A, WILSON J D. Farmland biodiversity: is habitat heterogeneity the key?[J].Trends in Ecology and Evolution, 2003, 18(4): 182–188.
[4] FISCHER C,THIES C, TSCHARNTKE T. Mixed effects of landscape complexity and farming practice on weed seed removal[J]. Perspectives in Plant Ecology, Evolution and Systematics, 2011, 13(4): 297–303.
[5] DONALD P F, GREEN R E, HEATH M F. Agricultural intensification and the collapse of Europe’s farmland bird populations[J]. Proceedings Biological Sciences, 2001, 168(1462): 25–29.
[6] PALLETT D W, PESCOTT O L, SCHäFER S M. Changes in plant species richness and productivity in response to decreased nitrogen inputs in grassland in southern England[J]. Ecological Indicators, 2016, 68: 73–81.
[7] KREMEN C, WILLIAMS N M, THROP R W. Crop pollination from native bees at risk from agricultural intensification[J]. Proceedings of the National Academy of Sciences of the United States of America, 2002, 99: 16812–16816.
[8] GODED S, EKROOS J, AZCáRATE J G, et al. Effects of organic farming on plant and butterfly functional diversity in mosaic landscapes[J]. Agriculture, Ecosystems & Environment, 2019, 284: 106600.
[9] HOLLAND J M, LUFF M L. The effects of agricultural practices on Carabidae in temperate agro-ecosystems[J]. Integrated Pest Management Reviews, 2000, 5(2): 109–129.
[10] HUANG P M, WANG M K, CHIUC C Y. Soil mineral–organic matter–microbe interactions: impacts on biogeochemical processes and biodiversity in soils[J]. Pedobiologia, 2005, 49(6): 609–635.
[11] CLIFTON E H, JARONSKI S T, HODGSON E W, et al. Abundance of soil-borne Entomopathogenic fungi in organic and conventional fields in the midwestern USA with an emphasis on the effect of herbicides and fungicides on fungal persistence[J]. PLoS ONE, 2015, 10(7): e0133613.
[12] BAUDRON F, GILLER K E. Agriculture and nature: Trouble and strife? [J]. Biological Conservation, 2014, 170: 232–245.
[13] SCHNEIDER M K, LüSCHER G, JEANNERET P, et al. Gains to species diversity in organically farmed fields are not propagated at the farm level[J]. Nature Communications, 2014, 5: 4151.
[14] SYBERTZ J, MATTHIES S, SCHAARSCHMIDT F, et al. Assessing the value of field margins for butterflies and plants: how to document and enhance biodiversity at the farm scale. Agriculture, Ecosystems & Environment, 2017, 249: 165–176.
[15] ELSEN T V, STOBBELAAR D J, MANSVELT J D V. Species diversity as a task for organic agriculture in Europe[J]. Agriculture, Ecosystems and Environment, 2000, 77: 101–109.
[16] SMUKLER S M, SANCHEZ-MORENO S, FONTE S J, et al. Biodiversity and multiple ecosystem functions in an organic farmscape[J]. Agriculture, Ecosystems and Environment, 2010, 139: 80–97.
[17] BENGUTSSON J, AHNSTROM J, WEIBULL A C. The effects of organic agriculture on biodiversity and abundance a meta–analysis[J]. Journal of Applied Ecology, 2005, 42(2): 261–269.
[18] 王长永, 王光, 万树文, 等. 有机农业与常规农业对农田生物多样性影响的比较研究进展[J]. 生态与农村环境学报, 2007, 23(1): 75–80.
[19] 钟平生, 梁广文, 曾玲. 有机稻田主要天敌类群及其群落多样性演替[J]. 中国生物防治, 2005, 21(3): 155–158.
[20] 杨国庆. 三种稻田节肢动物群落比较和水对稻田植株捕食功能的调控研究[D]. 扬州: 扬州大学, 2003.
[21] 李钰飞. 有机、无公害和常规蔬菜种植模式下温室土壤生物群落结构及食物网的特征研究[D]. 北京: 中国农业大学, 2014.
[22] 王磊, 李刚, 席运官, 等. 有机种植方式对稻田动物多样性的影响:以句容戴庄为例[J]. 生态与农村环境学报, 2018, 34(07): 614–621.
[23] 孙玉芳, 陈宝雄, 金彬, 等. 有机管理对不同农田生境草本植物α、β和γ多样性的影响[J]. 中国生态农业学报, 2019, 27(11): 1617–1625.
[24] GOMIERO T, PIMENTEL D, PAOLETTI M G. Environmental impact of different agricultural management practices: Conventional vs. organic agriculture[J]. Critical Reviews in Plant Sciences, 2011, 30(1/2): 95–124.
[25] 刘云慧, 李良涛, 宇振荣. 农业生物多样性保护的景观规划途径[J]. 应用生态学报, 2008, 19(11): 2538–2543.
[26] BATáRY P, MATTHIESEN T, TSCHARNTKE T. Landscape-moderated importance of hedges in conserving farmland bird diversity of organic vs. Conventional croplands and grasslands[J]. Biological Conservation, 2010, 143: 2020–2027.
[27] GABRIEL D,SAIT S M, HODGSON J A, et al. Scale matters: the impact of organic farming on biodiversity at different spatial scales[J]. Ecology Letters, 2010, 13(7): 858–869.
[28] TUCK SL,WINQVIST C, MOTA F, et al. Land–use intensity and the effects of organic farming on biodiversity: A hierarchical meta–analysis[J]. Journal of Applied Ecology, 2014, 51(3): 746–755.
[29] BRITTAIN C, BOMMARCO R, VIGHI M, et al. Organic farming in isolated landscapes does not benefit flower– visiting insects and pollination[J]. Biological Conservation, 2010, 143(8): 1860–1867.
[30] REY BENAYAS J M, BULLOCK J M. Restoration of biodiversity and ecosystem services on agricultural land[J]. Ecosystems, 2012, 15(6): 883–899.
[31] WEIBULL A C, OSTMAN O, GRANQVIST A. Species richness in agroecosystems: the effect of landscape, habitat and farm management[J]. Biodiversity and Conservation, 2003, 12 (7), 1335–1355.
[32] BIANCHI F J J A, BOOIJ C J H, TSCHARNTKE T. Sustainable pest regulation in agricultural landscape: a review on landscape composition, biodiversity and natural pest control[J]. Proceedings of the Royal Society B-Biological Sciences, 2006, 273(1595): 1715–1727.
[33] 谢坚, 屠乃美, 唐建军, 等. 农田边界与生物多样性研究进展[J]. 中国生态农业学报, 2008, 16(2): 506–510.
[34] ANDERSSON G K S, BIRKHOFER K, RUNDLÖF M, et al. Landscape heterogeneity and farming practice alter the species composition and taxonomic breadth of pollinator communities[J]. Basic and Applied Ecology, 2013, 14(7): 540–546.
[35] 齐月, 李俊生, 闫冰, 等. 化学除草剂对农田生态系统野生植物多样性的影响[J]. 生物多样性, 2016, 24(2): 228–236.
[36] 孟雪松, 欧阳志云, 崔国发, 等. 北京城市生态系统植物种类构成及其分布特征[J]. 生态学报, 2004, 24(10): 2200–2206.
[37] 中国植物志编撰委员会. 中国植物志[M]. 北京: 科学出版社, 1979.
[38] 马克平, 刘玉明. 生物群落多样性的测度方法Ⅰ: α多样性的测度方法(下)[J]. 生物多样性, 1994, 2(4): 231–239.
[39] 王寿兵. 对传统生物多样性指数的质疑[J]. 复旦大学学报: 自然科学版, 2003, 42(6): 867–868.
[40] 赖江山. 生态学多元数据排序分析软件Canoco 5介绍[J]. 生物多样性, 2013: 21(6): 765–768.
[41] 曹静, 苗艳明, 冯飞, 等. 稀有种不同处理对典范对应分析排序结果影响的比较[J]. 植物生态学报, 2015, 39(02): 167–175.
[42] 卢训令, 刘俊玲, 汤茜, 等. 黄淮海平原农业景观非农生境植物功能群特征分析[J]. 生态学报, 2019, 39(18): 6721–6730.
[43] PELLISSIER L, WISZ M S, STRANDBERG B, et al. Herbicide and fertilizers promote analogous phylogenetic responses but opposite functional responses in plant communities[J]. Environmental Research Letters, 2014, 9: 024016.
[44] 宋香静, 李胜男, 韦玮, 等. 山东省渤海沿岸滨海湿地草本植物群落物种组成及多样性研究[J]. 生态科学, 2017, 36(4): 30–37+63.
[45] HYVÖNEN T, SALONEN J. Weed species diversity and community composition in cropping practices at two intensity levels – a six–year experiment[J]. Plant Ecology, 2002, 154: 73–81.
[46] 李儒海, 强胜, 邱多生, 等. 长期不同施肥方式对稻油两熟制油菜田杂草群落多样性的影响[J]. 生物多样性, 2008, 16 (2): 118–125.
[47] 施林林, 沈明星, 蒋敏, 等. 长期不同施肥方式对稻麦轮作田杂草群落的影响[J]. 中国农业科学, 2013, 46(2): 310–316.
[48] CALLAUCH R. Ackerunkraut–Gesellschaften auf biologisch und konventionell bewirtschafteten Äckern in der weiteren Umgebung von Göttingen[J]. Tuexenia,1981,1: 25–37 (in German).
[49] HEINKEN T. Die Ackerwildkraut-Vegetation auf biologisch und konventionell bewirtschafteten Ackerflächen bei Gut Adolphshof (Ldkrs. Hannover)[J]. Beiträge zur Naturkunde Niedersachsens, 1990, 43: 38–45 (in German).
[50] ALBRECHT H, MATTHEIS A. The effects of organic and integrated farming on rare arable weeds on the Forschungsverbund Agrarökosysteme München (FAM) research station in southern Bavaria[J]. Biological Conservation, 1998, 86(3): 347–356.
[51] KAY S, GREGORY S. Rare Arable Flora Survey 1999[R]. Abingdon, UK: Report to Northmoor Trust and English Nature, 1999.
[52] LIEBMAN M, DAVIS A S. Integration of soil, crop and weed management in low–external–input farming systems[J]. Weed Research, 2000, 40(1): 27–47.
[53] BUREL F, LAVIGNE C, MARSHALL E J P, et al. Landscape ecology and biodiversity in agricultural landscapes[J]. Agriculture Ecosystems & Environment, 2013, 166: 1–2.
[54]刘云慧, 常虹, 宇振荣. 农业景观生物多样性保护一般原则探讨[J]. 生态与农村环境学报, 2010, 26(6): 622–627.
[55] 刘云慧, 张鑫, 张旭珠, 等. 生态农业景观与生物多样性保护及生态服务维持[J]. 中国生态农业学报, 2012, 20(7): 819–824.
Mixed effects of farm landscapes and management practices on plant diversity in paddy systems
YANG Mei1, LIU Nian1, YAN Yan1, WU Yunzi2, LIU Zhangyong1, *
1. College of Agriculture, Yangtze University, Engineering Research Center of Ecology and Agricultural Use of Wetland (Ministry of Education), Jingzhou 434025, China 2. Hubei Academy of Agricultural Sciences, Jingzhou 434000, China
Farmland biodiversity plays an important role in farm ecosystem services and sustainable agricultural development.To explore the mixed effects of farm landscape and management practices on plant community and species diversity in farm ecosystem, species composition and diversity were investigated in the interior, field bund and drainage ditch of the organic paddies (ON), conventional paddies (from the natural forest <100m, CN) and conventional paddies (from the natural forest>1km, CF). Detrended Correspondence Analysis (DCA) was carried out for 125 plots of three types of paddy field systems. The results showed the following: (1) there were 66 species (101 genera belonging to 51 families) distributed in ON, while 74 species(69 genera belonging to 35 families) distributed in CNand 54 species(49 genera belonging to 23 families)distributed inCF. (2) The Simpson dominance index and Shannon-Weiner diversity index ofONwere higher than that in CN but with no significant difference, while significantly higher than CF. (3) No matter the organic paddies or the conventional paddies, the diversity indexin field bund was higher than that in interior and drainage ditch, which showed the importance of non-crop habitats for maintaining biodiversity. (4) DCA ordination map showed that under the same habitat, plant communities of ON and CN had greater similarity, while those of CN and CF had smaller similarity. These results reveal that the plant diversity in paddy field is affected by mixed effects of management practices and landscape structure, and the low intensive farm landscapes may be the main driver of the diversity and composition of plant communities.
organic paddies; conventional paddies; farmland landscape; plant diversity; habitat
杨梅, 刘念, 阎艳, 等. 农田景观和管理方式对稻田系统植物多样性的复合影响[J]. 生态科学, 2021, 40(1): 43–51.
YANG Mei, LIU Nian, YAN Yan, et al. Mixed effects of farm landscapes and management practices on plant diversity in paddy systems[J]. Ecological Science, 2021, 40(1): 43–51.
10.14108/j.cnki.1008-8873.2021.01.007
Q14
A
1008-8873(2021)01-043-09
2019-10-17;
2019-12-27
湖北省重点研发计划项目(2020BBA044); 长江大学湿地生态与农业利用教育部工程研究中心开放基金(KF201804)
杨梅(1981—), 女, 湖北荆州人, 博士, 讲师, 主要从事保护生物学研究, E-mail: myang@yangtzeu.edu.cn
刘章勇(1965—) 男, 湖北荆州人, 博士, 教授, 主要从事农业生态学研究, E-mail: lzy1331@hotmail.com
猜你喜欢
幼儿100(2024年18期)2024-05-29 07:35:28
魅力中国(2021年50期)2021-11-30 03:16:04
山西林业(2021年2期)2021-07-21 07:29:28
农村百事通(2020年21期)2020-11-23 02:04:24
创新作文(小学版)(2019年22期)2019-12-07 05:44:44
绿色中国(2019年19期)2019-11-26 07:13:16
绿色中国(2019年19期)2019-11-26 07:13:16
新农业(2019年24期)2019-01-06 07:14:32
水利技术监督(2017年6期)2017-12-19 13:28:16
环境保护与循环经济(2017年10期)2017-03-16 03:16:20
