
自然生草对渭北旱塬苹果园小气候及果实灼伤和早期落叶病的影响
2021-04-08 10:55白岗栓郭江平杜建会
草地学报 2021年2期
白岗栓, 郭江平, 杜建会
(1.西北农林科技大学水土保持研究所, 陕西 杨凌 712100; 2.中国科学院水利部水土保持研究所, 陕西 杨凌 712100;3.博尔塔拉蒙古自治州林业和草原局, 新疆 博尔塔拉833400; 4. 江南大学, 江苏 无锡 214122)
在欧美及日本等国,果园生草是一种常见的土壤管理方式[1-3]。果园生草能够有效增加果园的生物多样性、增强果园土壤的蓄水保肥能力、改善果园的生态环境、提高果实品质及产量[4-8]。果园生草可分为人工生草及自然生草,人工生草需要对不同草种、不同地域的生态环境进行深入研究后才可选择适宜的草种进行播种,且需掌握相关的种植与管理技术,需投入一定量的劳动力;自然生草经过多年的自然淘汰与选择,往往春季萌发早、秋季枯黄迟、耗水量少、营养均衡,具有丰富的物种多样性,且不需要播种,只需定期刈割,就能有效改善果园的生态环境、提高果实品质、促进果园生态系统良性发展,目前已成为国内果园地面管理发展的新方向[9-13]。果园小气候是在一定的大气候背景下,由果树的树种、树龄、树冠结构与形态、郁闭度以及管理技术等综合影响下所形成的小气候环境。果园小气候影响果树与其所在环境之间的物质和能量交换,影响果树生长及其果实品质[14-16]。渭北旱塬是中国最大的优质苹果(Malusdomestica)生产基地,渭北旱塬西部的长武塬区,随着果园杂草的自然演替与选择,繁缕(Stellariamedia)和牛繁缕(Malachiumaquaticum)已成为果园地被植物的优势种类及顶级群落,不但覆盖度高、根系分布浅,而且不需要刈割,不与果树争水争肥,在渭北旱塬西部一带已快速推广[17]。果园自然生草在我国已有40年左右[18],近年来有关自然生草对果园土壤酶及土壤微生物[19-21]、土壤理化性状[22-24]、果实产量与品质[25-28]等报道较多,有关自然生草对梨(Pyrusbretschneideri)园、樱桃(Cerasusavium)园小气候环境的影响也有报道[29-30],但有关自然生草对渭北旱塬苹果园小气候的影响则未见报道。为了探寻自然生草对渭北旱塬苹果园小气候的影响,本试验以苹果园清耕和人工生草为对照,监测自然生草果树行间的温湿度、树冠温湿度、土壤温度、光照强度以及果实日灼和早期落叶病等,以期为自然生草调控果园的生态环境提供技术支撑。
1 材料和方法
1.1 试验区概况
试验于2018-2019年在中国科学院长武黄土高原农业生态试验站进行。长武黄土高原农业生态试验站位于渭北旱塬西部的长武塬区,海拔1 200 m,北纬35°12′,东经107°40′,年降雨量为551.7 mm,4~10月为494.1 mm,年日照时数2 226.5 h,年均气温9.1℃,≥10℃积温3 029℃,无霜期171 d。试验园土壤为黑塿土,耕层(0~20 cm)土壤有机质 9.44 g·kg-1,全氮 0.84 g·kg-1,全磷 0.24 g·kg-1,全钾 7.46 g·kg-1,碱解氮 73.65 mg·kg-1,速效磷 41.98 mg·kg-1,速效钾 234.65 mg·kg-1,pH值7.8,土壤容重 1.32 g·cm-3,田间持水量 22.21%,萎蔫系数 9.2%。试验前果园地面管理为自然生草,为繁缕和牛繁缕群落。试验园无灌溉设施,为雨养果园。
1.2 试验材料
供试苹果园建于1996年春季,面积为1.8 hm2,主栽品种为惠民短枝富士(M.domestica‘Fuji apple spur Huimin’),授粉品种为皇家嘎啦(M.domestica‘Royal gala’),砧木均为新疆野苹果(M.sieversii),株行距3.0 m×4.0 m,南北行向,小冠疏层形,干径10.0 cm左右,树高4.3 m左右,冠径3.8 m左右,其中惠民短枝富士外围延长枝长16.0 cm左右,皇家嘎啦为20 cm左右。试验前5年的单株产量平均为50.0 kg左右,处于盛果期。
自然生草的繁缕和牛繁缕均为石竹科的一年生或二年生草本,平伏于地表生长。人工生草草种为多年生的豆科牧草白三叶(Trifoliumrepens)。
1.3 试验设计与测定
1.3.1试验设计 试验以果园清耕和人工生草为对照,监测自然生草对果园小气候及果实灼伤等的影响。试验共设3个处理,每个处理重复3次,共9个小区。每个小区东西长40 m,南北宽30 m,面积1 200 m2,有9行9列81株苹果树。
试验于2018年4月至2019年10月进行。2018年长武塬区苹果遭遇严重晚霜危害,基本绝收。2018年4月初,用小型旋耕机对全园进行旋耕(深度15 cm),以消除长期自然生草对试验结果的影响。
自然生草:2018年4月初用小型旋耕机对全园进行旋耕(深度15 cm),让其自然生草,自然生草期间出现的大型恶性杂草如反枝苋(Amaranthusretroflexus)、灰藜(Chenopodiumalbum)等及时去除,保留繁缕、牛繁缕、鸡肠草(Herbacentipeda)、荠菜(Capsellabursa-pastoris)、蒲公英(Taraxacummongolicum)、箭叶旋花(Convolvulusarvensis)、马唐(Digitariasanguinalis)等低矮草本。2018年4月至2019年10月,由于自然生草以繁缕和牛繁缕为主,平伏于地表生长,草高未超过30 cm,故未进行刈割。
人工生草:2018年4月初果园用小型旋耕机旋耕后,在果树行间种植宽2.6 m的白三叶草草带(播种量15 kg·hm-2,距树行0.70 m)。2018年4月至2019年10月,树盘下的杂草及时去除。当白三叶草的高度达到40 cm时进行刈割并平铺于行间,刈割留茬高度15 cm左右,树盘下保持清耕。2018年分别于7月25日和10月2日进行刈割,2019年分别于6月25日、8月10日和10月4日进行刈割。
清耕:2018年4月初用小型旋耕机对全园进行旋耕后至2019年10月,果树行间及树盘下的杂草(包括繁缕和牛繁缕)均及时去除,保持果园无杂草滋生。
试验期间不同处理的修剪、施肥、病虫防治、疏花蔬果、果实套袋等管理措施相同。
1.3.2试验监测 果树行间空气相对湿度和温度:在不同小区中部的树行中间距地面150 cm处,悬挂DHM2型通风干湿温度表(天津气象海洋仪器厂),从2019年4月1日至2019年10月31日,每天 8∶00,10∶00,12∶00,14∶00,16∶00,18∶00和 20∶00监测行间空气相对湿度和空气温度月变化,并在夏至前后选择晴天连续3天从8∶00-20∶00每1 h监测1次行间空气相对湿度和空气温度的日变化。
树冠空气相对湿度和空气温度:在不同小区中部选择1树冠完整的惠民短枝富士,在树冠中部靠近树干东南部距地面1.8 m处,悬挂DHM2型通风干湿温度表,从2019年4月1日至2019年10月31日,每天8∶00,10∶00,12∶00,14∶00,16∶00,18∶00和 20∶00监测树冠空气相对湿度和空气温度月变化,并在夏至前后选择晴天连续3天从8∶00-20∶00每1 h监测1次树冠内空气相对湿度和空气温度的日变化。
土壤温度:在不同小区中部的树行中间,2019年4月1日至2019年10月31日,用L99-TWS温湿度记录仪(杭州路格科技有限公司)观测每天8∶00,10∶00,12∶00,14∶00,16∶00,18∶00和20∶00时的地表温度和5 cm,10 cm,15 cm及20 cm土层的土壤温度。
果园光照强度:2019年夏至前后选择晴天无风的天气,在不同小区中部的树行中间各选一个点距地面1.5 m处,用3台JD-3型照度计(上海嘉定学联仪表厂生产)同时测定不同处理果树行间中部的光照强度(6:00-20:00每1 h测定测定1次)[31]。
树冠反射光谱:2019年夏至前后选择晴天无风的天气,连续3天10:00-14:00,太阳高度角大于45°时,用Field Spec 3便携式地物光谱仪(美国ASD公司生产,测定光谱范围为300~2 500 nm)测定不同处理苹果树冠的反射光谱。
枝条尖削度:2019年落叶期用游标卡尺测定不同处理皇家嘎啦和惠民短枝富士树冠外围延长枝长度及直径,并计算尖削度。枝条尖削度=枝条粗度/枝条长度。
果实灼伤率:2019年8月20日左右皇家嘎啦成熟期,10月10日左右惠民短枝富士成熟期,常规方法调查不同处理皇家嘎啦和惠民短枝富士果实的灼伤率。
早期落叶病危害状况:皇家嘎啦果实成熟期和惠民短枝富士果实成熟期,常规方法调查不同处理皇家嘎啦和惠民短枝富士苹果树早期落叶病(褐斑病Cercosporainsulana,Cerosporaipomoeoeae和Cercosporaroseleri;灰斑病Phyllostictapirina和Coryneumfoliicolum;轮斑病Alternariamali)的发病率,并将早期落叶病划分为6级,根据不同级别的发病叶数和病级,计算发病指数。0级:叶面无病斑;1级,病斑所占叶片面积低于5%;2级:病斑所占叶片面积大于等于5%且低于15%;3级:病斑所占叶片面积大于等于15%且低于30%;4级:病斑所占叶片面积大于等于30%且低于45%;5级:病斑所占叶片面积大于等于45%且低于70%;6级:病斑所占叶片面积大于等于70%至落叶。

1.4 数据处理
试验数据用Excel 2010制作图表,用SPSS 19.0软件进行单因素方差分析;若差异显著,则采用邓肯氏新复极差检验法进行多重比较,检验不同处理间的差异显著性。
2 结果与分析
2.1 不同处理果园的空气相对湿度
4月至10月自然生草、人工生草和清耕果树行间空气相对湿度的月均值分别为50.42%,49.86%和45.11%,自然生草和人工生草的较清耕分别高出11.77%和10.53%,均极显著高于清耕(P<0.01)。自然生草的月均值略高于人工生草,但在6月至8月则略低于人工生草(图1A)。
4月至10月自然生草、人工生草和清耕果树树冠内空气相对湿度的月均值较其行间空气相对湿度月均值分别提高了9.98%,10.37%和9.75%,均显著高于其对应行间的空气相对湿度(P<0.05)。自然生草和人工生草果树树冠内空气相对湿度的月均值分别较清耕高出12.01%和11.16%,均极显著高于清耕(P<0.01,图1B)。

图1 不同处理行间及树冠内空气相对湿度月变化
夏季自然生草、人工生草和清耕果树行间空气相对湿度的日均值分别为50.00%,50.31%和42.50%,自然生草和人工生草的基本相同,均极显著高于清耕(P<0.01,图2A)。
夏季自然生草、人工生草和清耕果树树冠内空气相对湿度的日均值分别为53.97%,50.61%和46.65%,均显著高于其对应行间的空气相对湿度日均值(P<0.05)。自然生草和人工生草树冠内空气相对湿度的日均值均极显著高于清耕(P<0.01),自然生草和人工生草之间无显著差异(图2B)。
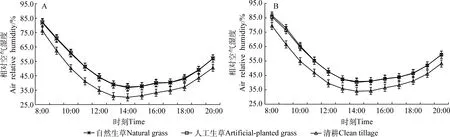
图2 不同处理行间及树冠内空气相对湿度日变化
2.2 不同处理果园的空气温度
4月至10月自然生草、人工生草和清耕果树行间空气温度的月均值分别为19.49℃,19.51℃和19.91℃,自然生草与人工生草的基本相同;清耕的略高于自然生草与人工生草,其中清耕5月至8月的月均值较自然生草和人工生草的均高出0.57℃,但未达到显著差异(图3A)。
4月至10月自然生草、人工生草和清耕果树树冠内空气温度的月均值分别为19.00℃,19.07℃和19.57℃,较其对应行间空气温度月均值分别降低了0.49℃,0.44℃和0.34℃,但未形成显著差异。自然生草和人工生草的树冠内空气温度略低于清耕,未形成显著差异(图3B)。
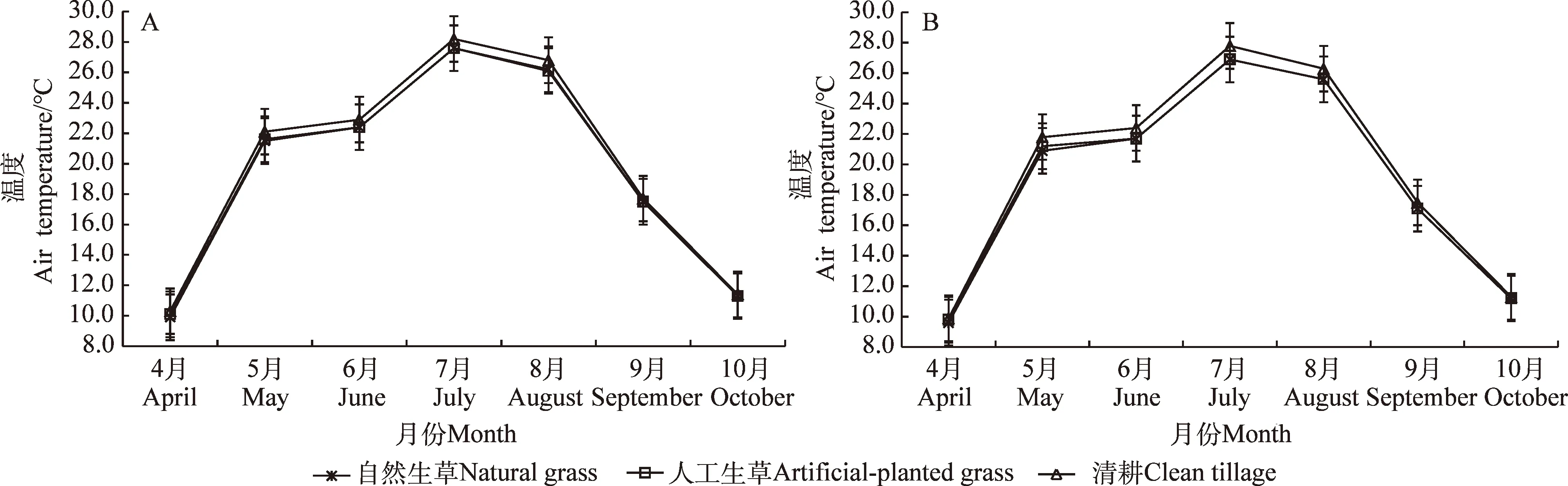
图3 不同处理行间及树冠内空气温度月变化
夏季自然生草、人工生草和清耕果树行间空气温度的日均值分别为28.71℃,28.76℃和31.00℃,自然生草和人工生草基本相同,均显著低于清耕(P<0.05,图4A)。
夏季自然生草、人工生草和清耕果树树冠内空气温度的日均值分别为28.07℃,28.13℃和29.83℃,较其对应行间空气温度日均值分别降低了0.64℃,0.63℃和1.17℃,与其对应行间空气温度未达到显著差异。夏季自然生草和人工生草的树冠内日均温基本相同,均显著低于清耕(P<0.05,图4B)。
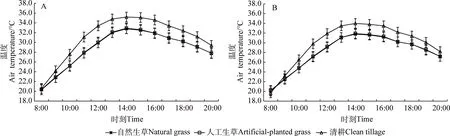
图4 不同处理行间及树冠内空气温度日变化
2.3 不同处理果园的土壤温度
自然生草和人工生草4月和10月的地表温度均显著低于清耕(P<0.05),5月至9月的均极显著低于清耕(P<0.01),且4月至10月的地表温度月均值也极显著低于清耕(P<0.01,表1)。
自然生草和人工生草4月和10月5 cm土层的土壤温度略低于清耕,与清耕无显著差异;6月的显著低于清耕(P<0.05),5月、7月、8月和9月的均极显著低于清耕(P<0.01),4月至10月的月均值显著低于清耕(P<0.05)。
自然生草和人工生草4月、5月和10月10 cm土层的土壤温度略低于清耕,与清耕无显著差异;6月至9月的均显著低于清耕(P<0.05),4月至10月的月均值也显著低于清耕(P<0.05)。
自然生草和人工生草4月、5月、6月、9月和10月15 cm土层的土壤温度略低于清耕,与清耕无显著差异;7月和8月的均显著低于清耕(P<0.05),4月至10月的平均值与清耕未形成显著差异。
自然生草和人工生草4月至10月20 cm土层的土壤温度及4月至10月的月均值均略低于清耕,与清耕之间无显著差异。
由表1可知,自然生草和人工生草对地表温度影响较大,随着土层深度的逐渐增加,其对土壤温度的影响逐渐减弱,且自然生草的影响力略高于人工生草。7月和8月人工生草的白三叶生长旺盛,对地表温度和5 cm,10 cm和15 cm土层的土壤温度影响略高于自然生草。

表1 不同处理不同月份不同土层的土壤温度
2.4 不同处理果园的光照强度及树冠反射光谱
夏季自然生草、人工生草和清耕果树行间光照强度的日均值分别为8.78×104Lx,8.83×104Lx和9.30×104Lx,自然生草和人工生草的较清耕分别降低5.59%和5.05%,均显著低于清耕(P<0.05,图5)。
不同处理处于同一地块,自然光光谱相同(见表2)。树冠反射光谱中,自然生草和人工生草的蓝紫光、红橙光、总辐射及红光/远红光基本相同,二者均极显著低于清耕(P<0.01,表2)。
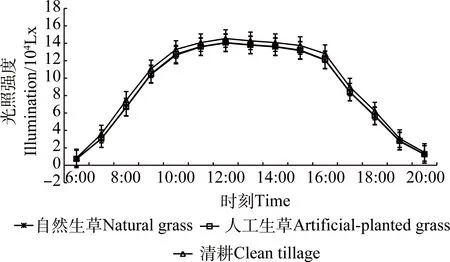
图5 不同处理行间的光照强度日变化

表2 不同处理的树冠反射光谱
2.5 不同处理的新梢尖削度、果实日灼率及早期落叶病状况
自然生草和人工生草均极显著促进了皇家嘎啦新梢的延长生长(P<0.01),但对新梢增粗生长无显著影响,因而自然生草和人工生草的新梢尖削度较清耕降低14.35%和11.21%,均极显著低于清耕(P<0.01,表3)。自然生草和人工生草均极显著降低了皇家嘎啦的果实日灼率(P<0.01),其中自然生草的果实日灼率显著低于人工生草(P<0.05)。自然生草和人工生草均极显著提高了皇家嘎啦早期落叶病的发病率及发病指数(P<0.01),加重了早期落叶病的危害程度(表3)。
自然生草和人工生草均极显著促进了惠民短枝新梢的延长生长(P<0.01),其中人工生草的显著高于自然生草(P<0.05);与清耕相比,人工生草极显著促进了惠民短枝苹果新梢的增粗生长(P<0.01),且人工生草的新梢粗度显著粗于自然生草(P<0.05);自然生草略微促进了新梢的增粗生长,与清耕未形成显著差异;自然生草和人工生草的新梢尖削度较清耕降低13.61%和12.91%,均极显著低于清耕(P<0.01,表3)。自然生草和人工生草均极显著降低了惠民短枝富士的果实日灼率(P<0.01),但极显著提高了惠民短枝富士早期落叶病的发病率及发病指数(P<0.01),不利于树体健康生长(表3)。

表3 不同处理对苹果新梢生长、果实果实日灼率和早期落叶病的影响
3 讨论
果树生长发育与果园小气候密切相关,果园小气候往往直接影响果实的产量及品质[32]。果园自然生草及人工生草后,将清耕果园的“土壤-大气”接触模式改为“土壤-地被植物-大气”的接触模式,果园的光、热、水、气等生态因子随之发生改变[4,7,32-34]。果园自然生草及人工生草后,由于地被植物蒸腾的土壤水分受果树树冠遮阴及阻拦的影响,大多数滞留于果树行间及树冠中,不易快速蒸散到大气中[35],因而果园行间空气湿度及树冠空气湿度均高于清耕。自然生草的地被植物(繁缕和牛繁缕)覆盖面积大(全园覆盖),覆盖度高且萌发早,枯黄迟[17],因而自然生草果树行间及树冠内的空气相对湿度略高于人工生草(行间覆盖);人工生草的地被植物(白三叶)在夏季生长旺盛,蒸腾强烈,因而人工生草在6月至8月果树行间和树冠内的空气相对湿度略高于自然生草。
同一气候区域其辐射平衡的主要差异取决于下垫面性状[33],果园自然生草及人工生草后提高了果园地面粗糙度,降低果园地面的有效辐射,同时果园地被植物进行光合作用,将太阳能转化为生物能贮藏于地被植物体中,且果园地被植物提高了果园空气相对湿度,增加空气热容量,减缓了果园空气温度上升幅度[36],因而自然生草及人工生草的果树行间空气温度及树冠空气温度均低于清耕。自然生草及人工生草在4月份并未提高果园空气温度,与梅立新与李会科[32]监测的结果及范兴海与黄寿波[36]所述不同,这可能与地被植物种类、地域环境不同有关。
土壤温度往往受大气温度、近地面的空间热平衡及土壤热特性等的影响[32]。本试验不同处理处于同一地块,土壤热特性和气候条件相同,由于自然生草和人工生草的地被植物可将太阳辐射能转化为生物能,具有较强的遮阴作用,可有效降低太阳对土壤表面的直接辐射,并且地被植物降低了果园空气温度,这些作用均对果园土壤有效辐射的吸收转化和土壤热量的传导产生较大的影响,因而自然生草和人工生草均降低了果园土壤温度,且其影响程度随土层深度的增深而降低[4,7,36-37]。
叶绿素光合作用过程中对蓝紫光和红橙光均有强烈的吸收作用[29],清耕果园地面无植被进行光合作用,因而自然生草和人工生草果园的蓝紫光、红橙光和总辐射强度均极显著低于清耕果园。光谱中红光与远红光的比值往往影响树体高度,当其比值较大时枝条的节间变短,树体矮化,当其比值变小时则枝条的节间伸长,树体乔化[29]。自然生草和人工生草果园反射光谱中的红光/远红光比值低于清耕果园,因而其新梢延长生长增强,尖削度降低,不利于花芽分化。自然生草和人工生草果园的光照强度降低,总辐射降低,因而有效降低了因强光辐射而引起的果实“日灼”病,但自然生草及人工生草后均提高了果园空气相对湿度,降低了果园光照强度,降低果园蓝紫光、红橙光、总辐射及红光/远红光比值,导致果园早期落叶病严重发生,且果园种植白三叶易引起红蜘蛛(Tetranychuscinnbarinus)爆发[12]。
虽然自然生草及人工生草均可改善果园的生态环境[38-39],提高果实品质及产量[4-8],但自然生草和人工生草不利于果树花芽分化,易引起病虫害发生,自然生草和人工生草的果园应加强果树修剪,加强病虫害的综合防治,特别是加强生长季节的修剪,促进树冠通风透光、新梢和叶片健壮生长以及花芽分化,以消除自然生草及人工生草带来的不利影响。
4 结论
自然生草对果园小气候的影响与人工生草对果园小气候的影响基本一致。与清耕相比,自然生草与人工生草均提高了果树行间及树冠内的空气相对湿度,降低了行间及树冠内的空气温度和果园土壤温度,且对果园土壤温度的影响随土层深度的增深而降低。自然生草与人工生草均降低了果园的光照强度、树冠反射光谱、新梢尖削度和果实日灼率,但提高了早期落叶病发病率及发病指数。自然生草与人工生草的果园应加强生长季节的修剪及病虫害防治,以消除自然生草与人工生草对果树生长带来的不利影响。
猜你喜欢
河北果树(2022年1期)2022-02-16
数学物理学报(2020年6期)2021-01-14
中国果业信息(2020年6期)2020-12-16
水土保持研究(2019年6期)2019-10-19
烟台果树(2019年1期)2019-01-28
烟台果树(2019年1期)2019-01-28
现代装饰(2018年1期)2018-05-22
辽宁林业科技(2017年4期)2017-06-22
浙江大学学报(工学版)(2016年2期)2016-06-05
浙江柑橘(2016年1期)2016-03-11
