
外源调节物质对铅胁迫下格木幼苗生理特性的影响*
2021-04-02 04:40:16孙明升杨章旗
林业科学 2021年2期
陈 旋 胡 颖 孙明升 贾 婕 杨章旗
(1.广西壮族自治区林业科学研究院 南宁 530002; 2.贵州省林业调查规划院 贵阳 550003; 3.广西大学林学院 南宁 530004;4.南京林业大学林学院 南京 210037; 5.广西优良用材林资源培育重点实验室 南宁 530002)
植物在自然环境中常受到干旱、低温和重金属等胁迫; 此外,环境污染和极端气候加剧了非生物胁迫对植物生长和发育的影响。土壤中许多重金属是植物生长所必需的微量元素,但其含量过高会破坏植物的生物膜,干扰植物正常代谢和生长,甚至导致植物死亡(刘铃等,2018; Ciarmielloetal.,2011)。非生物胁迫下,植物会改变自身形态和生理机能以提高对环境的适应性(Taizetal.,2015)。铅(Pb)胁迫时,植物可以将土壤中的铅活化,利用根细胞表面进行吸附,并通过细胞质膜将Pb2+转运至地上部分; 但只有少量Pb2+可借助共质体通路进行转载,大多数仍在土壤中累积并毒害植物,导致膜透性改变、酶活性扰乱、光合作用受限、呼吸作用受抑、有丝分裂受阻、内源激素紊乱等生理过程的改变,从而增加植物在萌发和出苗阶段的死亡率(Küpper,2017; 段德超等,2014)。环境保护部和国土资源部发布的《全国土壤污染状况调查公报》显示:中国土壤重金属总超标率高达16.1%,其中林地点位超标率为10.0%,中国南方地区是重要的铅矿区,部分地区土壤铅含量严重超出国家《土壤环境质量标准》规定的标准值,直接对植物的生存造成不利影响。因此,了解植物对铅胁迫的响应机制以探寻修复铅污染土壤的途径具有重要意义。
添加外源调节物质调控植物代谢生理是缓解铅胁迫造成植物损伤的有效方法。甜菜碱(betaine,BT)是细胞质渗透调节物质,参与逆境胁迫下稳定植物细胞内酶、蛋白结构和维持膜结构完整的生理过程(王三根等,2015),BT通过提高植物体内抗氧化酶活性,降低氧化胁迫并提高植物光合作用从而有效地缓解铅胁迫对玉米(Zeamays)(刘慧等,2017)和草棉(Gossypiumherbaceum)(Bharwanaetal.,2014)幼苗的伤害。茉莉酸甲酯(methyle jasmonate,MeJA)作为参与调控植物生长和发育的信号分子,可以应对干旱、高盐、重金属等非生物胁迫(杨华庚等,2011)。Bali等(2018)认为,MeJA通过降低Pb2+的吸收改善番茄(Lycopersiconesculentum)植株的生长、光合特性、抗坏血酸-谷胱甘肽循环活性、增加渗透物质和金属螯合化合物的含量,从而缓解铅胁迫的毒性效应。水杨酸(salicylic acid,SA)是能够激活植物过敏反应和系统获得性抗性的内源信号分子,Kohli等(2018)研究发现SA处理的芥菜(Brassicajuncea)幼苗通过促进生长、提高色素含量、调节抗氧化防御系统来对抗Pb2+的不良影响。钙作为植物必需的营养元素,具有稳定细胞膜结构并诱导特定基因表达以防止膜损伤和渗漏的作用,施加钙可以导致离子间的拮抗作用来降低植物对污染物的吸收,增强抗氧化酶活性、植物细胞膜的选择性吸收能力和光合产物的合成,从而降低铅(王芳等,2016)、镉(石贵玉等,2010)等重金属胁迫对植物的伤害。
格木(Erythrophleumfordii)属豆科(Leguminosae),分布于广东、福建、台湾以及广西钦州、合浦、小明山等地,是中国南方主要用材珍贵造林树种之一。格木木材质硬而亮,纹理致密,是著名硬木之一,可用于房屋建筑和高档家具(黄忠良等,1997); 同时其具有较高的药用价值(Lietal.,2004),是中国二级重点保护珍稀濒危植物之一。目前对格木的研究主要集中在播种育苗、栽培措施、造林技术等方面,对其抗逆性研究较少。鉴于此,本文以格木幼苗为材料,通过添加外源调节物质,探讨铅胁迫下格木生理特性的变化及其响应机制,为格木栽培与引种及重金属污染土壤的修复提供理论依据。
1 材料与方法
1.1 试验材料
1年生格木幼苗为广西林业科学研究院培育的实生容器苗,容器上下口径15 mm,高20 mm,基质为蛭石与营养土按1∶3混合而成的土壤(pH6.5~7.8)。2018年6月将幼苗移至广西大学林学院实验苗圃基地(108°22′E,22°48′N),大棚规格长50 m、宽8 m、高3 m。移苗后采用相同的水肥管理和栽培措施培育1个月,选取长势基本一致,无病虫害的幼苗[苗高(50±5) cm]进行试验。
1.2 试验设计
试验采取随机区组设计,将培植于基质中的格木幼苗以20株为1个处理,分为14个处理,其中CK1正常培育,CK2进行铅胁迫处理,B1、B2和B3分别用100、500和1 000 mg·L-1的BT处理后再进行铅胁迫处理,S1、S2和S3分别用25、100和200 mg·L-1的SA处理后再进行铅胁迫处理,J1、J2和J3分别用0.1、1和3 mmol·L-1的MeJA处理后再进行铅胁迫处理,C1、C2和C3分别用5、20和40 mmol·L-1的CaCl2处理后再进行铅胁迫处理。外源调节物质处理采取叶面喷施法,喷施时间为每日早上9:00,喷施以叶片正反面浸湿并刚有液体下滴为宜,连续喷施3日后,将12组外源调节物质处理的幼苗及CK2连同其营养杯同时浸入10 mmol·L-1的PbSO4溶液中持续铅胁迫处理10天,期间补充溶液以保证浸泡液的浓度。于胁迫后的第1、3和10天分别采样,各处理中随机选择10~15株幼苗均匀摘取同一部位叶片,用打孔器分别打取叶面中心3个相近点位的叶片(即3次重复)作为实验材料并保存于-80 ℃超低温冰箱备用。
1.3 试验方法
采用氮蓝四唑法测定超氧化物歧化酶(superoxide dismutase,SOD)活性,愈创木酚比色法测定过氧化物酶(peroxidase,POD)活性,紫外吸收法测定过氧化氢酶(catalase,CAT)活性,酸性茚三酮法测定脯氨酸(proline,PRO)含量,蒽酮比色法测定可溶性糖(soluble sugar,SS)含量,二辛可宁酸法测定可溶性蛋白质(soluble protein,SP)含量,硫代巴比妥酸反应法测定丙二醛(malondialdehyde,MDA)含量。上述指标的测定和计算按照苏州科铭生物技术有限公司提供的试剂盒及说明书进行。吸光值用酶标仪(INFINITE 200 PRO,Tecan Austria GmbH)测量。利用日产SPAD-502叶绿素仪参照李合生(2000)的方法测定SPAD值。参照史树德等(2011)采用外渗电导法测定RC(DDB-303A型便携式电导率仪,上海精密科学仪器有限公司)。各指标均进行3次重复测量,(处理C3在胁迫第10天时由于幼苗受害严重导致样本不足,仅测定其SPAD值)。
1.4 数据分析
用Excel整理数据制表,运用IBM SPSS Statistics 23.0中的one way ANOVA进行方差分析、Duncan’s法进行多重比较、Pearson线性相关系数进行相关性分析。Fuzzy综合评价法根据模糊数学综合决策原理,采用加权平均和最大原则。公式如下:
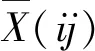
2 结果与分析
2.1 铅胁迫下外源调节物质对格木幼苗抗氧化酶活性的影响
2.1.1 对格木幼苗SOD活性的影响 胁迫1天时,CK1的SOD活性显著高于CK2(P<0.05),此时B2、J1、J3、C1、C3的SOD活性显著高于CK2(P<0.05),具有较敏感的SOD激活能力; 第3天时,CK2的SOD活性比CK1稍高,且较1天时显著升高(P<0.05),说明此时格木已改变体内代谢提升SOD活性以应答铅胁迫; 10天时,CK2的SOD活性显著高于CK1(P<0.05),S1、S2、J1和C1显著高于CK2(P<0.05)(表1)。
2.1.2 对格木幼苗POD活性的影响 由表1可知,CK1和CK2的POD活性随时间延后而逐渐升高,且CK2始终显著高于CK1(P<0.05),这说明铅胁迫时POD触发敏感、活性高且持续时间长,是格木抗铅胁迫生理的重要指标。胁迫1天时的B1、S1处理,胁迫3天时的B1、B2、S2、C2和MeJA处理,及胁迫10天时的S1处理表现出显著高于CK2的POD活性(P<0.05)。
2.1.3 对格木幼苗CAT活性的影响 CK1的CAT活性随时间明显增加,CK2则明显降低,胁迫1天和3天时,CK2显著高于CK1(P<0.05);胁迫10天时,CK2显著低于CK1(P<0.05)。以上结果表明,胁迫初期格木幼苗CAT酶活增强以响应外部胁迫,但CAT随胁迫时间延长而活性逐渐降低。B2、S1和S3在3天时表现显著高于CK2的CAT活性(P<0.05),J2在胁迫10天时有较强的CAT激活能力,S2在中期及后期均表现高度活性的CAT,B1、B3在中后期表现较CK2高的CAT活性,C2在1天和10天时的CAT活性显著高于CK2(P<0.05)(表1)。
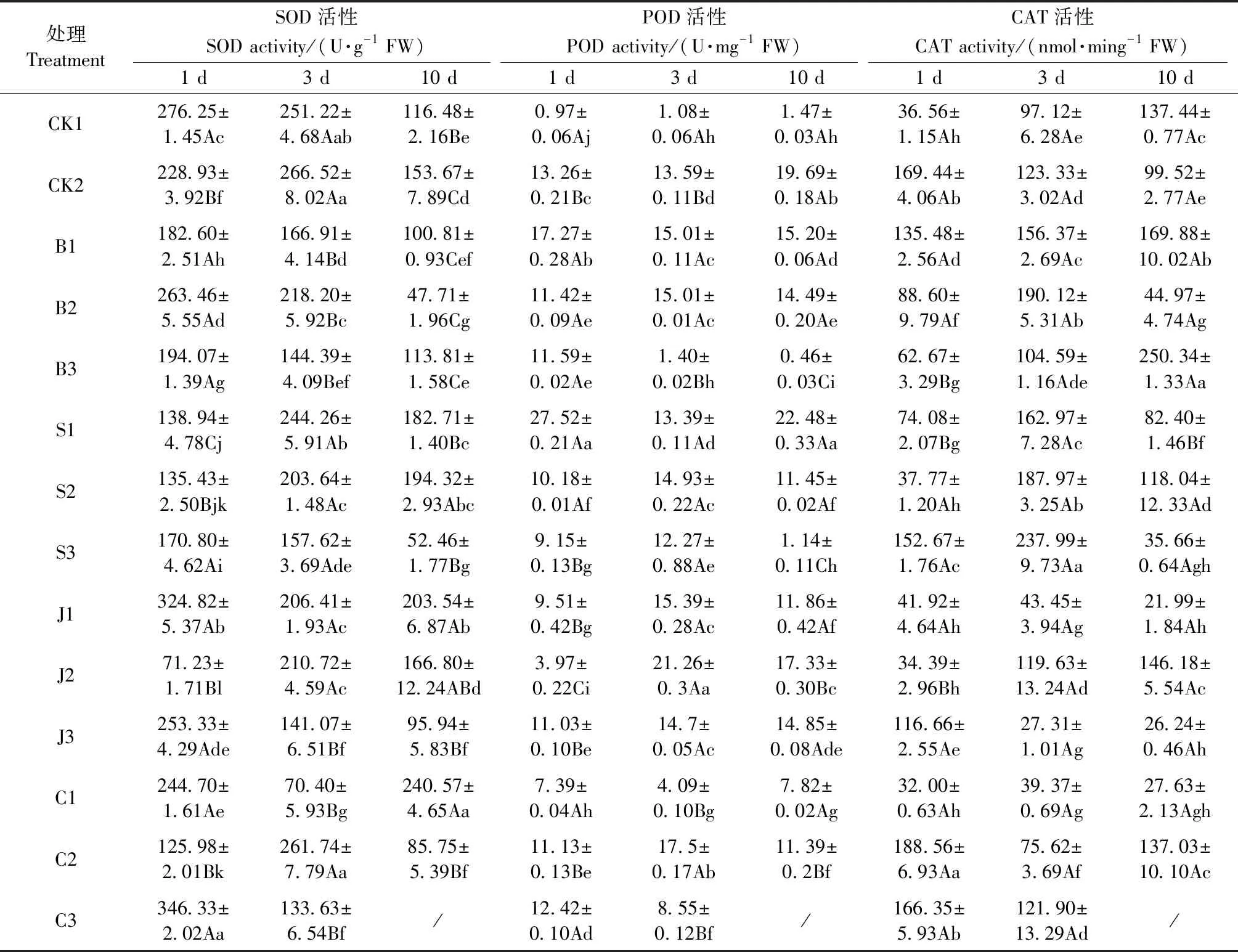
表1 铅胁迫下不同处理对格木幼苗抗氧化酶活性的影响①Tab.1 Effects of different treatments on the activities of antioxidant enzymes in Erythrophleum fordii seedlings under lead stress
2.2 铅胁迫下外源调节物质对格木幼苗渗透调节物质的影响
2.2.1 对格木幼苗PRO含量的影响 由表2可知,CK1和CK2的PRO含量随时间延后先降低后升高,CK2始终显著低于CK1(P<0.05),说明铅胁迫造成格木幼苗PRO的合成受抑制。铅胁迫1天时,除B1、S2、J3、C1和C2外,其余处理的PRO含量均显著高于CK2(P<0.05); 胁迫3天时,所有处理均显著高于CK2(P<0.05); 胁迫10天时,除B1、J3、C1和S3外,其余处理均对格木幼苗的PRO合成有显著促进作用(P<0.05)。
2.2.2 对格木幼苗SP含量的影响 CK1和CK2的SP含量随时间延后而显著升高又显著降低(P<0.05),CK2始终显著小于CK1(P<0.05),说明格木幼苗在遭受铅胁迫时可溶性蛋白的分解速率大于合成速率。胁迫1天时,除J1、J3和C3外的处理PRO含量均显著高于CK2(P<0.05); 胁迫3天时,S3、B2、B3、C1和C2分别比CK2高44.2%、6.7%、29.6%、51.6%和42.6%; 胁迫10天时,B1、B2和J2较3天时升高,B3和S3则呈减少趋势,但均显著高于CK2(P<0.05)。以上结果表明,BT、SA和CaCl2对铅胁迫格木幼苗SP的合成促进作用较强(表2)。
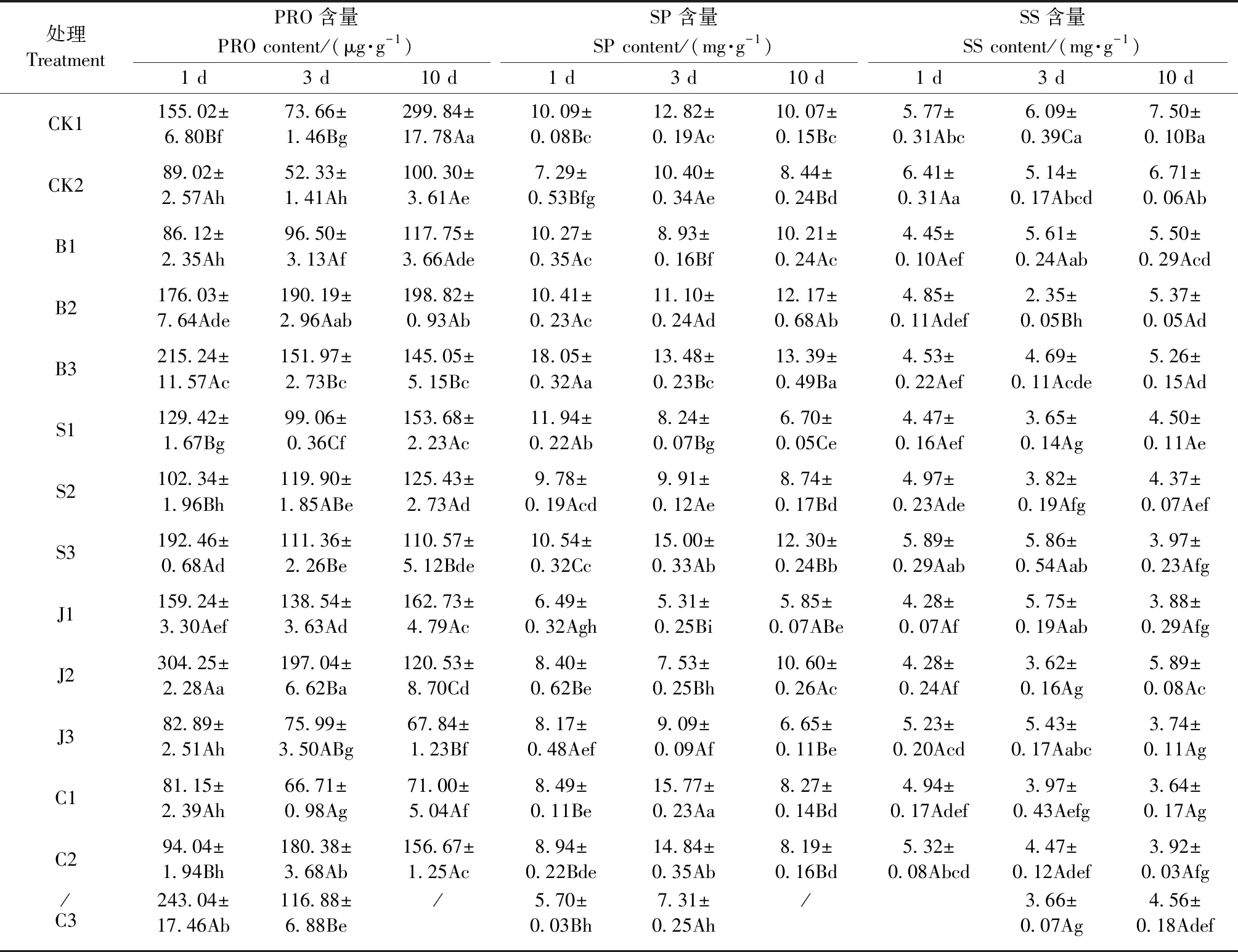
表2 铅胁迫下不同处理对格木幼苗渗透调节物质含量的影响Tab.2 effects of different treatments on the content of osmotic adjustment substance in E. fordii seedlings under lead stress
2.2.3 对格木幼苗SS含量的影响 由表2可知,胁迫1天时,CK2的SS含量显著高于CK1(P<0.05),且高于外源调节物质处理,说明铅胁迫下格木幼苗产生可溶性糖的应激反应较敏感; 胁迫3天时,B1、S3、J1和J3的SS含量略高于CK2; 胁迫10天时添加外源调节物质的处理均显著低于CK2(P<0.05); 且在胁迫3天和10天时CK2的SS含量均显著低于CK1(P<0.05),说明Pb2+累积对幼苗的破坏加重,导致光合效率降低或叶片内可溶性糖降解加快。
2.3 铅胁迫下外源调节物质对格木幼苗细胞膜透性的影响
2.3.1 对格木幼苗MDA含量的影响 由表3可知,CK2的MDA含量较随时间延后显著升高,在胁迫中后期显著高于CK1(P<0.05)。胁迫1天时,除S1、S2、J2、C1和C3外,其余处理的MDA含量均显著低于CK2(P<0.05); 胁迫3天时,除S1、J3、C1外的处理的MDA含量低于CK2,其中除B2和B3外均达显著差异(P<0.05); 胁迫10天时,除C1外的处理均表现显著抑制MDA合成的作用(P<0.05)。外源调节物质抑制格木合成MDA以抵御胁迫危害的作用。
2.3.2 对格木幼苗RC的影响 随胁迫时间延长,CK2的RC明显增加,由显著低于CK1,到与CK1相当,再到显著高于CK1(P<0.05)(表3)。铅胁迫1天时,外源调节物质处理的RC均显著高于CK2(P<0.05); 胁迫3天时,B1、B2、C1的RC稍低于CK2; 胁迫10天时,仅BT处理的RC显著低于CK2(P<0.05),其对RC的抑制作用逐步明显。
2.4 铅胁迫下外源调节物质对格木幼苗SPAD的影响
CK2的SPAD值随时间延长而降低且显著低于CK1(P<0.05),说明格木叶绿素的分解随Pb2+的富集增强。胁迫1天时,仅S1的SPAD值稍大于CK2; 胁迫3天时,除B2外的处理均显著高于CK2(P<0.05),其中S2、S3和J1分别较CK2高12、11和14倍; 胁迫10天时,B3、J1、J3、C1和施用SA处理的SPAD值仍显著高于CK2(P<0.05),其他处理则在胁迫后期有降低趋势(表3)。
2.5 外源调节物质对格木幼苗耐铅胁迫的综合评价
利用模糊隶属函数综合评价外源调节物质对格木幼苗耐铅胁迫的能力(表4)。各外源调节物质的耐铅胁迫能力在浓度上表现为B1>B2>B3,S3>S1>S2,J2>J1>J3,C2>C1>C3。所有处理综合评价排序为:J2>C2>B1>C1>J1>B2>CK2>S3>B3>S1>S2>J3>C3。格木幼苗产生适应性诱导以抵抗铅胁迫的外源调节物质以中低浓度的BT、MeJA、和CaCl2较为适宜。
表3 铅胁迫下不同处理对格木幼苗细胞膜透性及SPAD值的影响Tab.3 Effects of different treatments on cell membrane permeability and spad value of E. fordii seedling under lead stress
2.6 铅胁迫下格木幼苗生理指标的相关性
SOD活性与CAT活性呈显著正相关(P<0.05),与PRO和SS含量呈极显著正相关(P<0.01); POD活性与CAT活性呈显著负相关(P<0.05),与PRO、SP和SS含量呈极显著负相关(P<0.01); CAT活性与PRO和SS含量呈极显著正相关(P<0.01); PRO与SP、SS、RC及SPAD值呈极显著正相关(P<0.01); SP与SS呈极显著正相关(P<0.01); SS与RC呈显著正相关(P<0.05); MDA与RC呈显著正相关(P<0.05); RC与SPAD值呈极显著正相关(P<0.01); 各生理指标相互协同或拮抗,共同修复调节铅胁迫对格木的损伤(表5)。
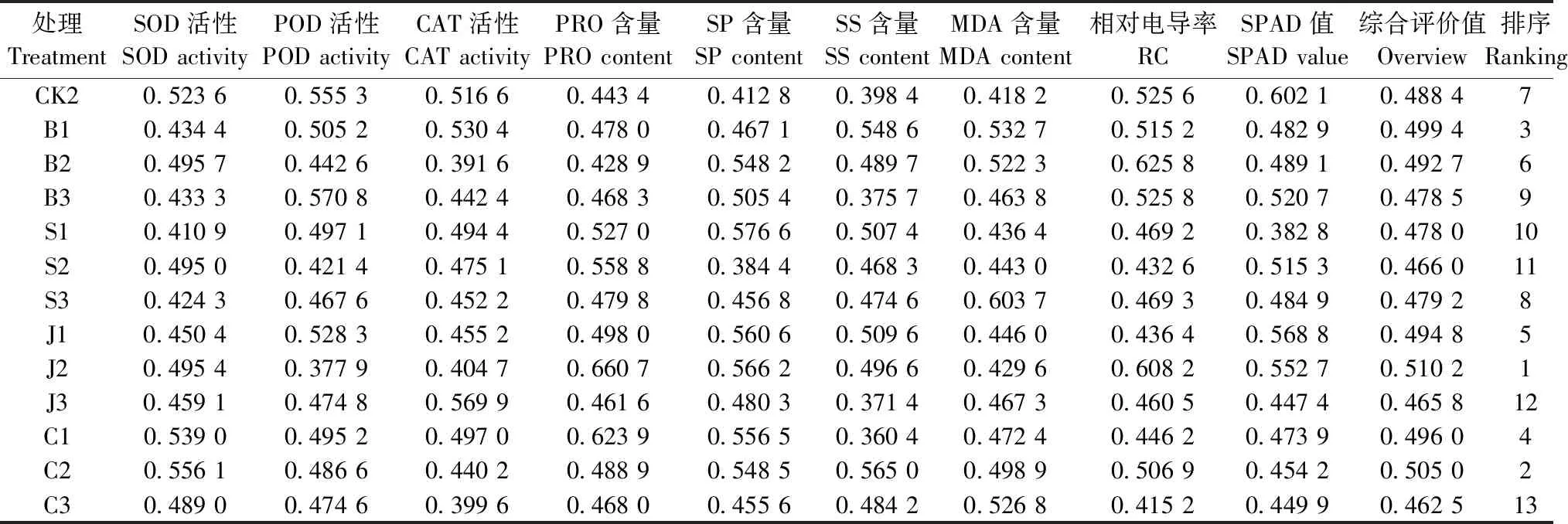
表4 铅胁迫下不同处理的格木幼苗的各项生理指标的隶属值及其综合评价Tab.4 membership value of various physiological indexes of different treatments of E. fordii seedlings under lead stress and its comprehensive evaluation
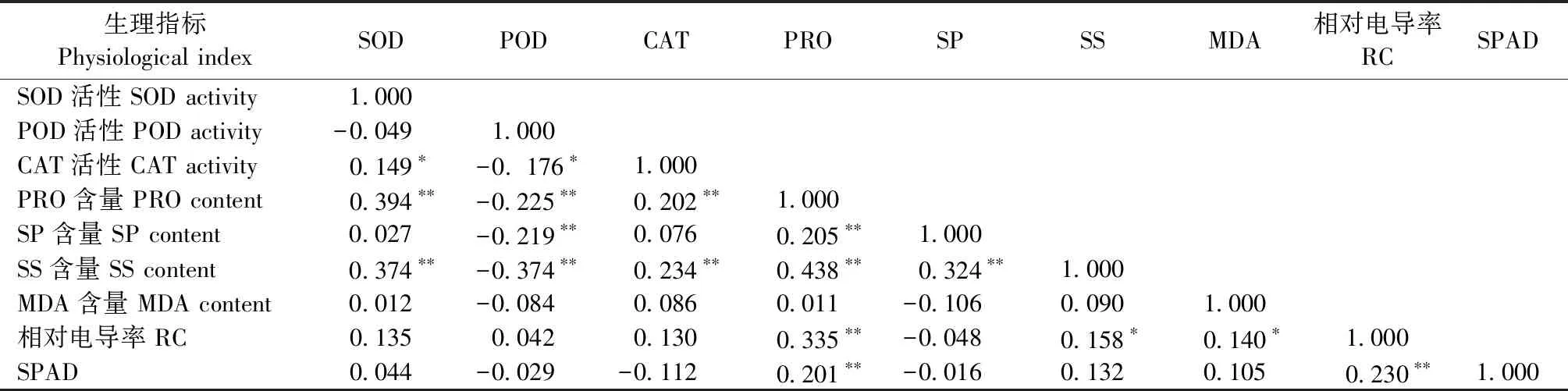
表5 铅胁迫下格木幼苗生理指标的相关性分析①Tab.5 Correlation analysis of E. fordii physiological indexes under lead stress
3 讨论
3.1 铅胁迫对格木幼苗的影响
铅胁迫会限制植物细胞有丝分裂,改变细胞膜透性,破坏叶绿体、线粒体、细胞核等亚显微结构,替换部分酶活性中心的金属离子以影响其活性,继而引起植物光合作用、呼吸作用、氮代谢、核酸代谢紊乱,同时造成植物体内元素失衡而导致营养胁迫(Kosobrukhovetal.,2004)。本研究发现,铅胁迫使格木幼苗的PRO含量显著减少,可溶性糖(SS)、可溶性蛋白质(SP)及光合色素的合成受到明显抑制,严重扰乱细胞维持其稳态的渗透压而增强细胞膜透性,这可能是Pb2+与膜蛋白上的-SH结合或与磷酸乙醇胺和磷脂丝氨酸反应,并伴有Pb2+与膜结合的Ca2+置换,从而改变细胞壁膜系统的弹性和可塑性(宋勤飞等,2006)。Pb2+的累积导致胞内MDA含量及相对电导率(RC)明显增加,同时刺激抗氧化酶活性增强而降低铅胁迫造成的脂质过氧化; 其中POD激活程度大、触发敏感且作用持久,是格木幼苗抗铅胁迫的主要抗氧化酶,这与常云霞等(2015)和周芙蓉等(2013)的研究相同。格木对Pb2+具有一定的吸收和转运作用,其通过质膜上ABC转运蛋白和负电荷对Pb2+的泵离、液泡中有机酸和生物碱与Pb2+的络合及有机配体(半胱氨酸、谷胱甘肽、植物螯合肽、金属硫蛋白)与Pb2+的螯合来缓解Pb2+对自身的毒害(段德超等,2014)。
3.2 外源调节物质对格木幼苗抗氧化酶活性的作用
铅会造成植物体内活性氧代谢加强而过氧化胁迫,使植物体内产生大量ROS(reactive oxygen species)而氧化细胞内核酸和蛋白质等生物大分子,导致体内代谢失衡并引起膜脂过氧化和膜结构破坏(Taizetal.,2015; 史树德等,2011)。SOD可以催化氧自由基歧化反应从而消除植物体内活性氧的累积,POD和CAT又将催化产生的歧化产物H2O2转化为无害的分子氧和水(史树德等,2011)。本研究结果表明,外源调节物质对3种抗氧化酶活性的激活程度不一,但均对抗氧化酶协同防御活性氧或其他过氧化物自由基起到促进作用,这与石贵玉等(2010)和常云霞等(2015)的研究结果一致。铅胁迫下3种抗氧化酶活性的诱导与激活有差异,是因在植物细胞中ROS有超氧化物(O2.-)、单线态氧(1O2)、过氧化氢(H2O2)和羟基自由基(OH·)等形式,其中SOD主要清除超氧化物阴离子,同时产生H2O2; POD则主要催化H2O2、氧化酚类和胺类化合物; CAT在过氧化物酶体及乙醛酸循环体中含量较高,负责清除由光呼吸或脂肪酸氧化过程中形成的过氧化物(Taizetal.,2015)。
3.3 外源调节物质对格木幼苗渗透调节物质的作用
逆境条件下,植物积累小分子有机化合物以增加细胞溶质浓度降低渗透势以保持体内细胞与组织的水分平衡,使体内与膨压有关的生理活动正常进行。PRO增强蛋白质的水合作用保持原生质形态,同时清除ROS保持膜结构的完整性(王三根等,2015); 可溶性蛋白质(SP)多为未与膜系统特异性结合的酶,有保护植物细胞中生物大分子和维持特定细胞结构的作用(曹莹等,2005); 可溶性糖(SS)能降低原生质的渗透势,利于细胞从外界吸水而产生具有固体机械特性的超饱和液体,避免细胞溶液结晶并防止细胞塌陷(Koster,1991; 李洁等,2009)。有研究表明,逆境条件下外施BT(Shaoetal.,2008)、SA(段小华等,2009)、MeJA(杨华庚等,2011)和Ca2+(王芳等,2016)能及时并显著提升PRO、SP和SS的合成以调节细胞渗透失衡。本研究结果显示,施加外源调节物质能较好缓解Pb2+造成的细胞渗透失衡; 胁迫初期,SP受外源调节物质触发而合成反应快; 胁迫中期,PRO的激活较为彻底,主导格木幼苗的渗透调节; 胁迫后期,SP和PRO共同均等地对渗透失衡进行修复调节。本研究外源调节物质对格木幼苗SS的合成无明显促进作用,这与邹文桐等(2012)和段小华等(2009)的研究结果相悖,但与谷文英等(2013)利用SNP对菊苣(Cichoriumintybus)果糖、葡萄糖和蔗糖含量的研究结论相似,可能是高浓度Pb2+对格木幼苗的小分子糖类物质和高分子量果聚糖的作用效果不一,导致可溶性糖总量存在差异。
3.4 外源调节物质对格木幼苗细胞膜透性的作用
铅胁迫时,植物体内产生强氧化自由基而诱发膜脂过氧化,导致膜脂分解和MDA累积,进而改变膜蛋白的性质和膜脂的流动性,细胞内电解质由于胞膜透性增大而外渗,继而增大植物细胞浸提液的RC。本研究中,格木体内MDA含量和RC随胁迫时间延长而明显增加,外源调节物质对减少MDA含量和RC的表达有差异,但均表现一定的分解MDA和降低RC的作用。这与赵腾飞等(2017)和Lou等(2015)的研究结果相同。本研究中Ca2+在胁迫中后期对MDA含量和RC的抑制作用较小,这与赵腾飞等(2017)和王芳等(2016)的研究结果有差别,可能是由于高浓度Ca2+使植物的蛋白质与核酸变性、原生质内含物外泄,增强对乙烯等内源激素的修饰和放大作用,从而加快植物的衰老和损伤。
3.5 外源调节物质对格木幼苗光合色素的作用
植物在铅胁迫下表现中毒症状与叶绿素的降解、叶绿体的功能障碍和光合作用受抑有关。铅胁迫破坏光合色素合成的原因可能是Pb2+沉积在类囊体并与膜蛋白体结合,置换其中的Mg2+,导致叶绿体酶系统破坏而阻碍叶绿素合成(Küpperetal.,1996)。在甜瓜(Cucumismelo)(左彬彬等,2018)、烟草(Nicotianatabacum)(卢军等,2011)和喜树(Camptothecaacuminata)(孟长军等,2018)的相关研究中表明,逆境胁迫对植物叶绿素合成的抑制作用明显,施加外源调节物质能有效提高叶绿素的含量而减少胁迫损伤。这与本研究中施加外源调节物质对铅胁迫下格木幼苗SPAD值的变化情况一致。
3.6 生理指标受外源调节物质的影响及其相关性
各生理指标之间存在一定的关联性:格木幼苗受铅胁迫时产生大量强氧化自由基,继而引起膜脂过氧化和MDA累积,高含量的MDA破坏细胞膜透性致使细胞质内含物外渗而RC激增。为抵御胁迫伤害,格木光合作用等代谢增强,加速碳水化合物、氨基酸和蛋白质等光合产物的合成,使胞内PRO和SP含量增加,渗透调节物质彼此协同促进并刺激SOD和CAT活性增强,清除胞内超氧化物阴离子自由基,同时消除植物减少自身损伤进行的光呼吸所形成的过氧化物。高活性的CAT清除大部分过氧化物,而光呼吸消耗POD将光合作用的副产物乙醇酸氧化为乙醛酸和H2O2,故POD活性与CAT活性呈负相关关系。
本研究中,BT对激活CAT活性、促进PRO和可溶性蛋白质(SP)合成、消除MDA累积、降低相对电导率(RC)的作用最大; SA对PRO和SP含量及SPAD值的提高有明显作用,能在一定程度提升POD活性; MeJA对增加PRO含量的作用较大,对促进光合色素合成的作用最明显; CaCl2能有效促进PRO和SP的合成和激活SOD活性,但一定程度增大MDA含量和RC。外源调节物质对铅胁迫诱导产生抗性生理从而提高格木幼苗抗铅胁迫的能力有差别,主要是由于外源调节物质的抗铅胁迫机制不同。BT是极溶性生物碱,对植物亚细胞、大分子生物物质结构和生物膜有保护作用,主要参与植物渗透调节,通过改变多糖代谢,影响植物根细胞壁中果胶的成分以增强细胞壁对重金属的结合能力(段德超等,2014),同时改变土壤重金属形态、生物有效性和无机离子的吸收转移,从而增强植物对土壤重金属的吸收和累积(姚伟卿等,2019)。SA是植物体自身可合成的小分子酚类物质,通过调控信号分子代谢酶的活性而积累H2O2,H2O2作为第二信使诱导保护酶的合成(Raskin,1992)。MeJA在胁迫中是驱动植物防御基因表达的信号因子,调节植物体保护酶及次生代谢酶活性,同时诱导特异基因表达以合成与植物细胞防御病菌有关的蛋白质及次生物质(Lehmannetal.,1995)。Ca2+与膜磷脂的极性头部结合,通过交联作用使膜脂分子和蛋白质分子结合紧密,从而降低膜的透性(李美如等,1996)。
4 结论
铅胁迫导致格木幼苗渗透调节物质含量减少、光合色素合成受抑、MDA含量和相对电导率(RC)增加、抗氧化酶活性增强,格木在受铅胁迫损伤的同时亦表现一定的耐铅胁能力。施加外源调节物质使抗氧化酶活性明显增加、PRO和可溶性蛋白质(SP)含量明显升高、叶绿素合成速率加快、MDA降解、RC降低,但对可溶性糖(SS)的合成促进作用不明显。外源调节物质有利于提升格木幼苗抗铅胁迫的抗性,各处理抗铅胁迫能力综合表现为J2>C2>B1>C1>J1>B2>CK2>S3>B3>S1>S2>J3>C3。中低浓度的甜菜碱(BT)、茉莉酸甲酯(MeJA)和CaCl2对提高格木幼苗抗性、抵抗铅胁迫损害的效果较好。抗性生理指标间存在一定的相关性,彼此协同或拮抗,共同修复调节铅胁迫伤害。
猜你喜欢
安徽农学通报(2024年1期)2024-01-30 10:04:32
舰船科学技术(2022年11期)2022-07-15 07:51:56
今日农业(2021年10期)2021-11-27 09:45:24
中南林业科技大学学报(2020年5期)2020-06-06 08:17:50
西藏农业科技(2019年3期)2019-11-04 00:35:10
现代园艺(2018年3期)2018-02-10 05:18:12
上海农业学报(2017年3期)2017-04-10 12:39:12
广西林业科学(2016年1期)2016-03-20 05:33:03
中国火炬(2015年12期)2015-07-31 17:38:35
中国民族医药杂志(2015年7期)2015-07-13 02:13:38
