
叉角厉蝽对烟草上斜纹夜蛾搜索效率影响因子的研究
2021-03-30 02:25:22陈雪梅谷星慧冼继东张立猛范悦莉黄智华周文兵赵进龙张翠萍
环境昆虫学报 2021年1期
陈雪梅,谷星慧,冼继东*,张立猛,范悦莉,黄智华,周文兵,赵进龙,张翠萍
(1.华南农业大学植物保护学院,广州 510642;2.云南省烟草公司玉溪市公司,云南玉溪 653100;3.烟草行业病虫害生物防治工程研究中心,云南玉溪 653100)
叉角厉蝽Eocantheconafurcellata(Wolff)属半翅目Hemiptera蝽科Pentatomidae,可捕食鳞翅目Lepidoptera、鞘翅目Coleoptera和半翅目Hemiptera等多种农作物害虫。由于叉角厉蝽发生早、时间长、捕食量大、分布范围广、活动能力强,因此被认为是一种极具应用潜力的捕食性天敌(Rani and Wakamura,1993;Revannavar,2004;Ray and Khan,2011),在印度和中国台湾叉角厉蝽都被作为一种重要的捕食农作物上害虫的天敌昆虫(Pillai and Agnihotri,2013;Sahayaraj,2014)。国内外对叉角厉蝽在生物学以及饲养方面进行了较多研究,朱涤芳(1990)报道了叉角厉蝽的一些生物学特性;李丽英等(1988)对叉角厉蝽的人工饲料进行了研究,发现用大蜡螟幼虫和米蛾幼虫饲养,可以完成正常繁殖;Yasuda and Wakamura(1992)报道了可用冷冻的斜纹夜蛾Spodopteralitura(Fabricius)幼虫饲养叉角厉蝽;何旭诺等(2013)的研究表明,用黄粉虫作为叉角厉蝽的饲料更方便经济。随着人工大量繁殖技术的研究推进,叉角厉蝽作为一种重要的生物防治因子在田间应用已成为可能。斜纹夜蛾是烟草上的常见重要害虫,以幼虫咬食烟叶,形成虫孔、缺刻甚至仅剩叶脉,由于该虫具有暴食性,如果防治不当会对烟草造成毁灭性损失(贤小勇,1995;陈代利等,2013)。目前对于烟草上的斜纹夜蛾仍以化学防治为主(孙光军等,2003;庞云红等,2007;汤心砚等,2018),而利用捕食性天敌叉角厉蝽防治烟草上的斜纹夜蛾,将可有效地解决化学防治带来的“3R”问题。研究表明,叉角厉蝽对斜纹夜蛾具有极强的捕食能力(Yasuda and Wakamura,1996;Kumar and Singh,2007),且更倾向于捕食斜纹夜蛾的中、高龄幼虫(蒋杰贤和梁广文,2001)。因此,叉角厉蝽可用于斜纹夜蛾的生物防治。
利用天敌昆虫来防治害虫,是害虫生物防治中的一个重要手段。提高天敌昆虫在田间控害效果,降低经济成本是天敌昆虫推广应用的重要保证(Debach and Rosen,1991;Morrisetal.,2004;Tylianakisetal.,2007)。在目前的实际应用中,利用天敌昆虫防治害虫在田间控害效果往往不稳定,从而影响生产效益,使得天敌难以在田间推广应用(陈学新,2013)。而想要有效地推广应用叉角厉蝽防治烟草上的斜纹夜蛾,一方面需要明确叉角厉蝽对斜纹夜蛾的捕食潜能,以确定田间释放密度;另一方面还要确定叉角厉蝽在烟草上对斜纹夜蛾的搜索效率,以确定田间释放方法。陈然等(2015)在室内研究了叉角厉蝽对斜纹夜蛾的捕食潜能,得到单头叉角厉蝽对斜纹夜蛾的捕食能力,为田间释放密度提供了依据。但是,叉角厉蝽在烟草上对斜纹夜蛾的搜索效率方面的研究尚未见报道。本文采用二次通用旋转组合设计,研究了在烟草上斜纹夜蛾的密度、叉角厉蝽释放数量和烟草株数这3个因子对叉角厉蝽捕食效率的影响,并分析了单因子主效应以及双因子效应,旨在为烟田上人工释放叉角厉蝽防治斜纹夜蛾提供科学的依据。
1 材料与方法
1.1 材料
叉角厉蝽:叉角厉蝽的卵采自于广州华南农业大学试验基地的甘薯叶上,待卵孵化后于室内温度26±1℃,相对湿度75%±5%,光周期14 L ∶10 D的人工培养箱中,以斜纹夜蛾幼虫喂饲至多代,备用。
斜纹夜蛾:采用陈其津等(2000)的人工饲料配方进行饲养,室内饲养条件同叉角厉蝽。
烟草NicotianatabacumL.:由云南省烟草公司玉溪市公司提供的实生烟苗一批,移至直径为16.5 cm的塑料花盆中,待烟草新长出5张叶片后用于试验。
1.2 方法
1.2.1试验设计
试验采用二次通用旋转组合设计,选择叉角厉蝽释放数量、斜纹夜蛾幼虫密度和烟草植株数量3个因子作为参试因子,以斜纹夜蛾幼虫被捕食量为目标函数,根据丁希泉(1986)试验设计要求共设20个处理,各处理按参试因子的编码值进行一一组合,重复3次。参试因子的水平及编码值如下(表1)。

表1 叉角厉蝽搜索效应研究的参试因子水平及编码值
1.2.2试验方法
试验前将供试的叉角厉蝽4龄若虫单头隔离后,先饱食24 h,然后饥饿24 h,使之基本处于相同的饥饿水平,作为被试者备用。试验时,按试验设计的要求,在每个处理的养虫笼(规格为50 cm ×50 cm ×50 cm)中分别放入相对应数量的烟草盆栽苗、斜纹夜蛾3~4龄幼虫、叉角厉蝽4龄若虫,并在容器内放入蘸水棉花团,供叉角厉蝽补充水分,24 h后观察记录斜纹夜蛾被捕食的情况,并计算出叉角厉蝽的捕食率。所有斜纹夜蛾幼虫密度的处理均设有不接天敌的空白对照,以校正斜纹夜蛾被天敌捕食的数量。试验是在变温条件下进行的,试验期间日平均温度范围为25~33℃,平均气温26.8℃。
1.3 数据处理与分析
应用Excel表格进行试验数据初步整理归纳。应用DPS 9.50专业版和SAS 9.2版,对试验结果进行回归分析和回归模型的失拟性检验、显著性检验以及对各个因子回归系数进行t检验。
2 结果与分析
2.1 不同因子与叉角厉蝽搜索效率关系模型的建立
各参试因子试验结构矩阵和对叉角厉蝽捕食效率的影响结果(表2),其中Y代表叉角厉蝽的捕食率,本实验中以捕食率代表相对应的搜索效率。对试验数据进行逐步回归和中心化转换,建立叉角厉蝽搜索效率(Y)和叉角厉蝽数量(X1)、斜纹夜蛾幼虫密度(X2)和烟草植株数量(X3)因素之间的二次多项式回归模型如下:
Y=67.82809+6.74050X1-4.92690X2-1.22581X3-9.22848X12-1.32656X22-9.98862X32+4.17500X1X2+8.17500X1X3+0.87500X2X3
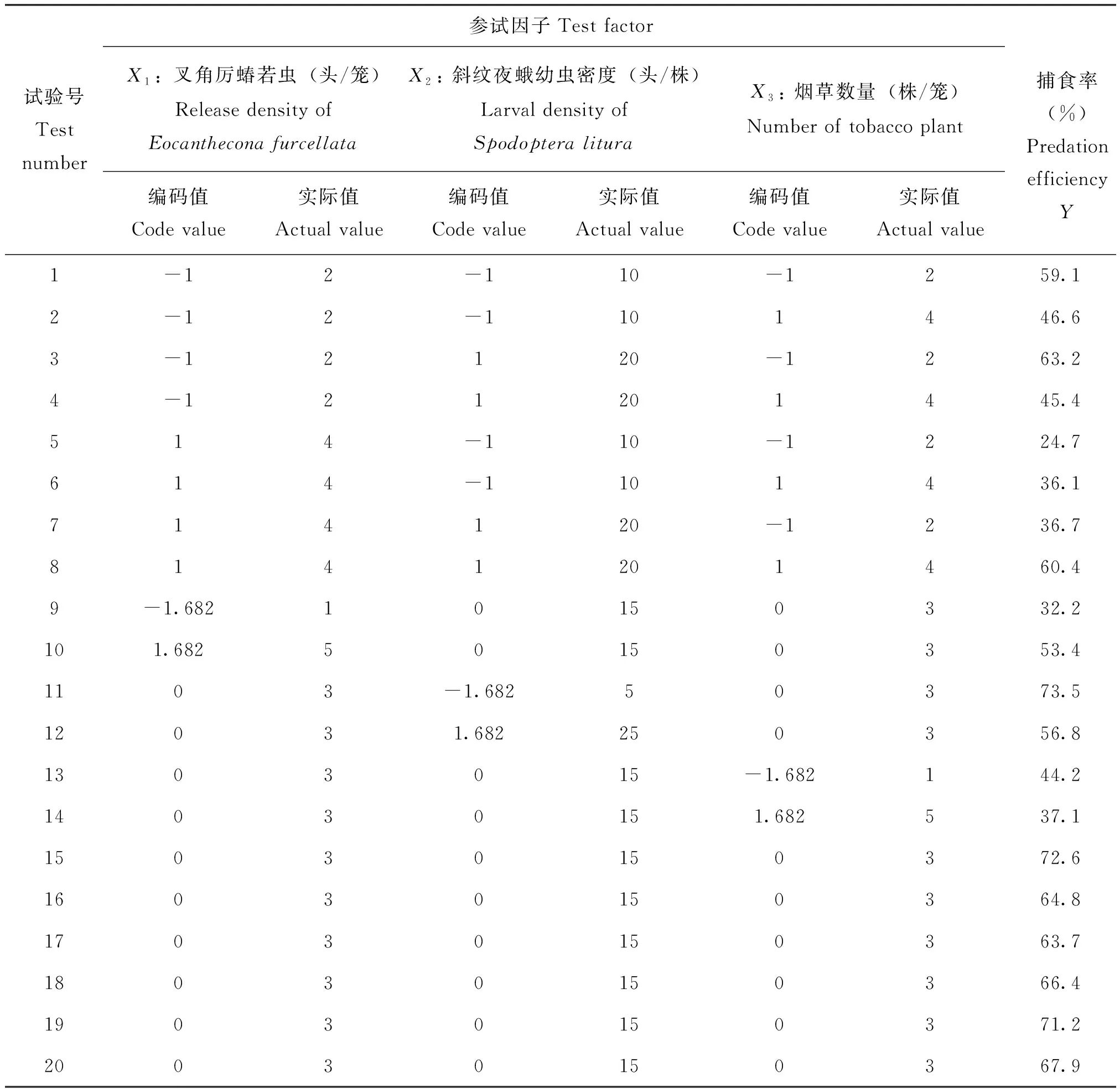
表2 不同因子处理下叉角厉蝽对斜纹夜蛾的捕食率
对模型进行失拟性检验和回归显著性检验,失拟性检验中的F1=0.9572
模型中各回归系数t检验及概率值P结果显示,模型的回归项b1、b2、b1b1、b3b3、b1b2、b1b3的显著水平均小于0.01,表明上述各回归项与叉角厉蝽捕食效率之间极显著相关(表4)。说明由参试因子X1和X2代表的叉角厉蝽释放数量和斜纹夜蛾幼虫密度对天敌的捕食效率有着极显著的影响,在交互项中的X1X2和X1X3回归项达0.01显著水平,说明叉角厉蝽释放数量与斜纹夜蛾幼虫密度、叉角厉蝽释放数量与烟草植株数量的交互作用对搜索效率也有显著影响。而b3、b2b2和b2b3达到0.5的显著水平,说明因子烟草植株数量(X3)和斜纹夜蛾幼虫密度与烟草植株数量的交互作用对叉角厉蝽的搜索效率也有一定的影响。整个模型拟合的相关系数为0.9720,F值为37.1389,极显著相关,即该模型能较好的拟合各参试因子与叉角厉蝽搜索效率的关系。

表3 回归模型的失拟性检验和显著性检验结果
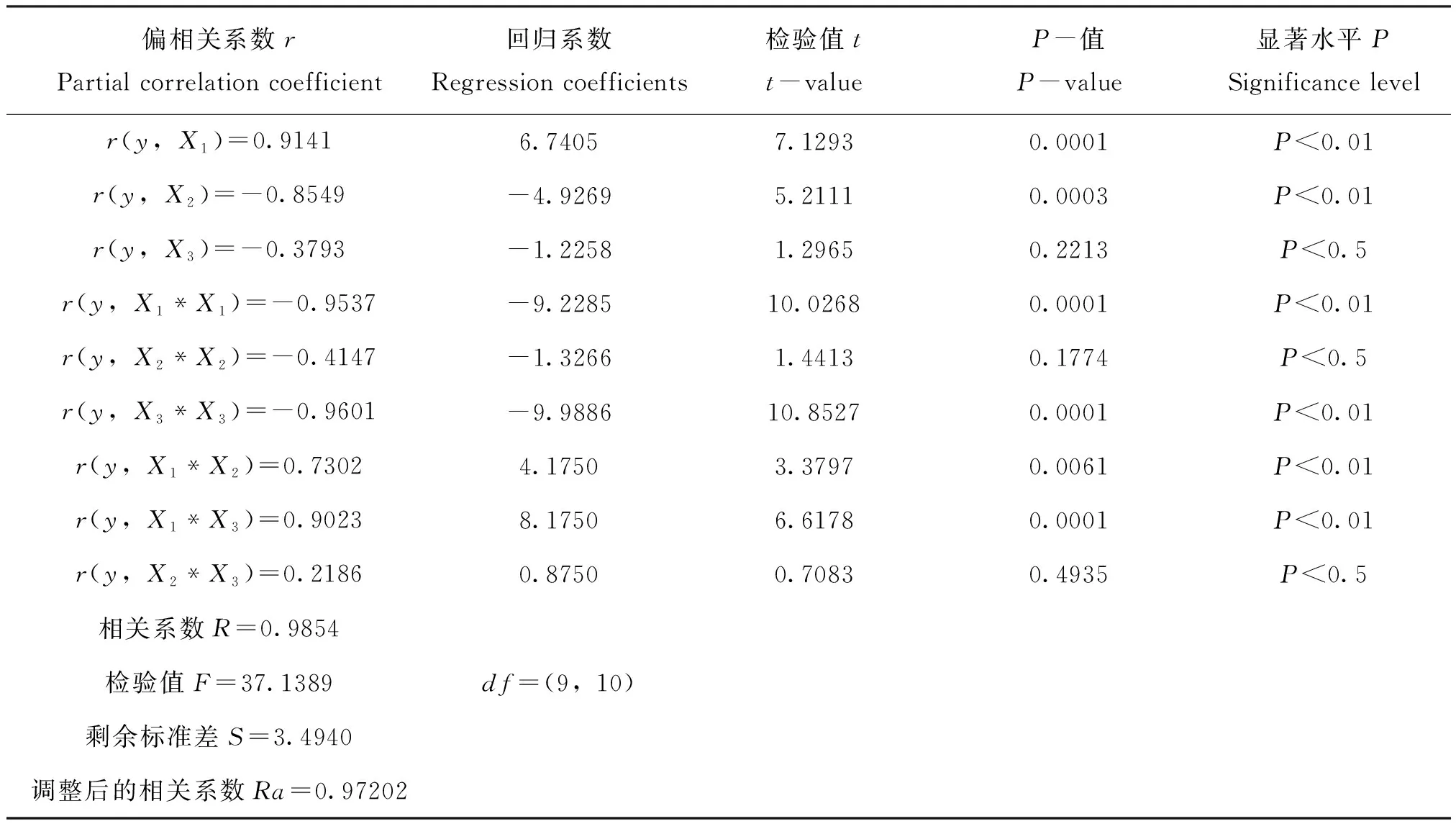
表4 各个因子回归系数t检验结果
2.2 影响叉角厉蝽搜索效率的单因子效应分析
用降维法求得叉角厉蝽的搜索效率与叉角厉蝽的数量、斜纹夜蛾和烟草植株数量各主效应的偏回归解析因子模型如下:
Ys1=67.82809+6.74050X1-9.22848X12X1:叉角厉蝽释放量
Ys2=67.82809-4.92690X2-1.32656X22X2:斜纹夜蛾密度
Ys3=67.82809-1.22581X3-9.98862X32X3:烟草植株数量
从模型分析中可以看出,在线性项中,对叉角厉蝽搜索效率的影响依次为叉角厉蝽释放数量(X1)>斜纹夜蛾幼虫密度(X2)>烟草植株数量(X3),其中叉角厉蝽数量对搜索效率的影响最大,且系数符号为正,表明该因子对其捕食效率呈现正影响,而斜纹夜蛾密度和烟草植株数均为负影响。
叉角厉蝽的搜索效率与叉角厉蝽释放数量(X1)和与烟草植株数(X3)的关系曲线均为开口向下的抛物线(图1),表明叉角厉蝽释放数量和烟草植株数量这两个因素对搜索效率的影响存在一个合理的范围,即叉角厉蝽释放数量在编码值为0-1(3~4头/笼)的范围天敌的捕食率最佳,低于或高于该范围的编码值捕食率下降;烟草植株数量在编码值为-0.5(2.5株/笼)和0.5(3.5株/笼)的范围天敌的捕食率最佳,低于或高于该范围的编码值捕食率下降。斜纹夜蛾数量(X2)与叉角厉蝽捕食效率的关系曲线较平直接近线性,表明斜纹夜蛾数量这个因子对捕食效率的影响较明显,随着斜纹夜蛾数量的增加,搜索效率逐渐下降。
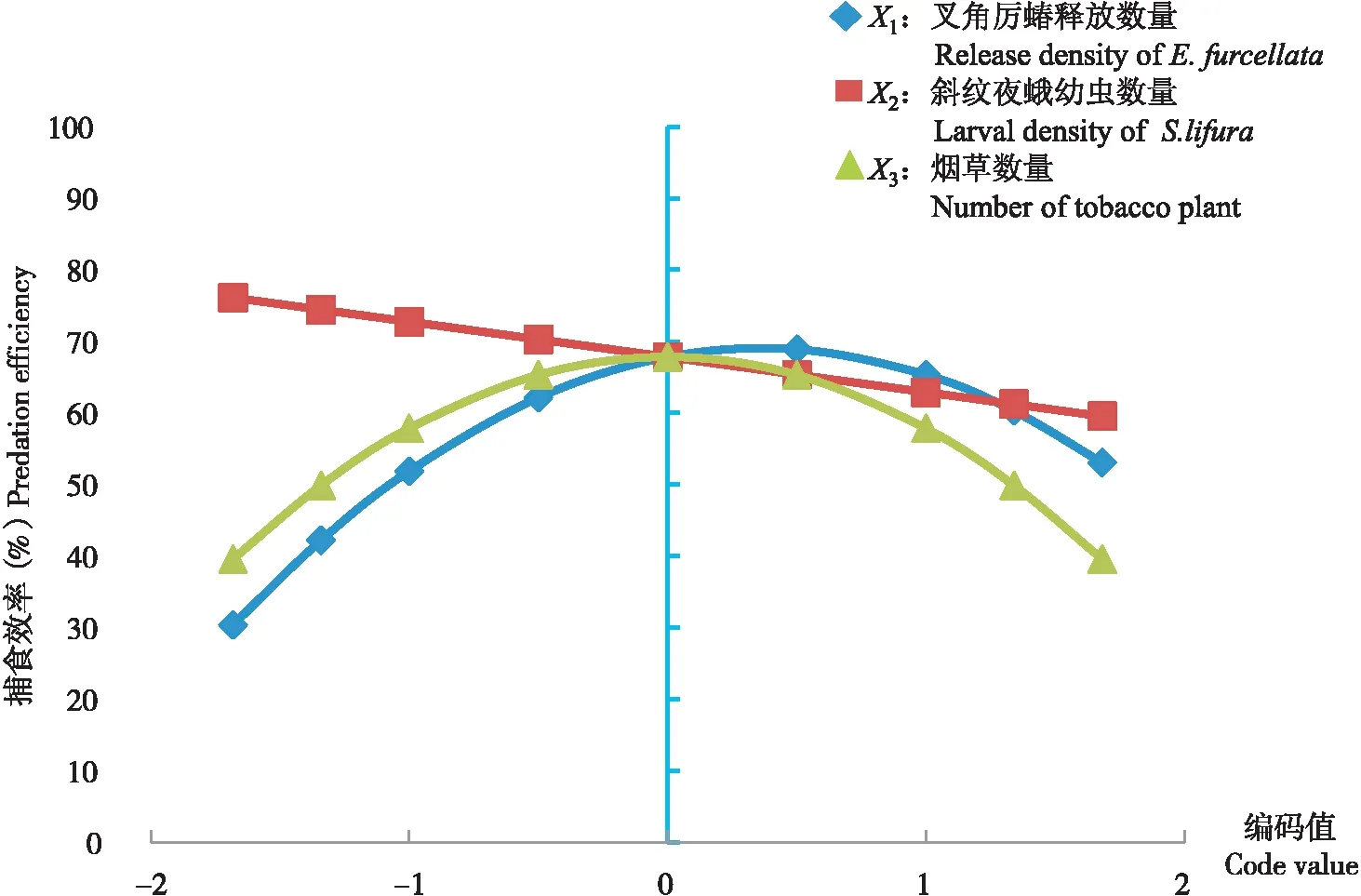
图1 单因子效应分析Fig.1 Analysis on single-factor effect
2.3 影响叉角厉蝽捕食效率的交互效应分析
叉角厉蝽释放数量(X1)、斜纹夜蛾幼虫密度(X2)和烟草植株数量(X3)三个因子之间的交互作用对叉角厉蝽搜索效率(Y)的影响如图2所示。
其中,叉角厉蝽搜索效率(Ys1,2)与叉角厉蝽释放数量(X1)和斜纹夜蛾密度(X2)的关系式如下:Y=67.82809+6.74050X1-4.92690X2-9.22848X12-1.32656X22+4.17500X1X2,相应的影响叉角厉蝽搜索效率的双因子(X1,X2)效应分析表明,叉角厉蝽的搜索效率随着叉角厉蝽数量的增加而增加(图2A),在增加到一定阶段又在缓慢下降,在斜纹夜蛾幼虫密度较低时,增加幅度较大,随着斜纹夜蛾幼虫密度的增加,搜索效率对叉角厉蝽释放数量的依赖程度在逐渐减弱。
叉角厉蝽搜索效率(Ys1,3)与叉角厉蝽释放数量(X1)和烟草植株数量(X3)的关系式如下:Y=67.82809+6.74050X1-1.22581X3-9.22848X12-9.98862X32+8.17500X1X3,相应的影响叉角厉蝽搜索效率的双因子(X1,X3)效应分析表明,随着叉角厉蝽释放数量和烟草植株数量的同时增加,叉角厉蝽的搜索效率呈现先增加后下降的趋势(图2B),当两个因子的编码值达到0(3头/笼)时或接近1(4头/笼)时,搜索效率也达到最大值,此后,随着烟草植株数量的加大,搜索效率对叉角厉蝽数量的依赖程度变小,而不再随着蝽量的增加而增加,反而呈下降的趋势,说明烟草植株数量对叉角厉蝽的搜索效率也存在一定的影响。
叉角厉蝽搜索效率(Ys2,3)与斜纹夜蛾密度(X2)和寄主植物植株数量(X3)的关系式如下:Y=67.82809-4.92690X2-1.22581X3-1.32656X22-9.98862X32+0.87500X2X3,由此关系式得到影响叉角厉蝽搜索效率的双因子(X2,X3)效应分析表明,在斜纹夜蛾幼虫低密度时,叉角厉蝽搜索效率随着烟草植物植株数量的增加而增加(图2C),当编码值达到0(3头/笼)时,叉角厉蝽的捕食率最大,而随着斜纹夜蛾密度的增加,减缓了这种增加的趋势。

图2 交互效应分析Fig.2 Analysis on interactive effect
3 结论与讨论
利用天敌昆虫防治害虫是害虫生物防治中的一个重要手段,研究阐明天敌昆虫对目标害虫的控害机理,是应用天敌昆虫防治害虫的首要基础(Debach and Rosen,1991;Morrisetal.,2004;陈学新等,2013)。研究明确捕食性天敌的捕食能力与搜索行为的影响因子,是提高天敌昆虫在田间控害效果的重要依据。本试验采用三因子二次回归旋转组合设计研究了影响叉角厉蝽搜索效率的主因子效应和双因子效应,结果表明,叉角厉蝽释放数量是影响其搜索效率的主要因素,斜纹夜蛾幼虫密度和烟草植株数量依次也对搜索效应产生显著的影响;因子之间的交互作用对搜索效率有一定的影响,但影响不如单独因子的作用大;三个独立因子并不单纯的与搜索效率呈正相关或负相关。这些结论与邓海滨等(2015)所报道的红彩真猎蝽Harpactorfuscipes对烟蚜Myzuspersicae的捕食作用及寻找效应的研究结果相似。张晓军等(2016)在研究蠋蝽Armachinensis对榆紫叶甲Ambrostomaquadriimopressum的捕食作用中发现,不同榆紫叶甲幼虫密度梯度下蠋蝽的捕食量不受影响,而高卓等(2012)报道,蠋蝽捕食量不仅与其虫龄高低有关,而且与猎物虫龄及体躯大小相关程度明显。许庆辉等(2014)的研究表明,烟盲蝽Nesidiocoristenuis对烟蚜的捕食量随着猎物密度增大而增多,且低密度下猎物被捕食的风险明显大于高密度下的猎物,本试验结果中,叉角厉蝽对烟草斜纹夜蛾幼虫的搜索效率随着斜纹夜蛾幼虫密度的增加而降低,这与黑刺益蝽Podisusnigrispinus及其他捕食者搜寻效应一致(Zanuncioetal.,2008;邓海滨等,2015)。在研究捕食性天敌的捕食潜能时,往往没有考虑猎物的寄主植物(高卓等,2012;许庆辉等,2014;邓海滨等,2015;张晓军等,2016)对天敌捕食行为的影响,本试验研究表明,在叉角厉蝽数量、斜纹夜蛾幼虫密度和烟草植株数量三个因子的相互干扰下叉角厉蝽对斜纹夜蛾的搜索效率不同,各个因子的改变所产生的效应也会随之改变,明确各因子改变所产生的干扰反应的大小,这为叉角厉蝽在田间的使用效果、放蝽的数量和质量、田间释放面积、斜纹夜蛾密度情况等都起着至关重要的作用。但本实验是在室内罩笼条件下完成,至于在田间的释放效果还有待进一步验证。
猜你喜欢
ELLE世界时装之苑(2022年11期)2022-05-30 20:23:43
ELLE世界时装之苑(2022年10期)2022-05-30 08:36:21
基层中医药(2021年4期)2021-07-22 07:15:32
今日农业(2020年17期)2020-12-15 12:34:28
今日农业(2020年13期)2020-12-15 09:08:51
——管氏肿腿蜂的应用技术
江苏林业科技(2020年3期)2020-08-31 01:00:08
今日农业(2019年13期)2019-08-12 07:59:08
现代园艺(2017年21期)2018-01-03 06:42:15
小天使·六年级语数英综合(2014年10期)2014-10-20 07:03:37
华东师范大学学报(自然科学版)(2014年1期)2014-04-16 02:54:58
