
高纺锤树形下2个中间砧富士苹果生长与叶片光合特性的比较
2021-03-26 08:08蔡华成王淑婷张小军高敬东杜学梅李春燕郝燕燕杨廷桢
华北农学报 2021年1期
王 骞,蔡华成,王淑婷,张小军,高敬东,杜学梅,李春燕,郝燕燕,杨廷桢
(1.山西农业大学 果树研究所,山西 太谷 030800;2.山西农业大学 园艺学院,山西 太谷 030801)
矮砧集约高效栽培是苹果生产栽培的必然趋势,具有优质高效、结果早、便于机械化操作等优点,利用矮化砧木是实现矮密栽培的主要手段,但不同矮化砧木对嫁接品种树体生长和叶片光合特性的影响存在显著差异[1-9]。选择适宜的矮化砧木是矮砧密植栽培成功的前提。目前,我国矮化砧木的应用以国外引入的M系(M26、M9、T337)为主,应用面积占矮化苹果总面积的70%;国内选育的SH系占15%;GM256占5%;其他砧木占10%。由于国外引进砧木适应性存在风险,不能满足我国不同产区的多样化需求,系统评价国内自主选育的砧木品系对嫁接品种生长习性、生理特性的影响,以筛选适应我国不同产区生产的砧穗组合,对实现苹果产业结构调整、保障我国苹果产业的健康可持续发展具有重要意义[10-14]。SH1和Y-1为山西省农业科学院果树研究所选育的本土砧木品种,其中,SH1苹果矮化砧木砧穗亲和性好、抗逆性强、综合性状优良[15],目前已在山西、山东和陕西等13个省市自治区广泛栽植,种植面积达10万hm2,已形成一套高效栽培管理模式,是当前黄土高原苹果产区的主推砧木;Y-1苹果矮化砧木是Y系砧木中综合性状突出的单系之一,2013年通过山西省农作物品种审定委员会审定,作为中间砧嫁接品种具有矮化性状突出、早花效果明显、抗逆性强等特点[12],应用前景广阔。本试验比较了SH1、Y-1矮化中间砧嫁接长富2号幼树的新梢生长、枝类组成及叶片光合特性,分析了2种矮化砧木对富士树体生长和光合指标的影响,旨在为2种矮化砧木在生产上的利用和选择提供理论依据。
1 材料和方法
1.1 试验材料
供试材料为矮化中间砧长富2号(Malus.domesticaBorkh.Fuji Nagafu No.2)苹果四年生幼树,基砧均为八棱海棠(Malus×robusta(Carrire)Rehder),中间砧分别为SH1、Y-1矮化中间砧。
1.2 试验方法
试验在山西农业大学果树研究所苹果矮砧示范园(112°20′E、37°15′N)进行,海拔为820 m,果园株行距为1.2 m×4.0 m,行内生草、树下起垄覆膜,全部采用细长纺锤形管理。试验采用随机区组设计,选取正常生长的四年生SH1和Y-1矮化中间砧长富2号苹果树各5株为试材,单株为1小区,挂牌标记。
1.3 测定指标及方法
1.3.1 树体生长量测定 于2017年5月28日开始,树体外围随机选取5个一年生枝条挂牌,每25 d测定新梢长度(卷尺)和粗度(数显游标卡尺)1次,共测定6次,所得数据取平均值,秋季落叶后统计枝条数量,并计算枝类组成比例。
1.3.2 光合指标测定 随机选取植株东部外围、地上1.2 m新梢无缺损、无病虫危害的基部第5~7片叶挂牌标记,于7月1日和8月28日分别测定标记叶片的光合生理指标,主要包括:净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)和胞间CO2浓度(Ci),初始荧光(Fo)、最大荧光(Fm)、可变荧光(Fv=Fm-Fo)和PS Ⅱ最大光化学量子产量 (Fv/Fm)、PSⅡ实际量子产量(ΦPSⅡ)、电子传递速率(ETR)、光化学猝灭(qP)和非光化学猝灭(NPQ),光响应参数光饱和点(LSP)、光补偿点(LCP)和最大光合速率(Pnmax)的拟合参考宋哲、叶子飘等[16-18]的方法进行;于5月28日开始,每25 d测定1次净光合速率Pn(叶室内光照强度设定1 000 μmol/(m2·s)),共测定6次,如遇阴雨天气,向后顺延,另采集无病害、完整叶片30片带回实验室测定叶绿素含量,具体参考高俊凤、薛思嘉等[19-20]的方法进行。
1.4 数据分析
采用Excel 2010和SAS 7.8软件进行数据整理、制图及差异显著性分析。
2 结果与分析
2.1 SH1和Y-1中间砧对富士幼树生长的影响
调查结果表明,SH1和Y-1中间砧富士均为7月上旬春梢停长,7月下旬抽生秋梢,周年生长趋势基本一致;SH1新梢长度、新梢粗度增长速度高于Y-1。由表1可知,SH1新梢粗度除5月28日表现低于Y-1外,7月1日之后始终比Y-1高,7月31日之后二者之间差异不显著;而全年新梢长度SH1分别较Y-1高36.38%(2017)和40.38%(2018),表明SH1中间砧富士幼树树势强于Y-1。
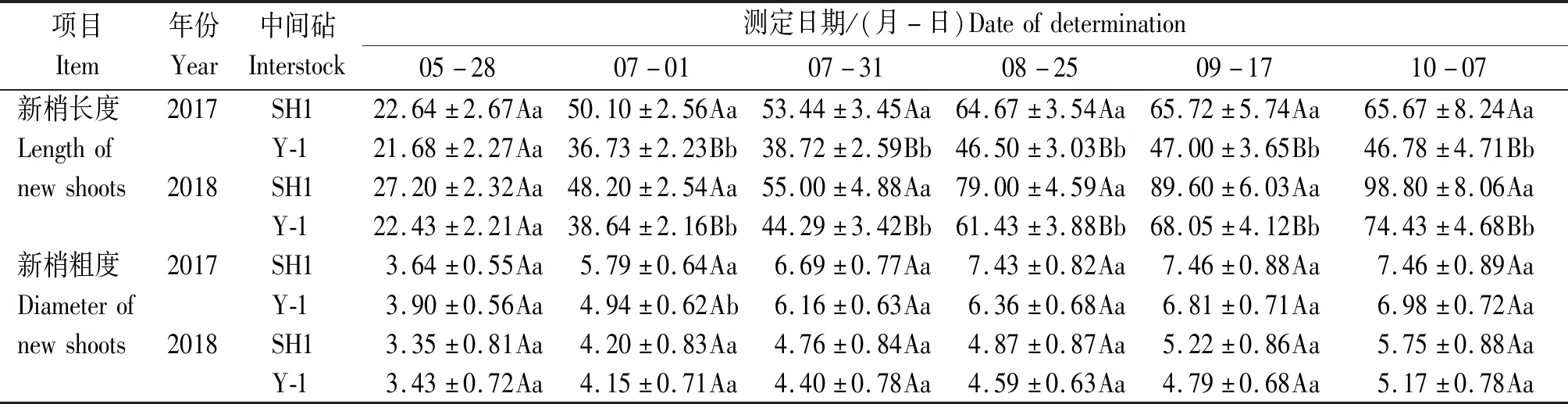
表1 SH1和Y-1中间砧富士幼树新梢周年生长比较Tab.1 Comparison of SH1 and Y-1 interstocks on annual growth in Fuji new shoots cm
由表2可知,2个中间砧富士幼树的新梢生长差异显著,Y-1春秋梢长度Y-1均显著低于SH1;节间长度(春梢)Y-1与SH1差异不显著,而节间长度(秋梢)Y-1极显著低于SH1;Y-1枝条上部/基部值高于SH1,表明Y-1控制秋梢生长能力强于SH1。
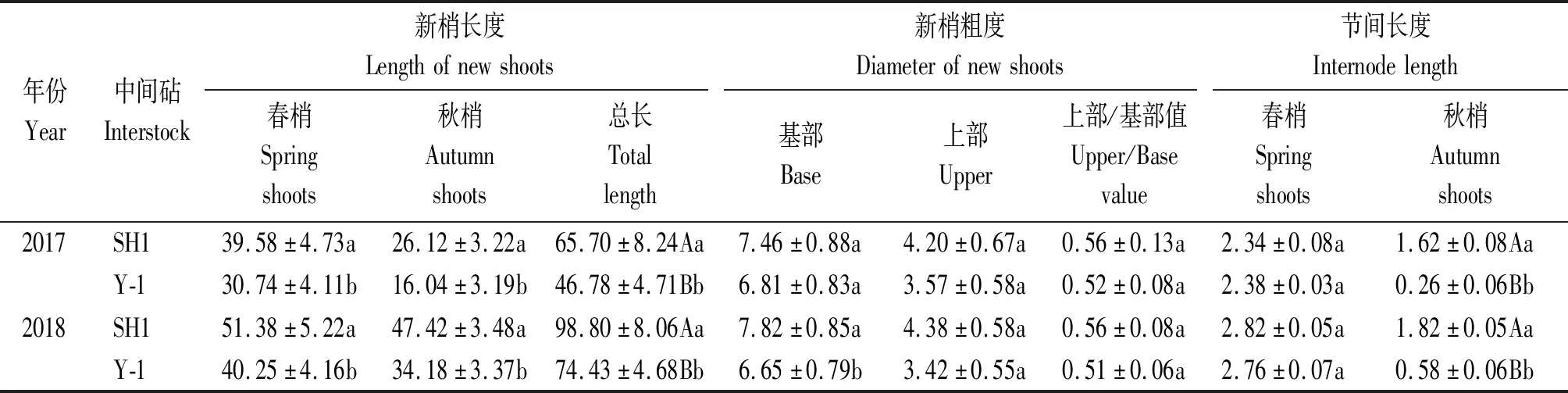
表2 SH1和Y-1中间砧对富士幼树新梢生长的影响Tab.2 Effect of SH1 and Y-1 interstocks on new shoots growth in Fuji cm
由表3可知,Y-1中间砧富士幼树长枝占比显著低于SH1,2017, 2018年分别为SH1的54.66%,74.04%;中短枝比例较高,2017,2018年分别为SH1的89.50%,84.14%,与SH1差异不显著;而短枝比率Y-1显著高于SH1,表明Y-1中间砧富士易成型性较SH1高。

表3 不同中间砧对富士枝类组成的影响Tab.3 Effect of different interstocks on branch composition of Fuji %
SH1和Y-1中间砧对果实的外观和内在品质影响存在差异,由表4 可知,与SH1相比,Y-1中间砧富士着色指数、果实硬度、可溶性固形物含量和固酸比均有显著提高,表明Y-1中间砧提高富士果实品质的效果优于SH1。
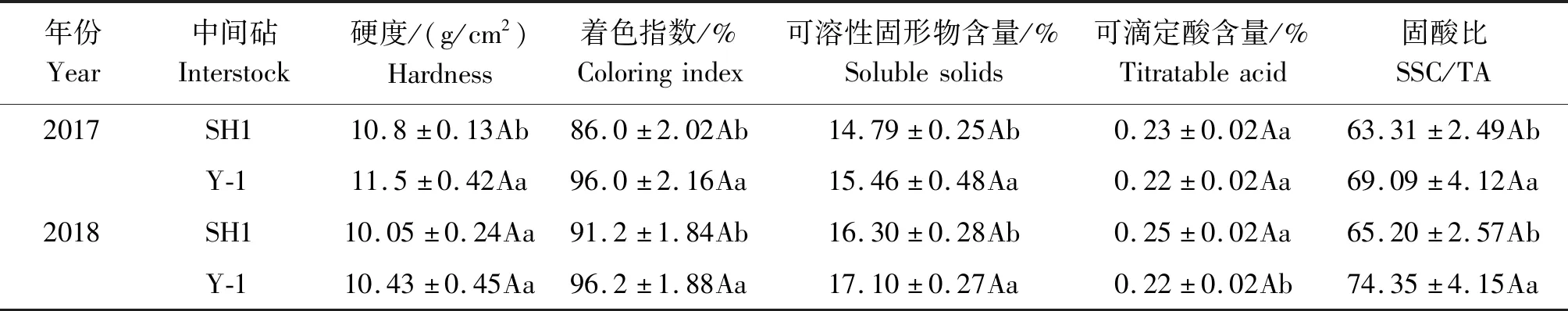
表4 不同中间砧对富士果实品质的影响Tab.4 Effect of different interstocks on Fuji fruit quality
2.2 SH1和Y-1中间砧对富士苹果叶片光合指标的影响
SH1和Y-1中间砧富士苹果叶片光合速率周年变化差异明显,其中,SH1 Pn周年变化为单峰曲线,Y-1为双峰曲线;生长前期SH1叶片Pn显著高于Y-1,生长后期SH1叶片Pn迅速降低,Y-1叶片Pn在9-10月较为稳定,出现第2次高峰,分别较SH1高27.21%,33.18%,且差异达极显著水平(表5),表明树体生长后期Y-1中间砧富士叶片光合速率能维持在较高的水平,更有利于积累营养物质,促进花芽的形成,这可能是生产中Y-1中间砧长富2号容易形成花芽的原因之一。

表5 不同中间砧富士叶片Pn周年变化Tab.5 The annual changes of Pn in Fuji leaves with different interstocks
不同时期SH1和Y-1中间砧富士苹果叶片Pn日变化也存在明显差异。由表6可知,生长前期SH1中间砧富士叶片Pn仅在9:20出现峰值,而Y-1分别于9:20和13:20出现峰值,且在9:20以后SH1组合的Pn均高于Y-1,11:20时差异达极显著水平,表明生长前期Y-1中间砧富士叶片容易受到强光抑制,而同时期SH1中间砧富士叶片对强光的利用优于Y-1。生长后期SH1和Y-1中间砧富士叶片Pn均在9:20出现峰值,Y-1中间砧富士叶片Pn比SH1组合高49.37%,两者差异极显著,表明生长后期Y-1中间砧富士叶片光合能力强于SH1。
由图1可知,SH1和Y-1中间砧富士叶片的光响应参数在不同生长阶段有较大差异,生长后期SH1中间砧富士叶片光响应参数较生长前期明显下降,而Y-1中间砧富士叶片光响应参数较生长前期大部分时段均升高,且相同时期组合之间也存在显著差异,生长前期SH1中间砧富士苹果叶片LSP、Pnmax显著高于Y-1,LCP显著低于Y-1,生长后期Y-1的LSP、LCP、Pnmax均大于SH1,且LCP较SH1高117%,且差异达极显著水平,说明生长前期SH1中间砧富士叶片对光环境的适应性强,光合能力强于Y-1;生长后期Y-1中间砧富士叶片光合能力强于SH1,但利用弱光的能力较SH1差。
由图2可知,SH1和Y-1矮化中间砧富士苹果叶片的叶绿素荧光参数间存在显著差异,同一矮化砧不同生长时期叶绿素荧光参数差异也明显。其中,生长前期,SH1矮化中间砧富士苹果叶片Fv/Fm、ΦPSⅡ、ETR极显著高于Y-1,NPQ较Y-1高107.50%,表明同时期SH1中间砧富士叶片吸收光能用于热耗散的比例低于Y-1,光合能力显著高于Y-1;生长后期SH1矮化中间砧富士苹果叶片Fv/Fm、ΦPSⅡ、ETR、qP极显著降低,NPQ较前期增加了79.52%,Y-1组合叶片Fv/Fm极显著升高,ΦPSⅡ有所升高但差异不显著、ETR、qP极显著降低,NPQ较前期降低了14.19%,表明生长后期,Y-1中间砧富士叶片光能利用和实际光能转换效率显著高于SH1。
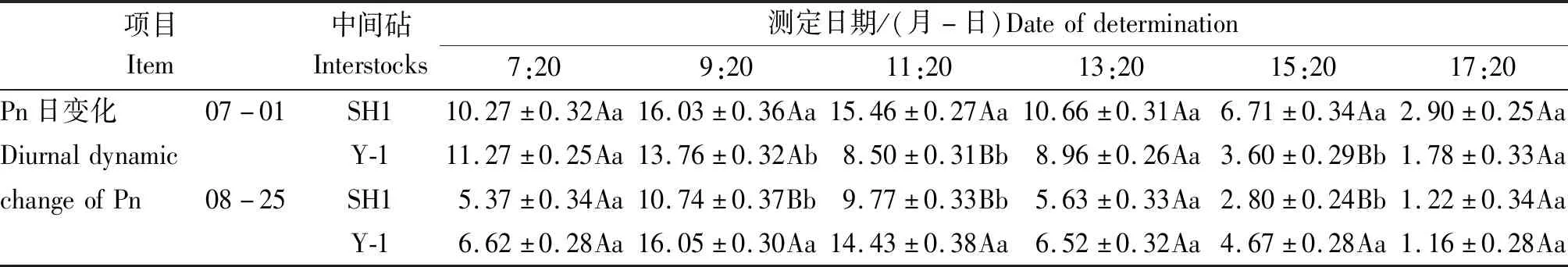
表6 不同中间砧对富士叶片Pn周年变化和日变化比较Tab.6 The daily changes of Pn in Fuji leaves with different interstocks
由图3可知,SH1和Y-1中间砧富士苹果叶片的各种光合色素含量均呈先增加后降低的变化趋势,但其峰值出现时间不同,SH1组合叶片光合色素含量峰值出现在7月1日,Y-1则出现于7月31日,且峰值之后Y-1组合叶片光合色素含量均高于SH1,类胡萝卜素含量在7-9月Y-1显著高于SH1,表明Y-1中间砧富士苹果叶片成熟滞后于SH1,叶绿素含量下降的速度低于SH1,Y-1组合具有较强的抵御逆境能力。
3 结论与讨论
苹果嫁接树的各种生理生化特性受到中间砧、基砧及嫁接品种特性与生长环境等多种因素的影响,不同中间砧对嫁接树体生长、枝类组成的影响差异较大[8-14]。赵同生等[1]对9种矮化中间砧宫崎短枝富士的研究表明,不同矮化中间砧组合的树体生长势、枝类组成、早花早果性和果实品质有明显差异。李海燕[8]、里程辉[9]的研究也得出相似结论。本试验中,Y-1中间砧富士新梢生长尤其是秋梢生长极显著低于SH1,短枝比例显著高于SH1,果实硬度、可溶性固形物含量、着色指数、固酸比均高于或显著高于SH1,表现出更好的易成型性和易早花性,表明随着中间砧矮化程度的提高,能有效限制嫁接品种新梢的生长,促进中短果枝的形成,果实品质提高效果明显,这与杨廷桢等[12]的研究结果一致。
光合作用是植物生产力构成的主要因素,受品种、光、温、水分等多种生理生态因子的影响[21-36]。李海燕[8]对6中矮化中间砧华红苹果的光合特性进行比较,认为不同矮化中间砧组合的光饱和点、光补偿点、强弱光的适应能力及光合潜力存在差异;里程辉[9]系统研究了3种矮化中间砧对岳冠成龄树光合生理特性的影响,结果表明,不同矮化中间砧能够影响嫁接品种的Pn,随着砧木的矮化性增加 Pn 值降低。本试验中,SH1、Y-1中间砧富士叶片Pn的日变化和年动态变化均呈现先升后降的趋势,但其光响应参数和叶绿素含量、叶绿素荧光参数存在显著差异。生长前期SH1中间砧富士叶片光合色素峰值出现明显早于Y-1,同等栽植密度下叶片成熟度高,对光环境的利用范围较广,Fv/Fm极显著高于Y-1,SH1总体光合效能高于Y-1,营养的快速积累促进植株生长,使SH1组合新梢生长显著高于Y-1;生长后期随着Y-1叶片逐渐成熟,光合色素积累高于SH1,Fv/Fm显著高于SH1,叶片衰老延后,总体光合效能Y-1显著高于SH1,而且Y-1能够有效控制秋梢生长、促进树体从营养生长向生殖生长的转变,有利于养分的积累和花芽的形成。可见,不同中间砧富士叶片光合特性与植株生长具有一定的相关性,其根本与中间砧的调控途径有关,具体作用机制有待进一步研究。
根据砧穗组合光合特性采用合理的栽植密度和配套管理措施,能够有效提高果品质量和产量[37]。本研究结果表明,SH1、Y-1作中间砧嫁接长富2号,其生长量和光合特性存在差异,SH1中间砧富士前期生长快速,叶片成熟早,对光的利用范围较Y-1广,Y-1中间砧能够在生长后期有效控制长富2号新梢生长、提升叶片光合作用效率、有利于营养物质的积累和花芽的形成,促进果实发育和品质的提升,生产中应考虑2个中间砧对富士生长和光合特性的差异,制定合理的栽植密度,为果园丰产稳产打好基础。
猜你喜欢
北华大学学报(自然科学版)(2022年5期)2022-11-21
今日农业(2021年19期)2022-01-12
今日农业(2021年17期)2021-11-26
鸭绿江·华夏诗歌(2021年6期)2021-09-13
今日农业(2021年7期)2021-07-28
落叶果树(2021年6期)2021-02-12
农家科技下旬刊(2017年5期)2017-06-20
山西果树(2017年1期)2017-03-02
环球时报(2014-09-04)2014-09-04
北京文学(2007年5期)2007-05-23
