
烟草西柏三烯二醇含量的遗传分析
2021-03-24 11:03鞠馥竹张洪博闫宁杜咏梅张忠锋付秋娟柳林青刘静刘艳华
中国烟草科学 2021年1期
关键词:烟草
鞠馥竹 张洪博 闫宁 杜咏梅 张忠锋 付秋娟 柳林青 刘静 刘艳华

摘 要:烟草西柏三烯二醇(CBT-diols)不仅是烟叶中香气成分的重要前体物质,还具有抗癌、抗菌等生物活性,为揭示CBT-diols含量的遗传模型及遗传效应,本研究以高CBT-diols含量种质织金黑吊把和低CBT-diols含量种质Mont Clame Brun为亲本配制杂交组合,获得P1、P2、F1、F2的四世代遗传分析群体,测定F2群体单株CBT-diols含量,利用“主基因+多基因”混合遗传模型对CBT-diols含量相关基因进行遗传分析。结果表明,α-CBT-diol、β-CBT-diol与CBT-diols含量3个性状均受2对加性-显性-上位性主基因+加性-显性-上位性多基因(E-0)控制,2对主基因的加性效应均为正值,显性效应值均为负值,且第1对主基因的加性效应及显性效应均高于第2对。CBT-diols含量3个性状的主基因遗传率分别为76.96%、76.88%、77.55%,多基因遗传率不到1%。因此,CBT-diols含量主要受2对主基因遗传效应的影响。
关键词:烟草;西柏三烯二醇;主基因+多基因;遗传分析
Abstract: Cembratrien-diols (CBT-diols) in tobacco is not only an important precursor substance of aroma components in tobacco leaves, but also has anti-cancer, antibacterial and other medicinal values. To reveal the genetic model and genetic effect of CBT-diols content, an F2 population was developed by crossing between the high CBT-diols content germplasm Zhijinheidiaoba (maternal parent) and low CBT-diols content germplasm Mont Clame Brun (paternal parent). Genetic analysis was carried out by using joint segregation analysis method with a mixed genetic model of major gene plus polygene.Results from genetic analysis showed that α-CBT-diol, β-CBT-diol and CBT-diols contents were controlled by 2 pairs of additive-dominant-epigenetic major genes plus additive-dominant-epigenetic polygenes (E-0). The additive effects of the two major genes were all positive, and the dominant effects were negative, and the additive and dominant effects of the first pair of major genes were stronger than those of the second pair. The heritability of the major genes of the 3 traits of CBT-diols content was 76.96%, 76.88%, and 77.55%, the polygenic inheritance rate was less than 1%. In general, the genetic effect inheritance of the 2 pairs of main genes was dominant in the three traits. Therefore, the content of CBT-diols was mainly affected by genetic effect, and the genetic effects of the major genes were the main genetic effects.
Keywords: tobacco; cembratrien-diols; major gene plus polygenes; genetic analysis
西柏三烯二醇(Cembratrien-diols, CBT-diols)是烟草腺毛主要分泌物,占腺毛总分泌物的60%左右[1],属于萜类化合物中的西柏烷型二萜,并且存在两种同分异构体α-CBT-diol和β-CBT-diol。CBT-diols在烟叶调制和陈化过程中可降解为茄酮、茄呢呋喃、降茄二酮等成分,是烟叶香气物质的重要前体物,另外,CBT-diols还具有抗肿瘤、抗菌及延缓衰老等生物活性,具有重要的医药价值。
CBT-diols含量在烟草不同類型种质、不同器官以及不同叶位间均存在显著差异,付秋娟等[2]对不同种质CBT-diols含量进行了检测,结果表明,供试种质烟花CBT-diols含量范围为0.08%~3.47%,其中晒烟>香料烟>烤烟>雪茄>白肋烟;刘翠翠等[3]利用UPLC方法对不同类型烟草种质CBT-diols含量进行鉴定结果表明,烤烟>晒烟>香料烟>雪茄>白肋烟,二者研究结果均表明不同烟草类型种质CBT-diols含量差异显著,且雪茄烟和白肋烟CBT-diols含量最低。另外,NIELSEN等[4]、鞠馥竹等[5]研究表明,CBT-diols含量的遗传受加性效应和显性效应的影响,不同种质间其遗传效应不同,且烟草CBT-diols含量是受多基因控制的,但控制该性状的多基因是否存在主效基因,主效基因间是否存在互作等研究未见报道。本研究利用高、低CBT-diols含量的烟草种质配制杂交组合,构建F2代遗传群体,对P1、P2、F1、F2四世代CBT-diols含量进行检测,并利用主基因+多基因混合遗传模型对四世代进行遗传分析。以确定CBT-diols含量的遗传模型,揭示其遗传机制,为通过遗传改良提高CBT-diols含量提供理论依据。
1 材料与方法
1.1 供试材料
供试材料为CBT-diols含量差异显著的织金黑吊把(P1)和Mont Clame Brun(P2)烟草品种。田间试验于2018—2019年在山东潍坊烟草有限公司诸城分公司试验站进行,2018年7月以织金黑吊把为母本、Mont Clame Brun为父本配制杂交组合,获得F1,2018年10月,将F1于温室种植套袋自交,获得F2代种子;2019年将P1、P2、F1和F2全部材料进行田间种植。亲本(P1、P2)种植2行区共50株,F1种植2行区共50株,F2种植10行区共250株。株行距120 cm×50 cm,田间管理按当地统一模式进行。
1.2 样品制备
在团棵期(移栽后30 d)对P1、P2、F1、F2全部群体单株进行取样,每株取第3叶位的叶片,并用锡箔纸包好,带回实验室放入冷冻冰柜(?20 ℃)进行贮存。冷冻干燥前,先用液氮浸泡10 min,之后放入冷冻干燥机进行冷冻干燥,72 h后取出磨样。
1.3 CBT-diols含量测定
采用付秋娟等[6]的方法对四世代材料进行α-CBT-diol和β-CBT-diol含量测定。CBT-diols含量为α-CBT-diol含量和β-CBT-diol含量之和。
1.4 数据分析
利用SPSS 25绘制F2群体频率分布直方图。采用植物数量性状主基因+多基因混合遗传模型多世代联合分析方法,对P1、P2、F1和F2群体的CBT-diols含量进行遗传效应分析。由模型的极大似然值(Maxlike Lihood Values, MLV)计算出AIC(Akaikes information criterion)值,根据最小AIC值原则,选择AIC值最小或相对较小的几组模型为备选模型,同时进行一组样本分布与模型所代表的理论分布间的适合性检验,包括均匀性检验(U12、U22、U32)、Smirnov检验(nW2)和Kolmogorov检验(Dn),根据适合性检验结果在备选模型中选出最佳遗传模型,同时选得相应的各成分分布参数。利用SEA软件中最小二乘法原理计算最佳模型中的一阶遗传参数和二阶遗传参数[7-10]。
2 结 果
2.1 CBT-diols含量表型数据分析
对四世代CBT-diols含量进行检测后结果显示(表1),2个亲本(P1、P2)的α-CBT-diol含量平均值分别为0.278 0%、0.041 3%,β-CBT-diol含量平均值分别为0.190 9%、0.030 3%,总CBT-diols含量平均值分別为0.468 9%、0.071 6%,两亲本之间CBT-diols含量差异较大。F1的α-CBT-diol、β-CBT-diol及CBT-diols含量平均值分别为0.056 6%、 0.043 5%、0.100 0%,介于两亲本之间,并略高于低值亲本。F2群体的α-CBT-diol、β-CBT-diol及CBT-diols含量平均值分别为0.138 7%、0.099 2%、0.237 9%,虽明显高于P2各性状平均值,但未达到中亲值。变异系数揭示了群体的数量性状之间的分离情况,分离群体F2的3个性状的变异系数分别为65.19%、62.52%、64.05%,均高于P1、P2及F1三世代的变异系数,并且F2群体在3个性状中均有超亲个体出现,说明分离世代具有更丰富的遗传变异。3个性状在四世代中偏度系数在?0.018 3~0.955 7之间,峰度系数在?1.329 3~0.827 4之间,从图1中也可以看出,F2群体的3个性状的含量均呈正偏态分布,推测α-CBT-diol、β-CBT-diol及CBT-diols含量可能均是主基因和多基因共同控制的数量性状,可利用主基因和多基因混合遗传模型进行进一步分析。
2.2 CBT-diols含量主基因+多基因混合遗传分析
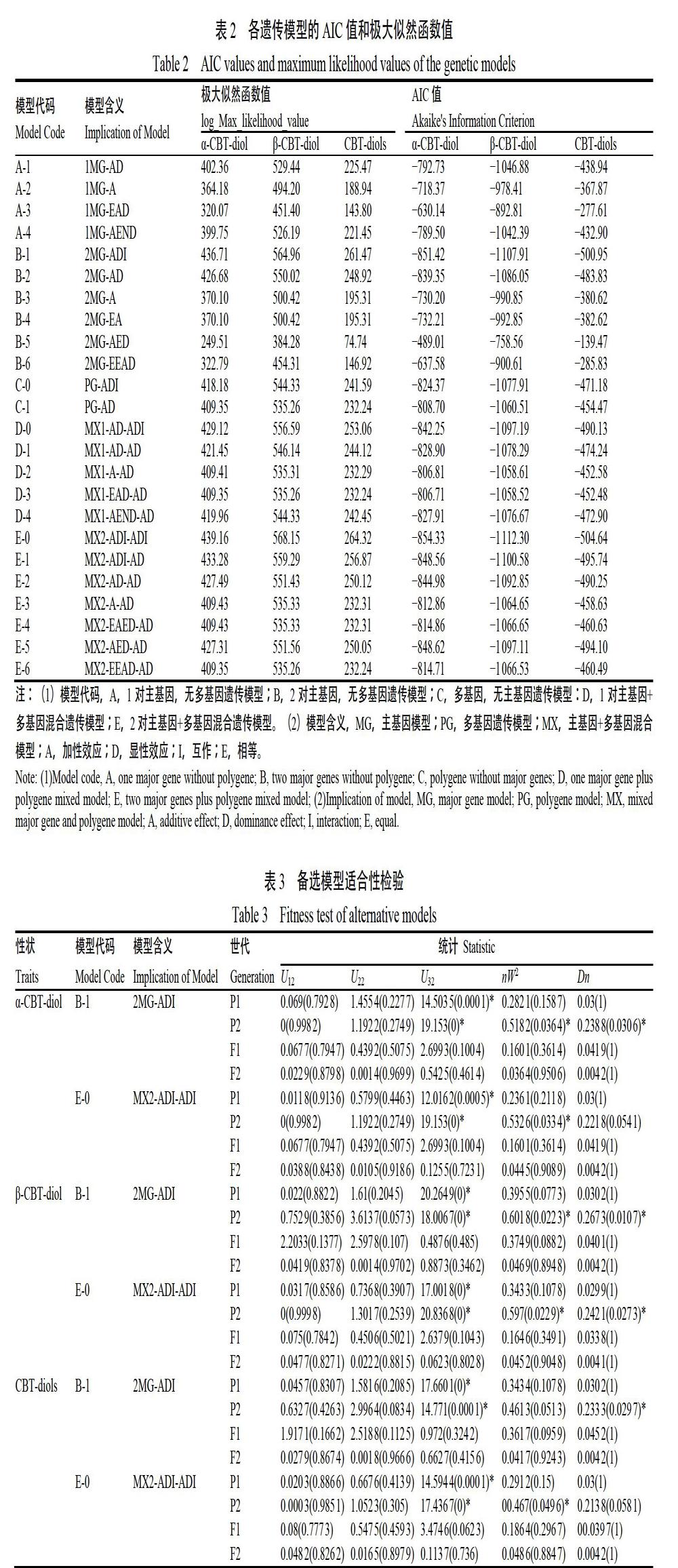
2.2.1 遗传模型的选择 利用植物数量性状主基因+多基因混合遗传模型中的多世代联合分析方法进行分析,获得5类共24种模型的MLV和AIC值(表2)。根据AIC值最小准则,选取AIC值最小的一组和接近最小的一组遗传模型作为备选模型。结果表明在这24种模型中,α-CBT-diol的B-1(2MG-ADI)及E-0(MX2-ADI-ADI)模型的AIC值最低,分别为?851.42、?854.33。β-CBT-diol的B-1(2MG-ADI)及E-0(MX2-ADI-ADI)模型的AIC值最低,分别为?1 107.91、?1 112.30。CBT-diols的B-1(2MG-ADI)及E-0(MX2-ADI-ADI)模型的AIC值最低,分别为?500.95、?504.64,可作为备选模型。从上述结果可以看出,3个性状中均可选择B-1(2MG-ADI)及E-0(MX2-ADI-ADI)为备选模型。
2.2.2 备选模型的适合性检测与最优遗传模型估算 对CBT-diols含量性状的备选模型进行均匀性检验和Kolmogorov-Smirnov检验,分别获得其统计量U12、U22、U32、nW2和Dn及其p值(表3),选择备选模型中达到显著水平的统计量个数最少且AIC值最小的模型为最优模型。结果显示,α-CBT-diol的E-0(MX2-ADI-ADI)模型中达到显著差异(p<0.05)的统计量有3个,B-1(2MG-ADI)模型达到显著差异的统计量有4个,E-0的AIC值最小;β-CBT-diol的E-0模型中达到显著差异的统计量有4个,B-1模型达到显著差异的统计量有4个,E-0的AIC值最小;CBT-diols的E-0模型中达到显著差异的统计量有3个,B-1模型达到显著差异的统计量有3个,但E-0的AIC值最小。由上述结果可知,CBT-diols含量相关的3个性状符合E-0(MX2-ADI-ADI)模型,即2对加性-显性-上位性主基因+加性-显性-上位性多基因遗传模型。
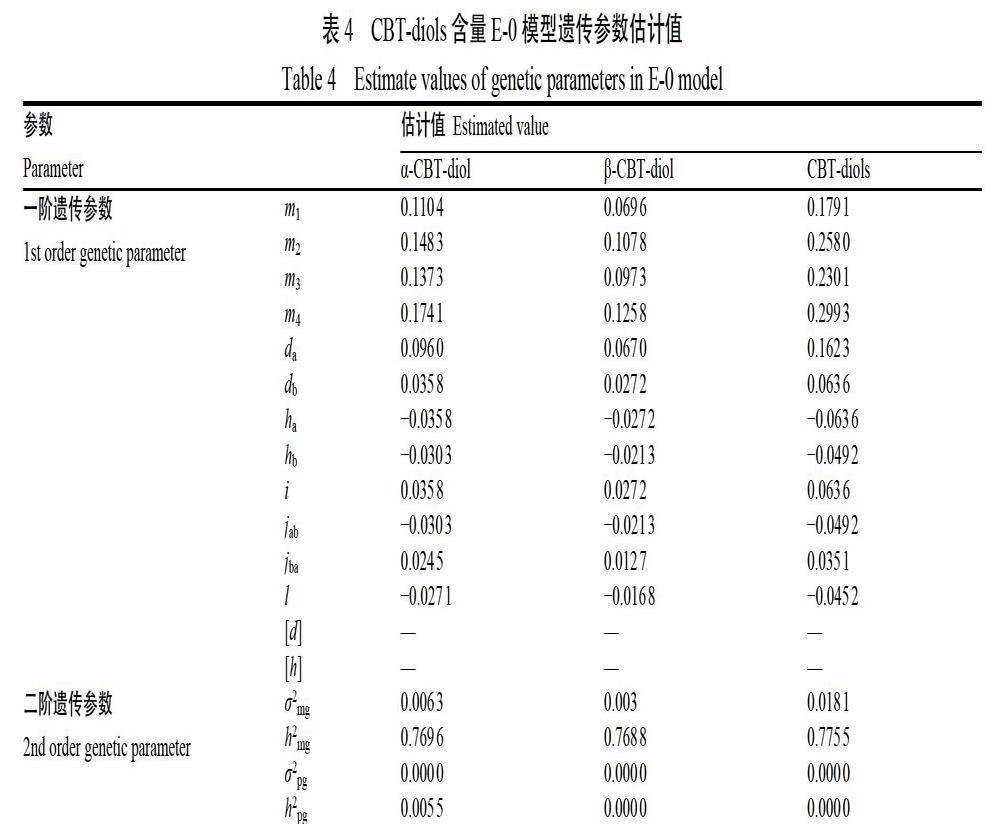
2.2.3 遗传参数估计 根据选择的最优模型(E-0)估算CBT-diols含量性状的一阶遗传参数和二阶遗传参数(表4),控制α-CBT-diol、β-CBT-diol及CBT-diols含量的第1对主基因的加性效应(da)分别为0.096、0.067、0.162 3,均为正向遗传效应,第2对主基因的加性效应(db)分别为0.035 8、0.027 2、0.063 6,同样也为正向遗传效应,说明第1对主基因加性效应大于第2对,并且2对主效基因进行累加可提高CBT-diols的含量;3个性状的第1对主基因的显性效应(ha)分别为?0.035 8、?0.027 2、?0.063 6,第2对主基因的显性效应(hb)分别为?0.030 3、?0.021 3、?0.049 2,2对主基因的显性效应均为负向效应,且第1对主基因显性效应略强于第2对主基因。3个性状第1、2对主基因显性度(h/d)均在?1~0之间,呈负向部分显性。3 个性状的2对主基因加性×加性上位性效应(i)分别为0.035 8、0.027 2、0.063 6,数值与第2对主基因的加性效应(db)相同,均为正向效应。2对主基因加性×显性上位性效应(jab)分别为?0.030 3、?0.021 3、?0.049 2,为负向效应;2对主基因显性×加性上位性效应(jba)分别为0.024 5、0.012 7、0.035 1,为正向遗传效应;2对主基因显性×显性上位性效应(l)分别为?0.027 1、?0.016 8、?0.045 2,为负向遗传效应。α-CBT-diol、β-CBT-diol及CBT-diols含量的主基因遗传率分别为76.96%、76.88%、77.55%,主基因加性效应分别为0.131 8、0.094 2、0.225 9,显性效应分别为?0.066 1、?0.048 5、?0.112 8,但多基因的加性及显性效应未检测到,故3个性状的遗传以2对主基因的遗传为主。总体来说,α-CBT-diol、β-CBT-diol及CBT-diols含量的遗传效应大小分别为:CBT-diols>α-CBT-diol> β-CBT-diol,并且3个性状的第1对主基因的加性、显性及上位性效应均大于第2对主基因。
3 讨 论
CBT-diols是烟草表面腺毛分泌物的主要成分,由于CBT-diols是由烟草植株表面腺毛的腺头细胞产生,其分泌后即留在植株表面,直接暴露在外界环境中,故环境变化对其含量的影响较大。有大量研究证明,土壤、肥料、水分、海拔、温度、光照等环境因素对CBT-diols含量均产生了直接或间接的影响。鉴于此,本试验在试验设计方面,各世代材料种植的土壤、肥料、海拔、温度以及光照等环境条件均保证了一致性。另外,取样方面,选取发育5~7 d、未经雨水的新叶(10~15 cm),保证了单株取样的一致性,提高了研究结果的可信度。烟草中CBT-diols含量是由多基因控制的数量性状[11],每个基因都对性状的表型产生或多或少的影响,性状没有明显的表型分组趋势。
传统数量遗传学一般只能将控制一个数量性状的所有基因作为一个整体进行研究,并不能得到具体到某个基因的遗传信息。而本研究利用主基因+多基因混合模型的方法研究烟草CBT-diols含量的遗传模式,可将多基因分为主效基因和微效基因,并能明确各基因的遗传参数,为下一步通过高通量测序开发SNP、QTL初步定位及精确定位,阐明各性状的多基因遗传调控网络及其互作调控关系以及分子设计育种提供重要理论依据。
四世代CBT-diols含量统计结果表明,P1、P2、F1及F2的CBT-diols含量平均值分别为0.468 9%、0.071 6%、0.100 0%、0.237 9%,F1及F2含量平均值均未达到中亲值,并倾向于低值父本,可在F2分离群体中选择CBT-diols含量较高的个体进行连续多代自交,以提高获得高含量优异后代的可能性。在控制环境条件一致的条件下,利用遗传改良手段提高后代CBT-diols含量、创制高含量種质是一种高效可行的方法。对CBT-diols含量相关的3个性状遗传模型进行筛选后确定最优遗传模型均是E-0(MX2-ADI-ADI)模型即2对加性-显性-上位性主基因+加性-显性-上位性多基因遗传模型。这说明控制3个性状的2对主基因具有明显的加性-显性-上位性作用,多基因也具有加性-显性-上位性效应。α-CBT-diol、β-CBT-diol及CBT-diols含量3个性状的主基因遗传率依次为76.96%、76.88%、77.55%,而α-CBT-diol多基因遗传率仅为0.55%,其余两个性状的多基因遗传率为0,这说明该性状在遗传上非常稳定,因此可以将3个性状的主基因遗传率视为该性状的广义遗传率。
鞠馥竹等[5]利用10个烟草品种进行了不完全双列杂交,并对其后代CBT-diols含量进行了检测,对遗传率进行估计后得到α-CBT-diol、β-CBT-diol及CBT-diols含量3个性状的广义遗传率分别为70.61%、68.02%、70.06%,与本研究中CBT-diols含量的广义遗传率相近。前人研究仅仅揭示了CBT-diols含量的遗传效应及遗传率,而本研究结果则进一步明确了3个性状主要受2对主效基因的影响,且主效基因的遗传效应大于多基因的遗传效应。因此,在CBT-diols含量育种实践中应充分考虑主效基因的影响,可以选择高CBT-diols含量亲本为轮回亲本来选育高含量优异后代。同时,下一步还需从分子水平上进行QTL定位研究,最终为CBT-diols含量分子标记辅助选择以及图位克隆提供理论基础和技术支持。
4 结 论
本研究以高CBT-diols含量种质织金黑吊把为母本、低CBT-diols含量种质Mont Clame Brun为父本构建了F2遗传群体,并以四世代群体为材料对CBT-diols含量进行主基因+多基因遗传分析,结果表明,CBT-diols含量相关性状的遗传符合2对加性-显性-上位性主基因+加性-显性-上位性多基因(E-0)遗传模型,2对主基因的加性效应均为正值,显性效应均为负值,且第1对主基因的加性效应及显性效应均强于第2对。CBT-diols含量3个性状的主基因遗传率分别为76.96%、76.88%、77.55%,多基因遗传率不到1%。总体来说,CBT-diols含量相关的3个性状以2对主基因遗传为主,主基因的遗传效应大于多基因的遗传效应。本研究结果为CBT-diols含量分子标记辅助选择奠定了基础,为高CBT-diols含量资源的筛选和高效利用提供了保障。
参考文献
[1]JOHNSON A W, SEVERSON R F, HUDSON J, et al. Tobacco leaf trichomes and their exudates[J]. Tob Sci, 1985, 29: 67-72.
[2]付秋娟,刘艳华,杜咏梅,等. 我国烟草资源西柏三烯二醇含量分析[J]. 中国烟草学报,2019,25(5):10-14.
FU Q J, LIU Y H, DU Y M, et al. Analysis of cembratriendiol contentin tobaccos growing in China[J]. Acta Tabacaria Sinica, 2019, 25(5): 10-14.
[3]刘翠翠. 烟草西柏三烯二醇检测方法及其积累动态、分布研究[D]. 北京:中国农业科学院,2016.
LIU C C. Establishment and application of a detection method for investigating the accumulation, distribution of cembratrien-diols in tobacco[D]. Beijing: Chinese Academy of Agricultural Sciences, 2016.
[4]NIELSEN M T, SEVERSON R F. Inheritance of a diterpene constituent in tobacco trichome exudate[J]. Crop Science, 1992, 32(5): 1148-1150.
[5]鞠馥竹,张洪博,闫宁,等. 烤烟西柏三烯二醇含量遗传效应分析[J]. 分子植物育种,2020,18(8):2633-2640.
JU F Z, ZHANG H B, YAN N, et al. Analysis on genetic effects of cembratrien-diol content in flue-cured tobacco[J]. Molecular Plant Breeding, 2020, 18(8): 2633-2640
[6]付秋娟,杜咏梅,刘新民,等. 超高效液相色谱法测定烟草西柏三烯二醇[J]. 中国烟草科学,2017,38(3):67-73.
FU Q J, DU Y M, LIU X M, et al. Determination of cembratriene-diol in tobacco by ultra performance liquid chromatography[J]. Chinese Tobacco Science, 2017, 38(3): 67-73.
[7]章元明,盖钧镒. 利用P1、F1、P2、F2和F2:3家系五世代联合分离分析的拓展[J]. 生物数学学报,2002(3):363-368.
ZHANG Y M, GAI J Y. An expansion of the joint segregation analysis of P1、F1、P2、F2 and F2:3 generations in the mixed major gene plus polygenes inheritance analysis of quantitative traits[J]. Journal of Biomathematics, 2002(3): 363-368.
[8]王建康,盖钧镒. 數量性状主-多基因混合遗传的P1、P2、F1、
F2和F2:3联合分析方法[J]. 作物学报,1998(6):15-23.
WANG J K, GAI J Y. Identification of major gene and polygene mixed inheritance model of quantitative traits by using joint analysis of P1、P2、F1、F2 and F2:3 generations[J]. Acta Agronomica Sinica, 1998(6): 15-23.
[9]盖钧镒,章元明,王建康. QTL混合遗传模型扩展至2对主基因+多基因时的多世代联合分析[J]. 作物学报,2000,26(4):385-391.
GAI J Y, ZHANG Y M, WANG J K. A Joint analysis of multiple generations for QTL models extended to mixed two major genes plus polygene[J]. Acta Agronomica Sinica, 2000, 26(4): 385-391.
[10]刘兵. 植物数量性状分离分析Windows软件包SEA的研制[D]. 南京:南京农业大学,2013.
LIU B. SEA: a software package of segregation analysis of quantitative traits in plants[D]. Nanjing: Nanjing Agricultural University, 2013.
[11]王冬,孙九哲,彭玉富,等. 不同品种(系)烤烟西柏三烯二醇代谢差异研究[J]. 中国烟草学报,2014,20(6):63-69.
WANG D, SUN J Z, PENG Y F, ET AL. Metabolism differences of cembratrien-diols in different tobacco cultivars[J]. Acta Tabacaria Sinica, 2014, 20(6): 63-69.
猜你喜欢
南方农业·中旬(2021年3期)2021-10-31
环球时报(2020-12-24)2020-12-24
农民致富之友(2020年14期)2020-05-25
人物画报(2020年22期)2020-03-09
环球时报(2017-07-31)2017-07-31
中国周刊(2017年4期)2017-04-11
商界评论(2016年4期)2016-04-20
绿色中国(2015年6期)2015-07-03
新民周刊(2014年22期)2014-06-24
