
金钟连翘与东北连翘种间杂交F1代表型性状遗传分析
2021-03-22 09:21:08吴雨桐赵峥畑申建双张启翔潘会堂
植物研究 2021年3期
徐 洋 吴雨桐 赵峥畑 申建双 张启翔 潘会堂
(花卉种质创新与分子育种北京市重点实验室,国家花卉工程技术研究中心,城乡生态北京实验室,北京林业大学园林学院,北京 100083)
连翘属(Forsythia)植物是我国传统的早春观花灌木,姿态优美,花色鲜艳,在园林中大量应用。连翘属现有11 个种,除1 种原产欧洲东南部外,其余均产自亚洲东部,我国产7 种、1 变型,其中1 种系栽培种[1]。19 世纪,连翘属植物被引到西方国家后,连翘新品种开始大量出现[2]。1885 年,人们发现了连翘属的天然杂交种——金钟连翘(F.intermedia),认为它是连翘(F.suspensa)和金钟花(F.viridissima)的天然杂交后代[3]。1906年,人们利用连翘变种(F.suspensa var. sieboldii)与金钟花杂交获得著名的金钟连翘品种‘Spectabilis’(F.intermedia‘Spectabilis’),该品种在各地得到广泛栽植。随后人们发现了‘Spectabilis’的芽变品种——‘Lynwood’[4]。Flemer 以金钟连翘品种‘Spectabilis’为母本,卵叶连翘品种‘Tetra Gold’(F.ovata‘Tetra Gold’)为父本进行种间杂交,培育出花大色艳、抗寒性强的连翘新品种‘Princeton Gold’[5]。国内的连翘育种起步于20 世纪90 年代,刘玮等以白丁香和东北连翘为亲本进行杂交,使用胚培养的方法,获得了杂交后代植株[6],但未见培育出品种。Shen 等以金钟连翘品种‘Courtaneur’(F.intermedia‘Courtaneur’)为母本,朝鲜白连翘(Abeliophyllum distichum)为父本,通过远缘杂交培育出具有明显花香的连翘新品种‘春香’[7]。马帅等以连翘属9个种或品种和华北紫丁香作为亲本进行远缘杂交,获得了杂交后代[8]。目前培育连翘新品种最直接有效的手段仍然是传统杂交育种方法,主要的育种目标性状集中在抗寒性、生长习性和花部性状上。
观赏植物的表型性状是基因型和环境共同作用的结果,对生长于同一环境中的杂交群体的表型性状进行测定和分析,对开展杂交育种工作具有重要的指导意义[9]。杨彦伶等以紫薇杂交F1代群体为研究对象,对亲本和F1代植株的花径、花序长、花朵数、株高、冠幅等性状进行了统计分析,为合理选配紫薇杂交亲本和新品种选育提供了依据[10]。周利君等通过对月季杂交F1代群体的花部和叶片形态性状进行了分析,为挖掘控制表型性状的优良基因及辅助选择育种提供了帮助[11]。张飞等对菊花杂交F1代6个花部性状进行遗传分析,了解了菊花花部性状的杂种优势和遗传基础[12]。Wang 等通过对观赏凤梨杂交F1代群体的株高、冠幅、分枝数等性状进行调查分析,为这些性状的QTL 定位及辅助选择育种提供了依据[13]。表型性状多样性是遗传多样性和环境多样性的综合表现,而杂交育种依赖于对表型性状遗传分离规律的准确把握[14]。在连翘方面,Wang 等对连翘黄色叶性状的形成和性状分离情况进行了研究,为培育不受或少受环境影响的黄叶色连翘新品种提供了依据[15]。目前,有关连翘杂交后代性状遗传规律的研究仍然较少,很多性状的遗传特性尚不清楚,研究连翘主要性状的遗传规律对连翘新品种定向选育具有重要的指导意义。
本研究对金钟连翘品种‘Lynwood’与东北连翘种间杂交获得的F1代群体的花冠口直径、花裂片长度、抗寒性等12个表型性状进行测定,分析这些性状的遗传规律,并利用植物数量性状的主基因+多基因混合遗传模型[16]对各性状进行了遗传分析,以期为连翘的定向育种提供参考。
1 材料与方法
1.1 植物材料
以金钟连翘品种‘Lynwood’×东北连翘的130株杂交后代和杂交亲本作为研究对象,亲本的主要信息见表1 和图1。该杂交群体露地栽植于国家花卉工程技术研究中心小汤山基地,该基地位于北京市昌平区小汤山镇,年平均气温11.8℃,年平均日照时数2 684 h,年平均降水量550.3 mm。栽植地的温度、光照、水分、土壤等环境因子基本相同,植株生长健壮、表现稳定。
1.2 杂交
东北连翘开花比金钟连翘品种‘Lynwood’早约10 d。于2013 年3 月中旬初花期采集东北连翘花粉,置于变色硅胶中干燥,4℃冰箱保存。授粉前采用铁军等的方法[17]离体培养花粉检测花粉活力,萌发率在25%以上的花粉用来杂交授粉。在‘Lynwood’开花前一天,人工去雄,套上硫酸纸袋隔离。第二天10:00~12:00 柱头分泌粘液时进行授粉。授粉后20 d 左右,柱头开始萎蔫,子房开始膨大时,将硫酸纸袋换成网袋。10 月初果实成熟后收种子,室内阴干存放。11 月下旬,种子经100 mg·L-1赤霉素处理6 h 后,点播于装有草炭基质的穴盘中。2014 年3 月上盆,5 月移栽于大田,对获得的杂交F1代进行大田常规管理。最终获得包括130株后代的群体。
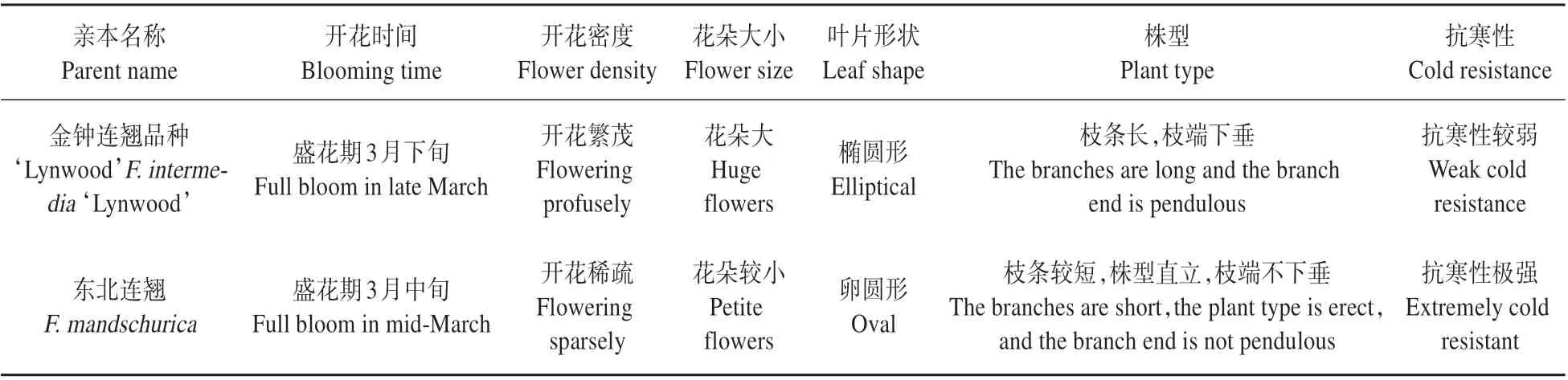
表1 金钟连翘品种‘Lynwood’和东北连翘的主要性状Table 1 Main traits of F.intermedia‘Lynwood’and F.mandschurica
1.3 表型指标和抗寒指标的测定
表型指标测定:根据国家林业局2010 年发布的《植物新品种特异性、一致性、稳定性测试指南——连翘属》,于2019 年3~4 月连翘花期,每个株系选取3朵完全开放的花,用游标卡尺测定花冠口直径(X1)、花裂片长度(X2)和宽度(X3),并计算长宽比(X4);选3 个成熟枝条,测定10cm 枝条长度内花芽的数量作为着花密度(X5)指标;8 月,选枝条从顶端向下数第5~7 节处的成熟叶片,测定叶片长度(X6)和宽度(X7),并计算叶片长宽比(X8);12 月植株落叶后,测定植株的株高(X9)和冠幅,以植株南北向和东西向的宽度作为冠幅(X10)指标,选3 个当年生枝条测定枝条长度(X11)。每个指标重复3次。
抗寒指标测定:参考Lim&Arora的方法[18],采用电解质渗出率拟合Logistic 方程,计算低温半致死温度(LT50)。植株经自然抗寒锻炼后,于2020年1 月上旬,选杂交植株上生长状况一致、粗细均匀的1年生枝条,每个株系剪取8~10个枝条,剪取的枝条长15~20 cm,用去离子水冲洗干净,擦干,横切成0.5 cm 的小段,分7 组用锡箔纸包好,置于5℃冰箱适应2 d。取出其中1份材料测定电导率作为对照,另外6份采用人工梯度降温法降温,温度梯度为-10、-15、-20、-25、-30、-35℃,降温速率为5℃/h,降至目标温度后维持12 h,取出1份材料测定电导率,其他材料继续降温。冷冻的枝条在5℃下放置2 h解冻,然后称取0.1 g装入15 mL的离心管中,加入5 mL的去离子水,摇床震荡24 h,采用DDS-307A电导仪测定初电导值(C1),将其放入沸水浴中30 min,冷却至室温后测定终电导值(C2)。每种枝条重复测定3次,取其平均值[19~20]。计算公式为:
以各个株系相对电导率值y 和相应处理温度x,拟合logistic回归方程:
式中:k 代表细胞伤害率的饱和容量,即k=100,依据方程得到参数a、b,拟合度显著时,进而求得LT50=-lna/b,作为抗寒性(X12)指标[21]。
1.4 数据统计分析
利用Excel2016、SPSS25 进行数据处理和分析,对表型性状进行描述性统计并计算变异系数,绘制频率分布图,进行相关性分析。采用盖钧镒等[16]的植物数量性状主基因+多基因混合遗传模型分析方法,利用SEA 软件统计11 种遗传模型的AIC 值(Akaike’s information criterion),选择最适合的模型,根据最小二乘法估算加性效应、显性效应、主基因遗传力等遗传参数[22]。
2 结果与分析
2.1 F1代群体表型性状的遗传变异
对F1代群体植株的花冠口直径、花裂片长度、抗寒性等12 个性状进行描述性统计(见表2)。杂交F1代个体的不同性状均有一定程度的差异,存在一定的变异幅度,12 个性状的遗传变异系数为11.33%~36.89%,变异系数均超过10%。群体性状中变异程度最大的是冠幅,变异程度最小的是花裂片长度。着花密度和冠幅的变异系数超过30%,变异较大。除花裂片长度、花裂片宽度、叶片长度外,其他性状的变异系数在15%~30%,为中等变异水平。株高的标准差最大,群体中极易出现极值个体,叶片长宽比标准差最小。
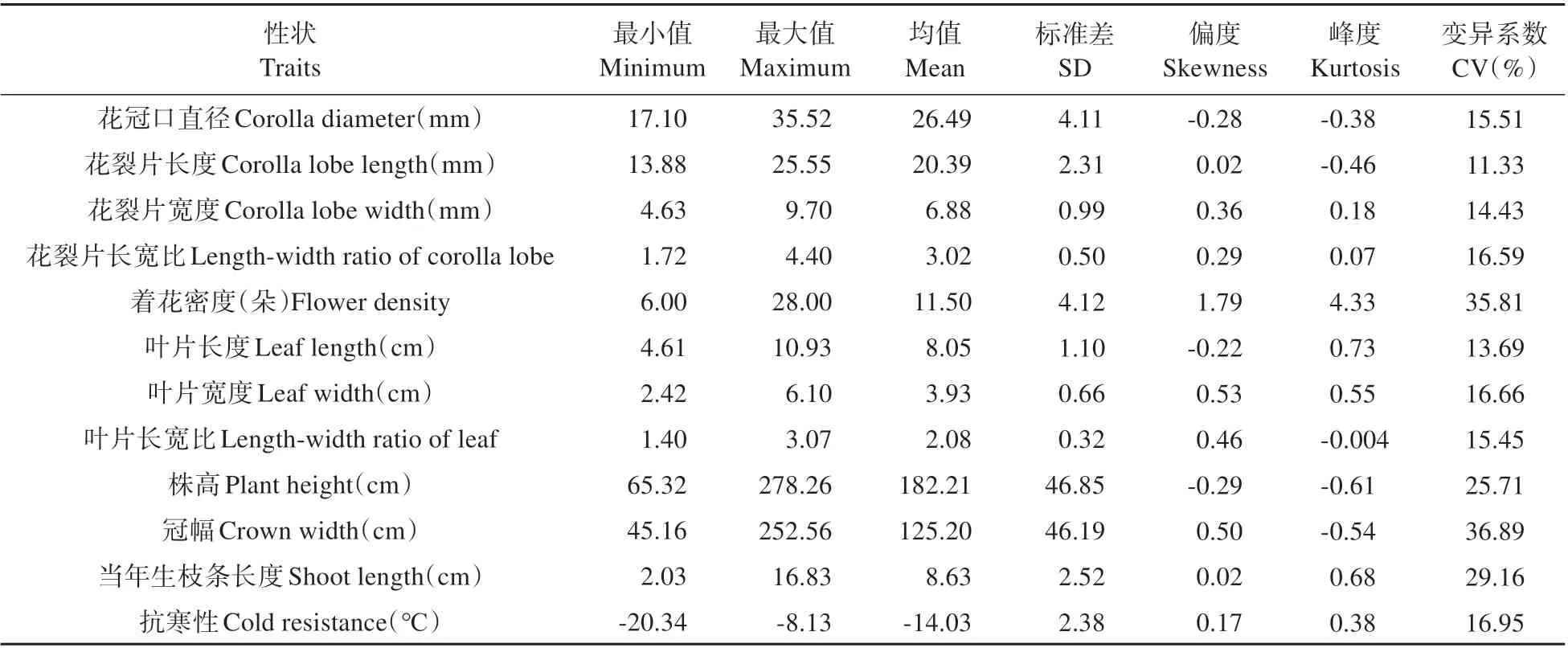
表2 金钟连翘品种‘Lynwood’×东北连翘群体表型性状的描述性统计Table 2 Descriptive statistics of phenotypic traits in F1 population of F.intermedia‘Lynwood’×F.mandschurica
对F1代群体的表型性状进行频率分布分析(见图2),结果表明,12个性状都呈现单峰分布,接近正态分布,符合数量性状的特点。除着花密度外,其他11 个性状的偏度和峰度绝对值均小于1,与正态曲线拟合较好。着花密度的偏度和峰度大于1,呈现出明显的正偏性和极端性。
对F1代群体的12个性状与亲本进行比较分析(见表3),总体上看,杂交后代分离广泛,各个性状均出现大于高亲或小于低亲的超亲个体。杂交F1代群体中花裂片长宽比的均值大于中亲值,花裂片长度、着花密度、叶片长宽比的均值与中亲值的比值接近100%,其余性状的均值小于中亲值。除花冠口直径、花裂片长宽比、叶片长度、冠幅、抗寒性以外,其他性状介于双亲之间的个体所占比值最大。F1代群体中多数个体(74.8%)的花冠口直径小于低亲,说明花冠口直径有变小的趋势。64.57%的个体花裂片长宽比大于高亲,说明花裂片形状有偏向更狭长的趋势。F1代群体没有出现冠幅大于高亲的个体,有66.15%的个体小于低亲,证明冠幅有偏向更小的趋势。抗寒性大于高亲的比例为51.11%,说明抗寒性遗传偏向于抗寒性较弱的‘Lynwood’,也有相当的比例(48.89%)介于双亲之间,能够从中筛选出抗寒性较强的植株,但群体中并未出现抗寒性大于东北连翘的植株。

表3 金钟连翘品种‘Lynwood’×东北连翘群体表型性状的遗传分析Table 3 The genetic analysis of phenotypic traits in F1 population of F.intermedia‘Lynwood’×F.mandschurica
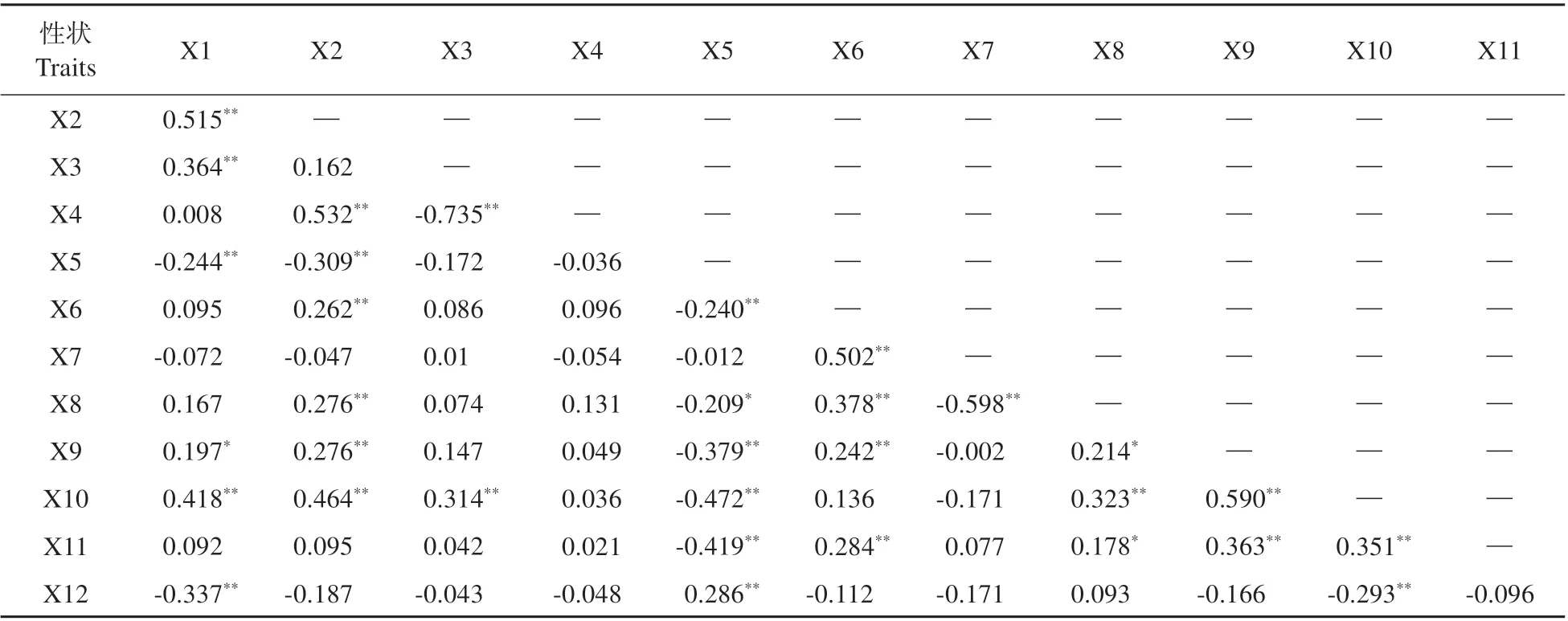
表4 金钟连翘品种‘Lynwood’×东北连翘群体表型性状的相关性分析Table 4 Correlation analysis of phenotypic traits in F1 population of F.intermedia‘Lynwood’×F.mandschurica
2.2 F1代群体表型性状间的相关性
对连翘的表型性状进行相关性分析(见表4)。花冠口直径与着花密度呈极显著负相关关系,表明着花密度越大的株系,其花冠口直径越小。着花密度与株高、冠幅、当年生枝条长度均呈现极显著负相关关系,说明着花密度与株型性状之间存在着十分紧密的相关性。当年生枝条长度与株高、冠幅呈极显著正相关关系,说明植株的长势对株高和冠幅均有显著影响。抗寒性与着花密度呈极显著正相关关系,与花冠口直径、冠幅呈极显著负相关关系。
2.3 F1代群体表型性状的遗传模型
由于连翘自交不亲和,基因型高度杂合[23]。其F1代类似于纯合亲本的F2代,常称为“假F2代”[24]。因此,可以采用植物数量性状混合遗传模型的主基因+多基因分析方法中的单个分离世代F2群体的分离分析方法来分析连翘杂交F1代群体表型性状的主基因遗传方式,11 个模型的AIC 值结果见表5。根据AIC 准则挑选AIC 值最小的一个或相对较小的几个模型,同时,基于均匀性检验(U12,,U32),Smirnov 检 验(nW2)及Kolmogorov 检 验(Dn)对候选的模型进行适合性检验。比较候选模型的统计量显著性情况,数目最少的确定为最优遗传模型。如果候选模型之间显著性数目一样,则AIC 值小的被认定为最优遗传模型[16]。以着花密度为例,AIC 值较小的模型有1MG-AD,2MGADI,2MG-AD 作为备选最适模型。然后利用适合性检验(见表5~6),结果显示3 个模型显著数目都为2,而2MG-AD 模型的AIC 值最小,所以着花密度的最适遗传模型为2MG-AD。

表5 金钟连翘品种‘Lynwood’×东北连翘群体表型性状各种遗传模型的AIC值Table 5 AIC values of various genetic models of phenotypic traits in F1 population of F.intermedia‘Lynwood’×F.mandschurica

表6 入选模型的适合性检验Table 6 Test for goodness-of-fit of selected genetic models
对入选模型进行适合性检验(见表6)。花裂片长度、花裂片宽度、花裂片长宽比、叶片长度、叶片宽度、叶片长宽比、当年生枝条长度、抗寒性的最适模型为0MG 模型,这些性状的遗传由受环境影响比较大的多基因控制。花冠口直径和冠幅的最适模型为1MG-AD 模型,这些性状由一对加性—显性主基因控制。着花密度的最适模型为2MG-AD模型,由两对加性—显性主基因控制。株高的最适模型为2MG-EAD 模型,由两对等加性—显性主基因控制[25]。
2.4 F1代群体表型性状的遗传参数
根据已选出的最适模型,采用最小二乘法估计除OMG 模型性状以外的其余性状的一阶、二阶遗传参数(见表7)。控制花冠口直径和冠幅的一对主基因的加性效应值分别为5.27 和40.48,显性效应值分别为0.77和-37.97,加性效应均大于显性效应,主基因遗传力分别为76.05%和60.3%。控制着花密度的两对主基因的加性效应值分别为4.99 和2.58,显性效应值分别为-7.02 和-0.91,说明两对主基因的加性效应均为正向增效,而显性效应均为负向效应。控制株高的主基因的等加性—显性效应值为30.71,主基因遗传力为64.75%。

表7 不同表型性状在各最适模型下的遗传参数估计值Table 7 Estimation of genetic parameters of phenotypic traits at its optimal genetic model
3 讨论
表型性状的多样性是育种工作的基础,了解和掌握杂交群体的表型性状的多样性水平和变异程度,对于新品种培育具有重要意义[26]。焦垚等通过对紫薇杂交F1代性状的统计分析,发现杂交后代在株高、冠幅上更容易产生变异[27]。Dhananjaya 等通过测定分析香石竹与中国石竹种间杂交F1代表型性状的遗传变异,发现了切花效果优良、产量高的新品种,并且掌握了遗传规律用以指导后续育种工作[28]。本研究所测定的金钟连翘与东北连翘杂交F1代群体的12 个表型性状均分离广泛,变异系数为11.33%~36.89%。根据李娟娟等对遗传变异度判定的标准[29],本研究中花裂片长度、花裂片宽度、叶片长度遗传变异度较小,遗传改良潜力一般。而花冠口直径、花裂片长宽比、着花密度、叶片宽度、叶片长宽比、株高、冠幅、当年生枝条长度、抗寒性等性状遗传变异度均达到中等水平以上,具有较大的遗传改良潜力,性状分离广泛,能够在后代群体中筛选出观赏性和抗寒性优良的连翘新品种。
通过对表型性状之间的相关性研究,可在育种中合理利用不同性状间的关系,来预测相关的性状表现,有效提高育种效率,加快育种进程。Kumar 等通过分析发现,唐菖蒲每个穗上花的数量与花朵直径呈显著正相关,开花的时长与穗长呈显著负相关关系[30]。何贵平等通过试验发现,杉木的结顶率与冻害率存在显著负相关关系,可以通过了解结顶率指标间接进行抗寒性材料的初选[31]。在本研究中植株抗寒性与花冠口直径、冠幅呈极显著的负相关,与着花密度呈极显著正相关关系。因此,在抗寒性育种过程中,可以选取比较容易测定的性状进行参考,以提高抗寒性状的育种效率。
植物大多数表型性状都是由微效多基因控制的数量性状,在同一杂交后代群体的不同个体中往往表现出连续的数量差异,易受环境影响,不易明确分组归类,需要应用统计学方法对整个群体进行测定和分析[32]。张琳等利用主基因+多基因混合遗传模型对牡丹杂交F1代20个表型性状进行分析,结果表明株高、冠幅等8 个性状由微效多基因控制,新枝长度、花朵数等12个性状受到一对或两对主基因控制[33]。Zhang 等对菊花杂交群体的初始开花时间和开花持续时间进行了混合遗传模型分析,结果发现两个性状的主基因遗传力分别为65%和72%,遗传力较大,在早期育种就可进行选择[34]。本研究发现花裂片长度、花裂片宽度、花裂片长宽比、叶片长度、叶片宽度、叶片长宽比、当年生枝条长度、抗寒性等8个性状的遗传无主基因控制,受环境影响较大的微效多基因控制;花冠口直径和冠幅由一对加性—显性主基因控制;着花密度由两对加性—显性主基因控制;株高由两对等加性—显性主基因控制。花冠口直径、冠幅、着花密度和株高的主基因遗传力分别为76.05%、60.3%、72.22%和64.75%,均大于50%,属于高度遗传力,受环境影响较小,在早期世代即可进行选择[35]。
猜你喜欢
东北林业大学学报(2023年6期)2023-05-31 02:56:16
丽水学院学报(2022年2期)2022-04-19 01:22:10
林业科技情报(2021年3期)2021-09-01 02:01:26
中国民间疗法(2021年8期)2021-07-22 05:53:08
今日农业(2020年16期)2020-09-25 03:04:42
中成药(2018年10期)2018-10-26 03:41:34
中南林业调查规划(2017年1期)2017-12-19 00:58:07
新疆农垦科技(2016年2期)2016-08-21 13:50:26
中国果菜(2016年9期)2016-03-01 01:28:40
中国马铃薯(2015年5期)2016-01-09 06:11:11
