
江西武夷山南方铁杉针阔混交林主要植物生态位特征*
2021-03-13 08:09赵家豪叶钰倩袁在翔刘江南袁荣斌关庆伟
林业科学 2021年1期
赵家豪 叶钰倩 陈 斌 袁在翔 刘江南 杨 涛 袁荣斌 关庆伟
(1.南京林业大学南京现代林业创新协同中心 南京 210037;2.南京林业大学生物与环境学院 南京 210037;3.江西武夷山国家级自然保护区 铅山 334500)
生态位是物种在群落或生境中时间和空间的位置及其在群落中的功能与地位(Soberonetal.,2005),可以体现种群在群落中的作用和重要性,探讨不同物种的生态位有助于理解自然界物种共存现象与种间竞争关系(Winemilleretal.,2015)。生态位理论中常用的量化指标主要为生态位宽度与生态位重叠度。生态位宽度是某个物种所利用各种资源总和的幅度,可以反映物种对环境资源的利用能力(陈俊华等,2010)。生态位重叠度量化了两个物种共同利用资源的程度,能反映生态位相似的物种对资源与空间的分享或竞争(Ingrametal.,2018),因此研究种群的生态位宽度和生态位重叠度对了解种群间具体关系及物种共存机制十分重要(Silvertown,2004)。经典的生态位理论认为物种间的生态位分化是物种共存的基本机制之一,对生物多样性维持机制有重要意义(Levineetal.,2009;Kraftetal.,2011),是群落构建和演替的主要动力(Tilman,2000),也会进一步影响树种组成与动态变化(Bannisteretal.,2017)。目前,单一林层优势种群的生态位特征对环境因子的响应机制(白晓航等,2017)及干扰对种群生态位特征影响(简尊吉等,2017;Baieretal.,2007;Sotoetal.,2019)的研究已取得一定进展,对干扰颇少的原始林各林层种群之间的相互关系及其对生态系统稳定性的影响鲜少研究。
南方铁杉(Tsugachinensisvar.tchekiangensis)是我国特有的第三纪孑遗植物,具有很高的科研价值。江西武夷山国家级自然保护区保存有1 560 hm2的南方铁杉天然林(袁荣斌等,2012),是南方铁杉在我国仅有的较大局域种群分布区域。以往有关南方铁杉的研究多集中在群落结构、密度制约效应(郭屹立等,2015)、种群动态(丁巧玲等,2016)、林隙干扰(何建源等,2009)及生长规律(罗金旺,2011)等方面,而研究珍稀古老物种的生态位特征,可以从资源利用能力和竞争关系的角度了解其现状及其形成因素。因此,本研究以江西武夷山核心区内的南方铁杉中亚热带针阔混交林为对象,基于保护区内海拔1 800 m的长期监测固定样地植物的调查数据,以调查的样方为资源轴,运用Levins生态位宽度、Shannon-Wiener生态位宽度和Pianka生态位重叠度,定量研究该群落中乔木层、更新层、灌木层和草本层主要植物的生态位特征,旨在明晰群落内各种群的资源利用状况及不同种群间的竞争关系,为珍稀物种南方铁杉保护及森林生态系统稳定性维持提供科学依据。
1 研究区概况
研究地点为江西武夷山国家级自然保护区,位于江西省上饶市铅山县南沿、武夷山脉北段西北坡(27°48′11″—28°00′35″E,117°39′30″—117°55′47″N),平均海拔1 200 m。保护区东南方与福建武夷山国家级自然保护区相连,构成完整的中亚热带中山森林生态系统(袁荣斌等,2012)。该地属海洋性气候和大陆性气候的过渡地带,区内地貌形态为强烈侵蚀的岩浆岩中山地貌,土壤是典型的亚热带中山土壤。年均气温14.2 ℃,月均气温1月最低(3.6 ℃),7月最高(23.8 ℃)。年均降水量1 814~3 545 mm,年均蒸发量553~941 mm,年均空气相对湿度80%~92%。
保护区主要生态系统类型为中亚热带中山山地森林生态系统,南方铁杉天然林面积达1 560 hm2(袁荣斌等,2012),其他常见植被类型有常绿落叶阔叶混交林(海拔1 300~1 800 m)、中山针阔混交林(海拔1 500~1 900 m)和山地暖性常绿针叶林(海拔1 700~1 900 m)(陈昌笃,1999)。保护区内的植物地理区系呈现由温带向亚热带过渡的现象(陈斌等,2018);植物种类十分丰富,南方铁杉、红豆杉(Taxuschinensis)、香榧(Torreyagrandis)、鹅掌楸(Liriodendronchinensis)、川黄檗(Phellodendronchinensis)等21种野生植物是保护区的重点保护对象。
2 研究方法
2.1 固定样地设置 根据史密斯桑尼亚热带研究所(Smithsonian Tropical Research Institute)的热带森林科学研究中心(The Center for Tropical Forest Science, CTFS)样地建设标准(Condit,1995),2014年于江西武夷山国家级自然保护区黄岗山西北坡设置1块中亚热带南方铁杉针阔混交林动态监测固定样地,中心点坐标为117°45′43″ E,27°50′26″ N,固定样地最低和最高海拔分别为1 758和1 843 m,平均海拔1 800 m。固定样地面积为6.4 hm2(400 m×160 m),运用全站仪将其分成160个20 m×20 m样方(陈斌等,2018)。
2015年8月,调查固定样地中的植物资源。在每个20 m×20 m样方内对胸径(DBH)≥1 cm的木本植物进行物种鉴别、坐标定位、胸径测量和编号挂牌,记录每个调查对象的胸径、树高和冠幅。在每个20 m×20 m样方中,随机设置16个1 m×1 m的小样方,记录每个小样方内草本种名及盖度。
2.2 植物重要值计算 林层划分为乔木层、更新层、灌木层和草本层,DBH≥5 cm的乔木树种为乔木层,DBH≤4 cm 且H≤3 m的乔木树种为更新幼树(康冰等,2011),划分于更新层,其余木本植物为灌木层。本研究以重要值来测度各林层的植物优势度。乔木层重要值IV计算公式为:
IV=RD+RP+RF。
式中:RD为相对密度;RP为相对显著度;RF为相对频度。
更新层和灌木层IV计算公式为:
IV=RD+RF。
草本层IV计算公式为:
IV=RC+RF。
式中:RC为相对盖度。
2.3 生态位宽度计算 本次研究采用Levins生态位宽度(BL)(Levins, 1970)和Shannon-Wiener生态位宽度(BSW)(Colwelletal.,1971)来计算主要植物种群的生态位宽度,公式如下:
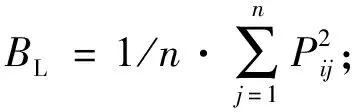
式中:n为样方数;Pij为物种i在j样方中的重要值占所有样方重要值总和的比例。BL具有域值[0, 1],当值为0时,生态位宽度最窄,当值为1时,生态位最宽。
2.4 生态位重叠度计算 各种群之间的生态位重叠度采用Pianka(1973)生态位重叠度公式来计算,公式如下:
式中:Oik为物种i和物种k之间的生态位重叠度,具有域值[0,1];Pkj为物种k在j样方中的重要值占所有样方重要值总和的比例。
2.5 数据处理 在Microsoft Excel 2016中进行数据校对和整理,并用于植物重要值、生态位宽度和生态位重叠度计算。使用SPSS21.0软件进行方差分析。
3 结果与分析
3.1 群落组成特征 固定样地内DBH≥1 cm的木本植物有18 816株,其中南方铁杉910株,占总株数的4.84%;林分密度为1 600株·hm-2;共有植物89种,隶属29科53属,个体数最多的是杜鹃花科(Ericaceae),其次为山茶科(Theaceae)和木兰科(Magnoliaceae);样地内胸高断面积最大的科为松科(Pinaceae),其次为壳斗科(Fagaceae)和杜鹃花科(陈斌等,2018)。从表1可看出,南方铁杉在针阔混交林乔木层中占有较大优势,其重要值高达59.11%。重要值大于20%的乔木树种有多脉青冈(Cyclobalanopsismultinervis)和闽皖八角(Illiciumminwanense),其余乔木树种的重要值皆低于10%。

表1 江西武夷山固定样地乔木层及更新层主要物种重要值
固定样地的更新层幼树共37种、1 432株,更新层幼树密度为224株·hm-2。闽皖八角在更新层的重要值最高,为68.51%;其次为老鸹铃(Styraxhemsleyanus),重要值为20.60%;多脉青冈的重要值排序第三,为18.72%。南方铁杉在更新层的重要值仅为5.65%,重要值排序为10,在更新层不具有优势。
由表2可知,灌木层的优势种是箭竹(Fargesiaspathacea),根据实地踏查结果,样地中箭竹盖度大于65%,平均高度高于1 m,且其地下茎发达。其余主要物种有岩柃(Euryasaxicola)、鹿角杜鹃(Rhododendronlatoucheae)、灯笼树(Enkianthuschinensis)和贵定桤叶树(Clethracavaleriei),重要值皆大于20%,明显高于其他灌木物种。草本层物种共有48种,共有5种草本的重要值大于10%,优势种有粉被苔草(Carexpruinosa)和钻地风(Schizophragmaintegrifolium)。
3.2 主要植物的生态位宽度 由表3可知,各层次主要物种的生态位宽度存在一定差异。一般来说,种群对环境适应能力强且对资源利用充分,则表现为较高生态位宽度。乔木层中Shannon-Wiener生态位宽度最大的物种为南方铁杉,其次为青冈(Cyclobalanopsisglauca),Levins生态位宽度最大的物种为青冈,其次为南方铁杉,这两个物种在每个样方中均有出现,表明南方铁杉和青冈的分布较广、数量较多、资源利用能力较高,对周围环境具有较强适应能力。而生态位宽度较小的叶萼山矾(Symplocosphyllocalyx)及天目紫茎(Stewartiagemmata)利用周围空间与资源的能力较其余主要乔木植物偏弱。
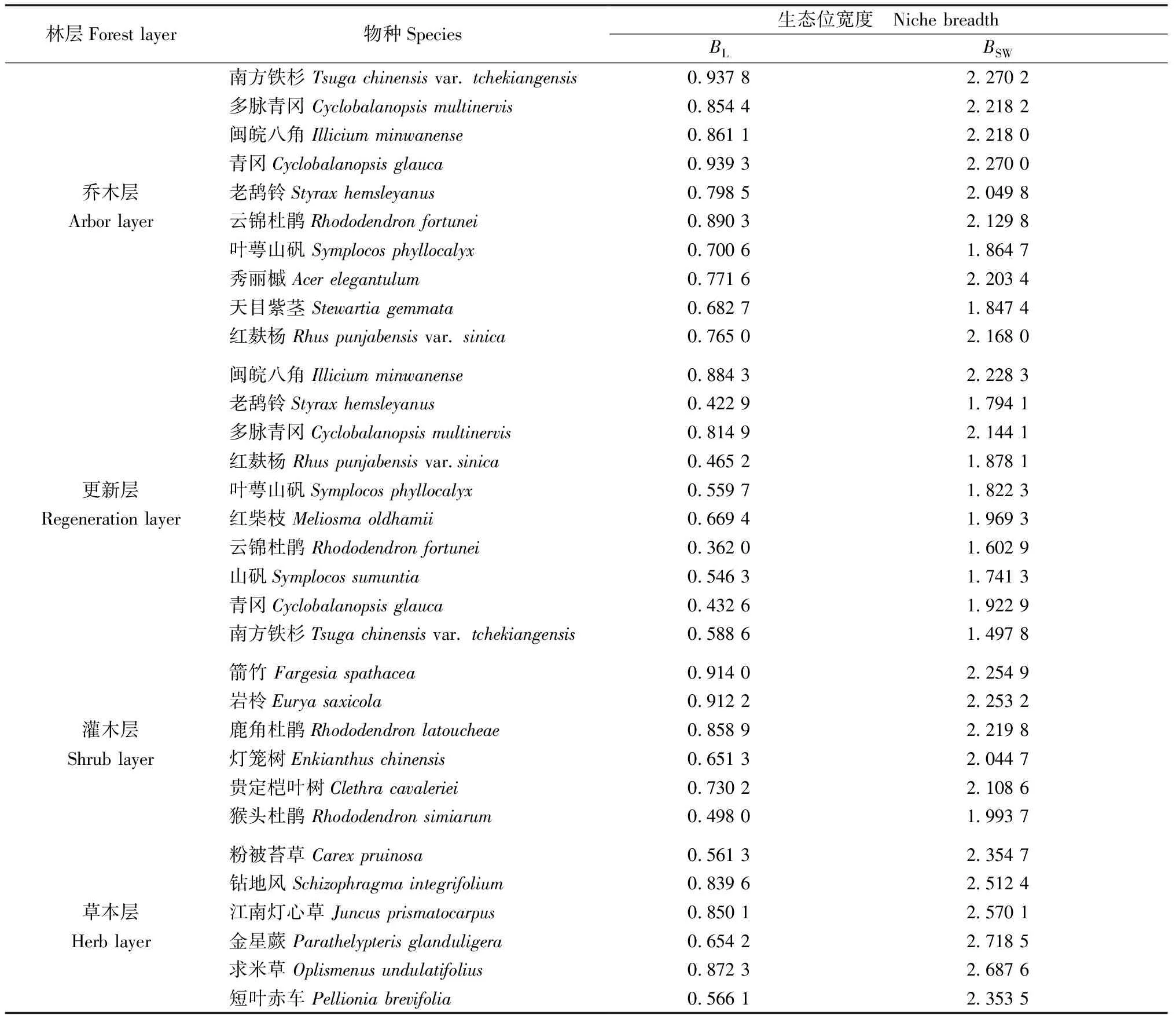
表3 江西武夷山固定样地各林层主要植物生态位宽度
更新层中闽皖八角的两种生态位宽度均最大,其次均为多脉青冈,闽皖八角和多脉青冈在更新层中资源利用能力较强,分布幅度较广。其余主要幼树的Levins生态位宽度皆低于0.669 4且Shannon-Wiener生态位宽度皆低于1.969 3,这些物种在更新层中资源利用能力较弱,其中,南方铁杉的生态位宽度较低,在更新层中优势度较差。与乔木层相比,老鸹铃、多脉青冈、红麸杨(Rhuspunjabensisvar.sinica)、叶萼山矾、云锦杜鹃(Rhododendronfortunei)、青冈、南方铁杉幼树的生态位宽度皆低于其在乔木层的生态位宽度,仅闽皖八角幼树的生态位宽度高于其在乔木层的生态位宽度,说明闽皖八角幼树在林下生境中利用资源的能力强,生态适应性较好。
重要值和生态位宽度都能表现种群的优势程度,但各层次中仅灌木层主要植物重要值的大小顺序与相对应的生态位宽度大小顺序基本相同。如灌木层箭竹的重要值与生态位宽度均为最大,猴头杜鹃(Rhododendronsimiarum)的重要值与生态位宽度均为最小。而在其他层次大小顺序均表现为不一致,如草本层中重要值最大的粉被苔草的生态位宽度最小。
3.3 主要植物之间的生态位重叠度 表4~7为4个层次中主要植物之间的生态位重叠度。生态位重叠度越大,两个物种利用资源的能力和性质越相似,越容易出现种间竞争关系;生态位重叠度越小,两个物种的资源利用能力和性质差异越大,意味着种群之间竞争关系不明显(简尊吉等,2017)。4个林层主要植物的生态位重叠度均值表现为乔木层(0.866 4±0.116)>灌木层(0.856 8±0.066)>草本层(0.645 8±0.147)>更新层(0.528 9±0.197),说明乔木层主要植物对资源利用的能力和性质的相似度最高,生态位分化不明显。更新层主要树种对资源和空间利用的能力和性质的相似度最低,生态位分化明显,表明南方铁杉幼树与其他更新层植物的种间竞争的强度和潜力较小。乔木层生态位重叠度最大的物种对是南方铁杉和多脉青冈(Oik=0.997 8),生态位重叠度最小的物种对为多脉青冈和红麸杨(Oik=0.501 6)。与南方铁杉的生态位重叠度较高的树种有多脉青冈、闽皖八角、青冈和红麸杨,这些树种与南方铁杉在生态因子联系上具有一定的相似性,对南方铁杉的竞争强度较高。

表4 江西武夷山固定样地乔木层主要植物生态位重叠度
表5显示更新层共有45组植物生态位重叠关系,其中有24组植物之间的生态位重叠度低于0.5,其余21组皆高于0.5。更新层中生态位重叠度最大的物种对是闽皖八角和多脉青冈(Oik=0.972 0),生态位重叠度最小的物种对是云锦杜鹃和青冈(Oik=0.130 0)。与南方铁杉幼树生态位重叠度较高的物种为云锦杜鹃和红麸杨,表明云锦杜鹃和红麸杨需要与南方铁杉利用更多的同一种资源或共同占有某一资源,对南方铁杉幼树的生长具有一定影响。
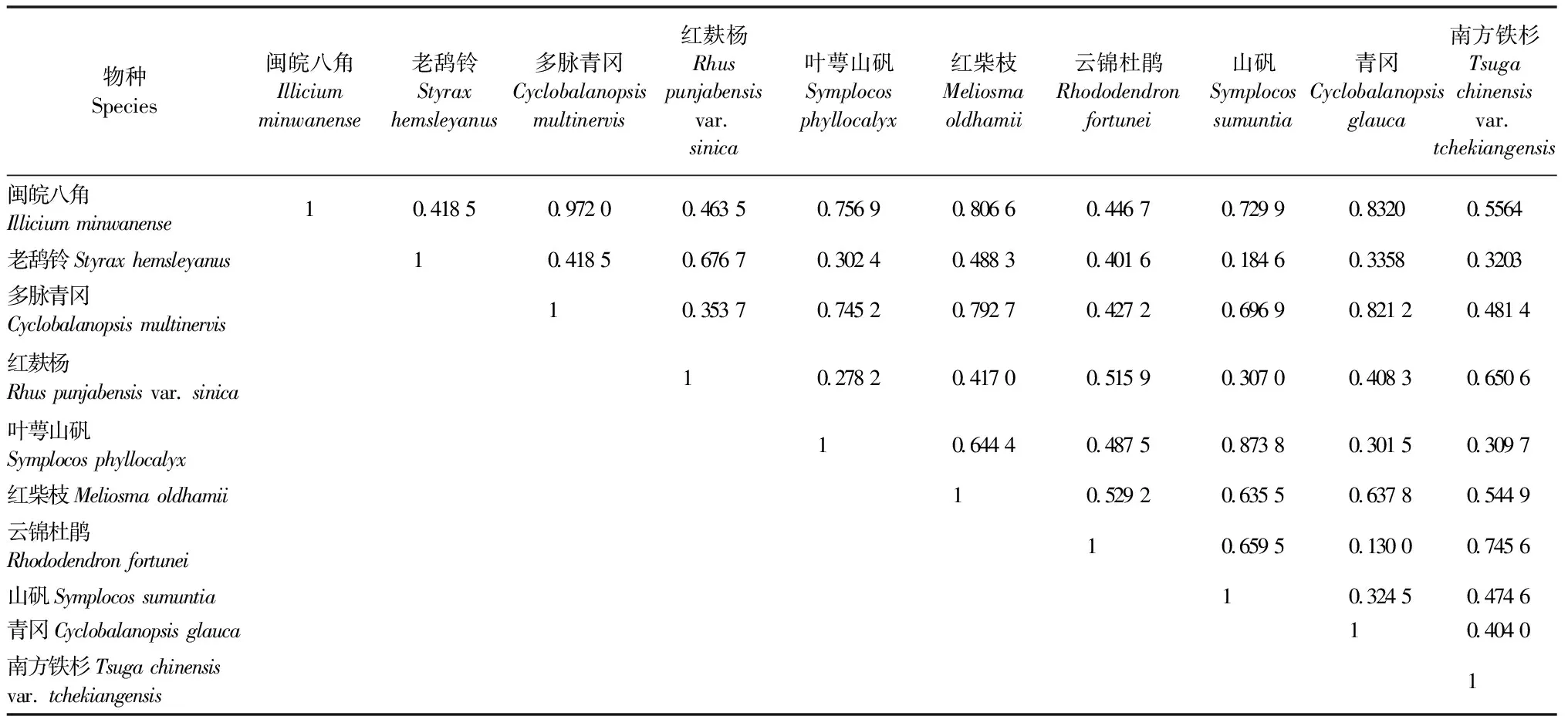
表5 江西武夷山固定样地更新层主要植物生态位重叠度
由表6和7可知:灌木层生态位重叠度最高的物种对为箭竹和岩柃(Oik=0.947 2),最小的为鹿角杜鹃和贵定桤叶树(Oik=0.729 7);草本层生态位重叠度最高的物种对为钻地风和求米草(Oplismenusundulatifolius)(Oik=0.907 5),最小的为钻地风和短叶赤车(Pellioniabrevifolia)(Oik=0.421 7)。
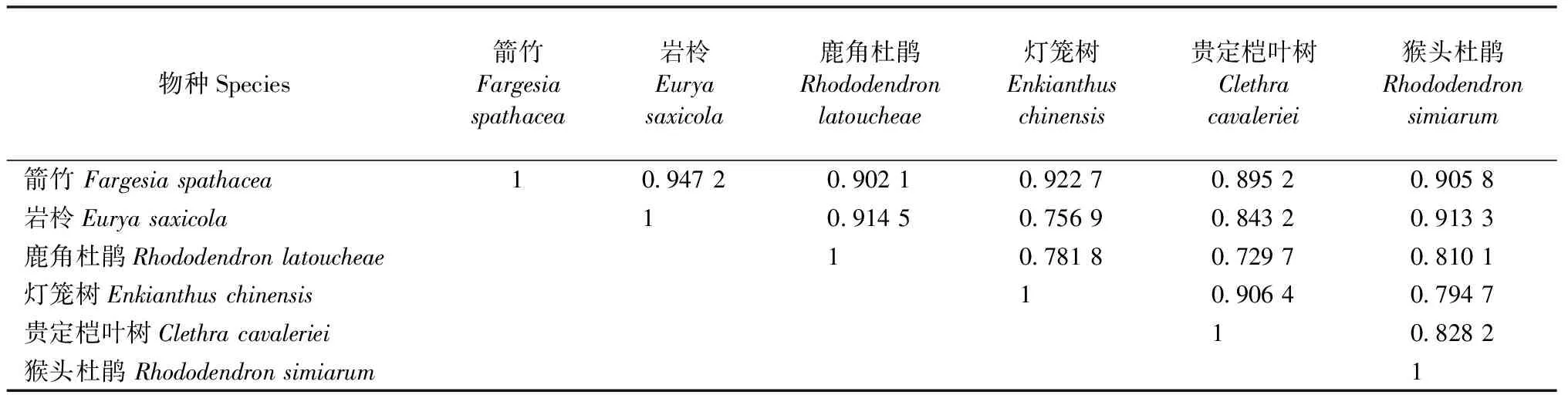
表6 江西武夷山固定样地灌木层主要植物生态位重叠度
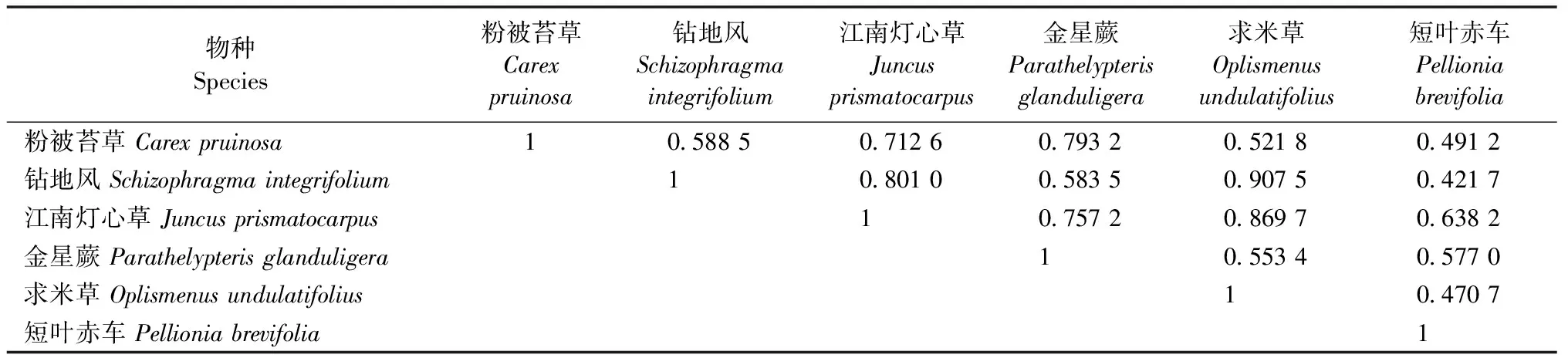
表7 江西武夷山固定样地草本层主要植物生态位重叠度
4 讨论
4.1 主要植物的生态位宽度 生态位宽度反映了物种的资源利用能力、生态适应性和分布幅度,揭示了各种群在群落中的功能地位(Arellanoetal.,2014)。本研究中乔木层生态位宽度最大的树种是南方铁杉和青冈,其次为多脉青冈和闽皖八角,是固定样地大径级和中径级个体的主要树种,与浙江九龙山(刘春生等,2009)的南方铁杉分布一致。南方铁杉喜光、根系发达、寿命长(罗金旺,2011),在江西武夷山仍有广泛分布,且大、中径阶个体是种群的主体部分,种群占据着优势地位,这与封磊等(2008)的研究结果类似,即:南方铁杉在福建武夷山黄山松(Pinustaiwanensis)林中数量不多,但其生态位宽度较大。乔木层生态位宽度最小的树种有叶萼山矾和天目紫茎,二者在样地中分布较少。本研究中南方铁杉幼树的生态位宽度明显低于南方铁杉大树的生态位宽度,这与杨清培等(2010)的研究结果不一致,这可能是因为南方铁杉在该群落中不具有优势,大树在优势层的竞争能力较弱。而在本样地中,南方铁杉是群落的建群种,随年龄增长,南方铁杉对环境的适应力逐渐增强,对环境资源的利用越来越多,并使天目紫茎等多数物种在不适的环境中退出,最终确定南方铁杉在群落中的优势地位和作用。此外,树种在不同生长阶段对资源和空间的需求和耐性差异导致了幼树和大树之间的生态位宽度或有不同(Brazetal.,2014),南方铁杉幼树对光照和生存空间的需求较高,相比于优势层,更新层的光照和生存空间受到限制,导致了南方铁杉幼树利用资源的能力较弱,这可能是南方铁杉幼树更新能力较差的原因。老鸹铃、多脉青冈、红麸杨、叶萼山矾、云锦杜鹃、青冈的大树与幼树的生态位宽度同样如此,但闽皖八角的变化趋势不一致,这可能与各树种不同的生物学和生态学特性有关(董冬等,2019)。
4.2 主要植物的生态位重叠度 生态位重叠度可反映不同种群对环境资源利用能力的相似性及种群间所占据空间位置的交错程度,一定程度上可表明物种间资源利用的竞争关系(Robertetal.,2003)。一般来说,在老龄林中各种群的动态变化相对稳定,种群调节以密度制约的自疏方式为主,因此种内竞争加强、种间竞争减弱,生态位重叠度普遍较低(林勇等,2017),而本研究中的乔木层各种群之间的生态位重叠度较高,均大于0.5,其均值高于温带群落主要种群之间的生态位重叠度(王惠等,2011),而又低于热带群落主要种群之间的生态位重叠度(吴庭天等,2017),生态位分化不明显,资源利用竞争较为激烈,种间竞争强度较大,说明群落还未至稳定状态,仍属于演替中后期阶段。更新层的树种组成及竞争关系一定程度上决定了群落的演替方向,本研究中更新层主要树种之间的生态位重叠度较小,生态位分化明显,竞争强度较弱。
生态位宽度越大,物种资源利用能力越强,分布幅度越广,更易与其他种群产生较大生态位重叠度(Rosasetal.,2019)。乔木层生态位宽度最大的南方铁杉和青冈之间的生态位重叠度最大,两者竞争关系较强,因此在群落的演替过程中,当南方铁杉幼树更新困难时,建群种南方铁杉有可能被青冈替代。另一方面,群落演替既是物种对环境的适应过程,也是环境对物种的自然选择过程(潘高等,2017),在广西的元宝山冷杉(Abiesyuanbaoshanensis)群落中,元宝山冷杉虽与南方铁杉有较大的生态位重叠度,但元宝山冷杉与南方铁杉相互依赖性强,两者之间具有原始协作的种间关系,使该群落成为稳定的顶级群落(向悟生等,2002)。在种间竞争和资源环境压力的作用下,南方铁杉和青冈也有趋于适应、协同进化的可能。叶萼山矾和天目紫茎的生态位宽度较小,与南方铁杉、多脉青冈等大乔木的生态位重叠度也较低,在乔木层与南方铁杉的潜在竞争强度较弱。更新层中生态位宽度较大的闽皖八角和多脉青冈与其他幼树的生态位重叠度较大,对幼树天然更新具有一定的影响。但是与南方铁杉幼树生态位重叠度最高的物种是云锦杜鹃和红麸杨,表明南方铁杉幼树具有很大的更新潜力。
箭竹是灌木层的优势种,在样地中广泛分布,且地下茎发达,箭竹入侵很有可能对南方铁杉幼树生长产生胁迫,导致种群更新能力降低。岩柃多生于矮林和密阴处,具有很强的耐荫耐寒性和适应能力,在该群落演替过程中个体分布广泛而均匀。因此,为保障珍稀树种南方铁杉的正常更新,可对岩柃和箭竹种群数量进行适当调控。
5 结论
乔木层中南方铁杉和青冈利用资源能力较强,中、大径级个体是种群的主要部分,且两者具有较强的竞争关系。南方铁杉幼树在更新层的光照和生存空间受到限制,导致了重要值和生态位宽度低,更新状况较差。青冈等耐荫性树种的幼苗和大树皆有较高的优势度,南方铁杉可能被青冈等树种替代。生态位重叠度的研究结果表明南方铁杉幼树仍具有一定的更新潜力,因此为维持南方铁杉原始林的稳定性以及完善其管理措施,可适当采用择伐等人为干扰措施增加林下光照,不仅可为种子传播提供良好的通风条件,也可为幼苗幼树生长营造适宜生境。
猜你喜欢
北华大学学报(自然科学版)(2022年5期)2022-11-21
现代园艺(2020年18期)2020-12-22
农村实用技术(2020年1期)2020-12-16
新农民(2020年2期)2020-12-08
报刊荟萃(上)(2017年7期)2017-07-16
魅力中国(2016年29期)2017-05-27
读写算·小学中年级版(2017年4期)2017-05-08
江苏农业科学(2016年6期)2016-07-25
学与玩(2009年2期)2009-03-09
北京文学(2007年5期)2007-05-23
