
旱盐交叉胁迫对甜高粱种子萌发和生理特性的影响
2021-03-13 07:32:32王志恒杨秀柳邹芳黄思麒周吴艳徐中伟魏玉清
中国农业科技导报 2021年2期
王志恒, 杨秀柳, 邹芳, 黄思麒, 周吴艳, 徐中伟, 魏玉清
(北方民族大学生物科学与工程学院, 国家民委生态系统模型及应用重点试验室, 银川 750021)
土壤干旱和盐渍化是农业发展的重要限制因子,严重威胁作物产量和农村经济发展[1-2]。我国干旱和半干旱土地面积超过全国总土地面积的二分之一,其中盐渍化土壤面积达到0.346×108hm2[3-4]。由于干旱和半干旱地区具有降雨量偏少、土壤盐分含量高等特点,同时人类对环境破坏加剧以及不合理的耕作方式,导致土壤次生盐渍化程度不断加深,使得在这些地区生长的植物受到干旱-高盐的复合逆境[5-6]。甜高粱[Sorghumbicolor(L.) Moench]又称糖高粱、甜秆等,作为一种优质的饲料作物,具有适应能力强、营养价值和生物产量高等特点,在干旱半干旱和盐渍地区均适于种植[7]。种子萌发阶段作为植物对外界环境最敏感的时期,极易受到外界环境的影响[8]。因此,研究旱盐交叉胁迫对甜高粱种子萌发的影响,对干旱半干旱地区甜高粱生产具有重要意义。
目前,甜高粱种子萌发期响应单一干旱或盐胁迫方面已有大量研究。Patanè等[9]研究发现,在受到水势低于-0.6 MPa的水分胁迫时,甜高粱种子发芽率开始有所降低;在水势等于-0.4 MPa时,初始发芽时间延迟。杨帆等[10]利用100 mmol·L-1的NaCl和等渗的甘露醇分别模拟盐胁迫与干旱胁迫,结果表明,不同品种甜高粱萌发和幼苗生长都受到了不同程度的抑制。与干旱胁迫类似,盐胁迫使甜高粱种子发芽率、发芽势、发芽指数、活力指数等萌发指标逐渐降低,同时显著抑制干物质转移和早期幼苗的生长,对幼根的抑制高于幼芽[11]。在自然条件下,植物常常面临多种外界胁迫,在遭受某种胁迫后,植物不仅增强对这种胁迫条件的耐受力,同时也会提高其他胁迫的耐受力,这种对外界环境不同胁迫间的相互适应作用称为植物的交叉适应[12]。目前,有关旱、盐胁迫对甜高粱的影响已有一些报道,但都是单因素胁迫,关于旱盐交互胁迫尚未见报道。因此,本研究采用PEG-6000模拟干旱胁迫,采用NaCl模拟盐胁迫,研究旱盐交叉胁迫对甜高粱种子萌发以及生理生化特性的影响,以期为甜高粱响应复合胁迫的生理机制提供理论参考。
1 材料与方法
1.1 供试材料
供试种子为甜高粱杂交品种辽甜1号,由辽宁省农业科学院提供。
1.2 试验处理
利用PEG-6000用蒸馏水配置成0(CK)、50(D1)、100 g·L-1(D2)的溶液;将NaCl配置成0(CK)、75(S1)、150 mmol·L-1(S2)盐溶液;以及二者交互胁迫D1+S1、D2+S1、D1+S2、D2+S2,共计9个处理。
1.3 测定项目及方法
种子试验处理及指标计算公式[6]如下。
发芽率=萌发种子数/供试种子数×100%
发芽势=第3 d发芽种子数/供试种子数×100%
发芽速度=∑G/t
式中,G为每天的发芽率,t为发芽总时间。
活力指数=∑Gt/t×S
式中,Gt为t天内发芽的种子数,S为胚根平均长度。
干物质转移量=发芽前种子干质量-发芽后种子残留干质量
干物质转运率=(芽干重+根干重/芽干重+根干重+种子残留干重)×100%
1.4 生理指标测定
超氧化物歧化酶(superoxide dismutase,SOD)活性测定采用氮蓝四唑光还原法[13];过氧化物酶(peroxidase,POD)活性测定采用愈创木酚显色法[14];过氧化氢酶(catalase,CAT)活性测定采用紫外吸收比色法[15];抗坏血酸过氧化物酶(ascorbate peroxidase,APX)活性测定参考孙云[16]的方法;可溶性蛋白含量测定采用考马斯亮蓝G-250法[17];脯氨酸(proline,Pro)含量测定采用南京建成生物工程研究所的脯氨酸测试盒;丙二醛(malondialdehyde,MDA)含量采用硫代巴比妥酸法[17];还原性谷胱甘肽(glutathione,GSH)含量测定参考孙群等[17]的方法。
1.5 耐受性综合评价
应用隶属函数法对不同胁迫条件下甜高粱种子萌发的伤害作用进行综合评价。在综合评价中,不同胁迫下的各个指标以隶属值来表示。若某一指标与伤害作用为正相关,采用式(1)计算隶属值;若为负相关,则采用式(2)计算隶属值[18]。
X=(X-Xmin)/(Xmax-Xmin)
(1)
X=1-(X-Xmin)/(Xmax-Xmin)
(2)
式中,X为某一指标测定值,Xmin某一指标测定值的最小值,Xmax为该指标中最小值。
最后将各个指标隶属值累加求平均值,平均值越大则伤害越小,根据平均隶属值来比较其伤害大小。
1.6 数据处理
采用 Microsoft Excel 2003进行数据统计处理,使用GraphPad Prism 5.0作图,利用SPSS 21.0进行双因素方差分析。图表中参数数据以平均值±标准差表示。
2 结果与分析
2.1 旱盐交叉胁迫对甜高粱种子萌发指标的影响
由图1可知,单一胁迫下,甜高粱的萌发指标均随着胁迫浓度增大整体呈降低趋势。D2+S2处理下,发芽速度和活力指数显著低于CK处理(P<0.05),并且随着浓度增大呈减少趋势。交互胁迫处理能够抑制甜高粱种子的萌发指标,其中D2+S2处理的发芽率和发芽势显著低于CK处理(P<0.05)。低浓度的旱盐交互作用可以提高发芽速度和活力指数,同一S2处理下,通过添加50 g·L-1的PEG-6000可以提高发芽速度和活力指数;过高的PEG-6000浓度会降低种子发芽速度和活力指数。但在S1处理下,不同浓度的PEG-6000处理均可降低种子的发芽速度和活力指数。以上结果表明,在特定的NaCl处理下,低浓度的PEG-6000处理能提高甜高粱种子对盐环境的耐受力,而PEG-6000浓度过高则会产生相反的效应。双因素方差分析(表1)表明,对于发芽率和发芽势,从D、S及D+S处理所占总平方和的百分比(PP)来看,D处理贡献度较高,D+S处理次之,S处理最小;对于发芽速度和活力指数,D处理对其的贡献度较高,S处理次之,D+S处理最小。因此,D处理对萌发指标的影响较大。
2.2 旱盐交叉胁迫对甜高粱种苗转化的影响
由图2可知,单一胁迫均可抑制甜高粱种苗转化指标。除D1处理外,其余单一胁迫均可显著抑制甜高粱早期幼苗的芽长、根长、总长及侧根数(P<0.05),并随着处理浓度的增大呈逐渐减小趋势,其中D处理对芽长的抑制大于根长。交互胁迫处理能够抑制甜高粱种苗转化指标,当PEG-6000浓度为50 g·L-1时,甜高粱的芽长、根长及总长随着NaCl浓度的增大呈逐渐减小趋势;当PEG-6000浓度达到100 g·L-1时,甜高粱的芽长随着NaCl浓度的增大呈先增大后减小趋势,根长和总长则呈逐渐减小趋势。而对于芽干重+根干重、种子残留干重、干物质转移量及干物质转运率均随着单一胁迫的程度加深而逐渐抑制,交叉胁迫下,这四类指标同样随着胁迫程度的加深逐渐被抑制。双因素方差分析(表2)表明,从D、S及D+S处理所占总平方和的百分比(PP)来看,对于芽长、总长、种子残留干重和干物质转移量,S处理的贡献度较高,D处理次之,D+S处理最小;对于根长、芽干重+根干重和干物质转运率,D处理的贡献度较高,S处理次之,D+S处理最小;对于侧根数,D+S处理的贡献度较高,S处理次之,D处理最小。因此,旱盐交叉胁迫对甜高粱种苗转化指标的影响各有不同。
2.3 旱盐交叉胁迫对甜高粱早期幼苗生理指标的影响
2.3.1旱盐交叉胁迫对甜高粱早期幼苗抗氧化物酶活性的影响 图3显示,在PEG-6000或NaCl单一胁迫下,甜高粱早期幼苗的幼芽和幼根SOD、POD、CAT、APX活性随胁迫浓度的增大呈先升后降趋势,说明低浓度的干旱或盐胁迫可以提高甜高粱早期幼苗抗氧化酶活性。但在D2+S2处理下,甜高粱的幼芽和幼根抗氧化物酶活性显著低于CK处理(P<0.05),这可能是因为该胁迫程度超过了甜高粱早期幼苗过氧化防御系统的阈值。相比于单一胁迫,交叉胁迫下,甜高粱早期幼苗的抗氧化物酶活性呈现先升高后下降趋势,说明低浓度交叉胁迫可以提高抗氧化物酶活性,而高浓度的交叉胁迫则明显抑制抗氧化物酶活性。双因素方差分析(表3)表明,从D、S及D+S处理所占总平方和的百分比(PP)来看,甜高粱早期幼苗中幼芽和幼根的SOD、POD、CAT、APX活性,D处理的贡献度较高,S处理次之,D+S处理最小。因此,干旱胁迫对甜高粱幼苗抗氧化物酶活性的影响较大。
2.3.2旱盐交叉胁迫对甜高粱早期幼苗可溶性蛋白、Pro、MDA和GSH含量的影响 图4显示,在PEG-6000或NaCl单一胁迫下,甜高粱早期幼苗的幼芽和幼根的可溶性蛋白、Pro含量随胁迫浓度的增大呈先升高后降低趋势。D2+S2处理下,甜高粱的幼芽和幼根可溶性蛋白、Pro含量显著低于CK处理(P<0.05)。而MDA、GSH含量随着胁迫浓度的增大呈升高趋势。相比于单一胁迫,交叉胁迫下,甜高粱早期幼苗的可溶性蛋白、Pro含量呈现先升高后下降趋势,说明低浓度交叉胁迫可以提高可溶性蛋白、Pro含量,而高浓度的交叉胁迫则降低可溶性蛋白、Pro含量。交叉胁迫下,甜高粱早期幼苗的MDA、GSH含量随着胁迫程度的加深呈现逐渐升高的趋势。双因素方差分析(表4)表明,从D、S及D+S处理所占总平方和的百分比(PP)来看,甜高粱早期幼苗中可溶性蛋白含量,D+S处理的贡献度较高,S处理次之,D处理最小;而D处理对Pro含量的贡献度较高,S处理次之,D+S处理最小;S处理对MDA和GSH含量的贡献度较高,D处理次之,D+S处理最小。因此,旱盐交叉胁迫对可溶性蛋白含量的影响较大,干旱胁迫对Pro含量的影响较大,盐胁迫是影响MDA和GSH含量的主要因子。

表2 旱盐交叉胁迫对甜高粱种子种苗转化指标影响的双因素方差分析Table 2 Two-factor ANOVA of the effect of drought salt cross stress on seed-seedling transformation of sweet sorghum

表3 旱盐交叉胁迫对甜高粱早期幼苗抗氧化物酶活性影响的双因素方差分析Table 3 Two-factor ANOVA of the effect of drought salt cross stress on antioxidant enzyme activity of sweet sorghum at early seedling

表4 旱盐交叉胁迫对甜高粱早期幼苗可溶性蛋白、Pro、MDA和GSH含量影响的双因素方差分析Table 4 Two-factor ANOVA of the effect of drought salt cross stress on soluble protein, Pro, MDA and GSH content of sweet sorghum at early seedling

注:不同小写字母表示差异在P<0.05水平具有显著性。Note: Different small letters indicate significant difference at P<0.05 level.图3 旱盐交叉胁迫对甜高粱早期幼苗抗氧化物酶活性的影响Fig.3 Effects of drought salt cross stress on antioxidant enzyme activity of sweet sorghum early seedling
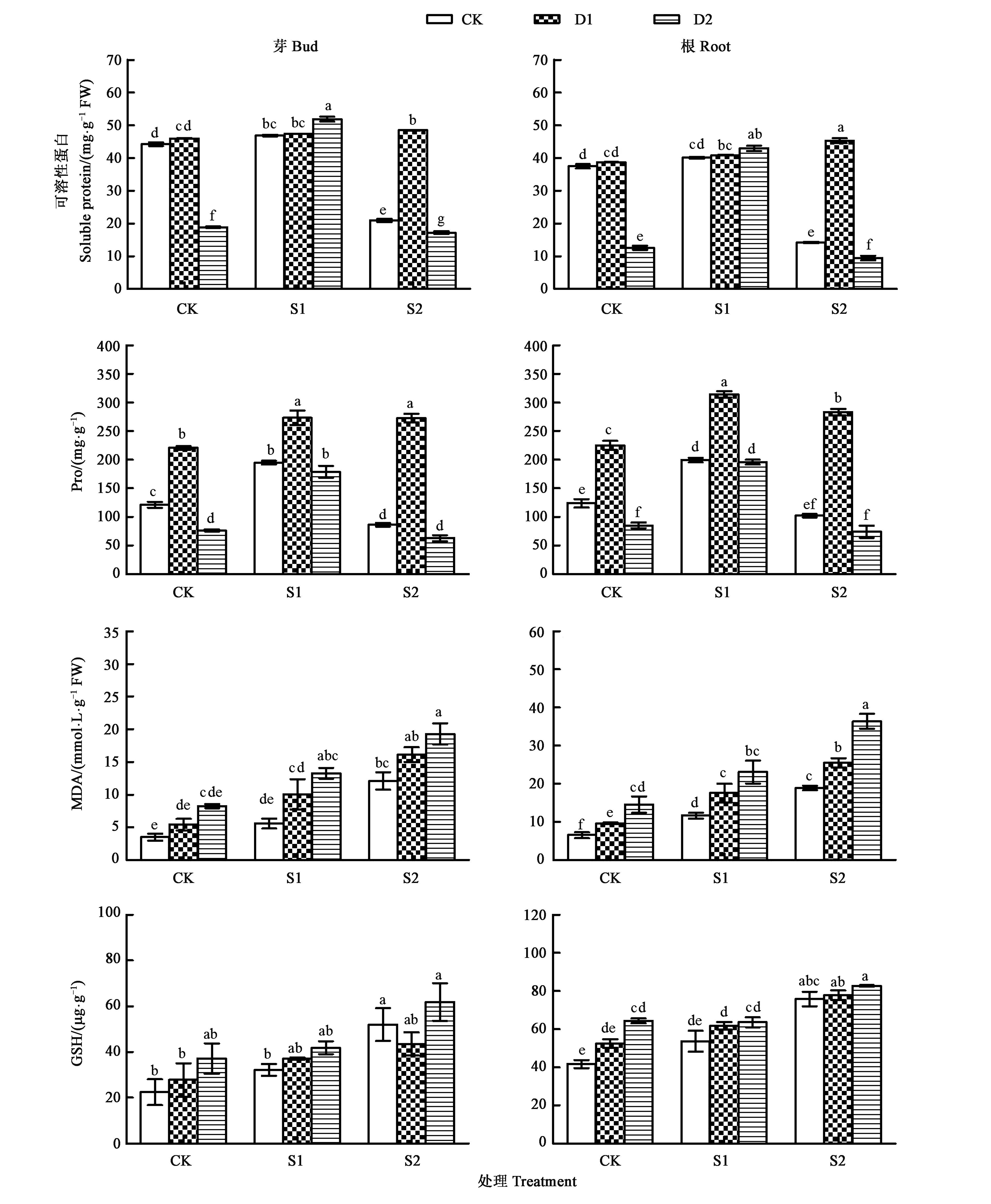
注:不同小写字母表示差异在P<0.05水平具有显著性。Note: Different small letters indicate significant difference at P<0.05 level.图4 旱盐交叉胁迫对甜高粱早期幼苗可溶性蛋白、Pro、MDA和GSH含量的影响Fig.4 Effects of drought salt cross stress on soluble protein, Pro, MDA and GSH content of sweet sorghum early seedling
2.4 相关性分析
20个指标的相关分析(表5)表明,发芽率、发芽势、发芽速度及活力指数之间,芽长、根长、总长、侧根数、芽干重+根干重、干物质转移量及干物质转移率之间均呈极显著正相关;种子残留干重与干物质转移率、干物质转移率呈极显著负相关;SOD活性、POD活性、CAT活性、APX活性、可溶性蛋白含量、Pro含量之间均呈极显著正相关;MDA含量、GSH含量与SOD活性、POD活性及可溶性蛋白含量呈极显著负相关。从20个指标相关系数矩阵可以看出,所有指标间都存在着相关性,从而使得它们所提供的信息发生重叠。因此,直接利用这些单个指标进行伤害作用评价可能具有不可靠性。
2.5 单一胁迫与交互胁迫差异比较及综合评价
本研究为探究甜高粱对单一胁迫与交互胁迫条件的差异比较,采用隶属函数法分别对单一胁迫及交互胁迫处理下的伤害作用进行全面、综合的评价。由表6可知,隶属函数结果分析评价出各个处理对甜高粱种子萌发期的伤害作用大小依次为:D2+S2>S2>D2+S1>D2>D1+S2>D1+S1>S1>D1>CK。通过以上种子萌发情况及幼苗生理指标的变化分析及隶属函数法综合评价发现,甜高粱种子萌发阶段具有植物的交叉适应特点。
3 讨论
种子萌发期是植物生长发育的关键时期,决定植物群体在逆境条件下能否建植成功。发芽率、发芽势、发芽指数和活力指数是评价种子活力的常用指标,可以反映出种子的萌发能力[19]。植物从种子转化到幼苗过程中所需的物质及能量是由种子中贮藏的有机物提供,因而种子萌发期的总长、芽长、根长、侧根数、种子残留干重、芽干重+根干重和干物质转运率能体现出植物种苗转化状况,当植物早期幼苗胚芽和胚根生长均衡时,可以表现出更好的生长状况[6]。干旱和盐胁迫对植物体均能造成渗透效应,除此之外,盐胁迫可造成植物体内离子平衡失调、有毒物质积累等,从而抑制植物生长发育,胁迫条件下植物的发芽率及外部形态胁迫影响的直接体现,受影响越大,发芽率越低,导致出苗不齐,从而影响作物产量[20]。张宇君等[21]对燕麦的研究指出,中低浓度的单一胁迫及交互胁迫均有利于促进燕麦种子萌发和增强幼苗SOD、POD、CAT活性以及提高GSH含量,当胁迫程度较高时,原本的促进作用转变为抑制作用。在本研究中,单一及交互胁迫下甜高粱种子的萌发指标随着胁迫加重呈现抑制作用,然而在高浓度的NaCl处理下,一定浓度的PEG-6000处理能提高甜高粱种子的萌发指标和种苗转化指标,而PEG-6000浓度过高则会产生相反的效应,这与赵丽丽等[22]在二色胡枝子的研究结果一致。
非生物胁迫下,植物会产生大量活性氧自由基,大量的活性氧会影响蛋白质的合成、破坏膜系统,造成植物细胞严重损坏甚至死亡,SOD、POD、CAT和APX是细胞内清除活性氧的主要保护酶,这4种酶彼此协调的综合结果是整个保护酶系统防御能力的体现[23]。本研究中,在单一及交互胁迫下,甜高粱早期幼苗的SOD、POD、CAT和APX活性随着浓度增大呈现先升高后下降趋势,抗氧化物酶活性的升高有利于清除植物遭受胁迫后产生活性氧自由基,而当胁迫伤害较大时,这4种抗氧化物酶已经不能完全清除活性氧自由基,因此活性降低。这与姚海梅等[20]对玉米的研究结论相似。
逆境胁迫下植物体内可溶性蛋白含量的增加使植物维持更低的渗透势,从而增强抗逆性,Pro通常被认为是植物在盐胁迫下用于调节细胞质和液泡渗透势平衡的一种主要的渗透物质,GSH是植物细胞中重要的抗氧化剂之一,其含量与膜脂过氧化程度呈正相关,而叶片MDA含量则反映其细胞氧化损伤程度[17,24]。在本研究中,单一胁迫条件下,随着胁迫浓度的增加可溶性蛋白和Pro含量呈现先升高后下降趋势,MDA和GSH含量呈现升高趋势;150 mmol·L-1的NaCl处理显著降低可溶性蛋白和Pro含量,显著提高MDA和GSH含量,在加入50 g·L-1的PEG-6000后,甜高粱幼苗、幼根中的相应指标显著升高,说明适量干旱条件可以清除ROS的积累,从而减轻膜脂过氧化程度,维持了细胞膜的完整性,并且可以提高可溶性蛋白含量,减轻氧化胁迫,从而保证生理生化活动的进行。由此表明,适度干旱可以减轻盐胁迫对甜高粱早期幼苗的伤害,陈成升等[25]的研究指出,小麦幼苗的可溶性蛋白和Pro含量随着干旱胁迫的加重呈现先增加后减少趋势,在干旱胁迫的条件下加入适量的盐分可以提高可溶性蛋白和Pro含量,这与本研究结果类似。
本研究分析了甜高粱对干旱、盐单一胁迫及二者交互胁迫的萌发和生理响应特征,发现特定条件的交互胁迫可以促进种子萌发及增强抗氧化物酶活性,提高渗透调节物质和抗氧化剂含量。萌发指标、种苗转化指标及生理指标在双因素方差分析结果中,干旱胁迫显著影响甜高粱种子的萌发指标;对于种苗转化指标,干旱和盐以及二者交互作用的影响各不相同;而对于生理指标,干旱胁迫是影响抗氧化物酶活性、MDA和GSH含量的主要因子,盐胁迫则是影响可溶性蛋白含量的主要因子。隶属函数法结果表明:相比于单一胁迫,特定条件下交叉胁迫的平均隶属函数值更小,并且在交叉胁迫下甜高粱能通过干旱、盐交互作用来提高自身对外界环境的适应性,表现出植物的交叉适应,从而提高自身耐受性。

表5 各指标的相关性分析Table 5 Correlation analysis of each index

表6 旱盐交叉胁迫下甜高粱耐受性综合评价Table 6 Comprehensive evaluation of sweet sorghum tolerance under drought salt cross stress
猜你喜欢
快乐作文(1.2年级)(2023年9期)2023-05-12 11:31:34
古今农业(2022年2期)2022-08-15 01:39:52
青年文学家(2022年1期)2022-03-11 12:27:39
初中生世界·八年级(2019年6期)2019-08-13 18:41:18
中成药(2018年8期)2018-08-29 01:28:34
小学生导刊(低年级)(2016年6期)2016-07-02 22:17:33
计算机工程(2015年8期)2015-07-03 12:19:54
医学研究杂志(2015年9期)2015-07-01 17:28:24
中国当代医药(2015年9期)2015-03-01 02:02:13
振动、测试与诊断(2014年6期)2014-03-01 01:14:47
