
施氏獭蛤早期生长性状遗传参数和育种值估计
2021-03-11 05:44张守都彭慧婧郑德斌
海洋科学 2021年2期
邹 杰, 张守都, 彭慧婧, 郑德斌
(1. 广西壮族自治区海洋研究所, 广西 北海 536000; 2. 青岛国家海洋科学研究中心, 山东 青岛 266071; 3. 天津渤海水产研究所, 天津 300457)
施氏獭蛤(Lutraria sieboldii)隶属瓣鳃纲(Lamellibranchia)、异齿亚纲(Heterodonta)、帘蛤目(Veneroida)、蛤蜊科(Mactridae)、獭蛤属(Lutraria), 俗称象鼻螺, 为北部湾潮下带重要经济贝类之一。施氏獭蛤适合底播增养殖, 部分群体也能适应前中期的筐式养殖, 近年来养殖规模不断增长, 但养殖群体生长性状出现衰退, 为此, 施氏獭蛤相关的选择育种工作随之开展。生长性状选择育种是动物育种传统且有效的方法, 通过对目标性状的累代选育, 获取优良生长性状的品种。生长性状受遗传和环境因素共同影响, 从数量性状的遗传机制可知, 影响数量性状的效应中只有加性效应才能稳定遗传给后代, 准确估计遗传参数和育种值是数量性状育种的核心。约束极大似然法(restricted maximum likelihood, REML)是动物育种遗传参数估计中较为常用方法[1-2], 最佳线性无偏预测法(best linear unbiased prediction, BLUP)在获取的个体育种值方面具较高精确性[3], 本研究以施氏獭蛤 2 代选育群体为亲本建立全同胞家系, 基于REML 和BLUP 法对早期生长性状遗传参数和育种值进行估计和分析, 为施氏獭蛤选择育种提供基础资料和参考。
1 材料与方法
1.1 早期家系建立
从施氏獭蛤第二代选育群体(F2)中挑选体型相近的7 个雄性和21 个雌性为亲本, 采用巢式设计构建21 个全同胞家系(M1、M2、M3、……、M21)。通过1 雄配3 雌及人工授精方式获得的幼体分家系分别培养于1 000 L 的水桶中, 用水经紫外线消毒,每3 d 倒桶并调整幼体密度差异, 培育前期以投喂湛江等鞭金藻(Isochrysis zhanjiangensis)为主, 后期以投喂亚心型扁藻(Platymonas subcordiformis)为主, 保持各家系培育操作一致, 附着期保留每桶幼体数4 万~5 万个。幼体培育30 d 后将苗种收集以塑料筐装沙沉水方式进行海上培育, 培育密度0.5 个/cm2, 每个家系留3 000 个体, 海上培育50 d。
1.2 生长性状测量
用游标卡尺测量壳长、壳高和壳宽, 精确至0.01 mm;用电子天平测量体质量, 精确至0.01 g。每家系取样30 个体, 30 日龄测量壳长和壳高, 80 日龄测量壳长、壳高、壳宽和体质量。
1.3 数据处理与分析
用SPSS 22.0 统计软件对测量数据进行正态性检验(Kolmogorov-Smirnov 法), 计算各性状表型值(phenotypic value, PV)与育种值(breeding value, BV)的Pearson 相关系数。
建立混合线性模型(动物模型), 利用ASReml-R V4 软件采用REML 和BLUP 法对施氏獭蛤进行遗传参数和育种值估计。单性状模型估计遗传力(heritability,h2)和育种值, 两性状模型估计表型相关系数(rp)和遗传相关系数(rg)。
混合线性模型:

式中,yij为性状测量值,μ为平均值,ai为加性遗传效应,fj为全同胞效应,eij为残差。
性状1 和性状2 的表型相关系数和遗传相关系数计算依据公式:

2 结果与分析
2.1 施氏獭蛤生长性状描述
施氏獭蛤生长性状参数统计见表1, 30 日龄和80 日龄的施氏獭蛤生长性状测量值经Kolmogorov-Smirnov 检验均符合正态分布。80 日龄4 个性状中,壳长、壳高和壳宽性状的变异系数(coefficient of variation, CV)相近, 体质量变异系数最高, 80 日壳长和壳高性状变异系数较30 日龄减少, 差异变小。对30 日龄和80 日龄壳长与壳高性状比较, 极值差比为1.61 和1.89, 均值比为1.53 和2.09, 均表明施氏獭蛤早期发育过程中壳长生长速度较壳高快。
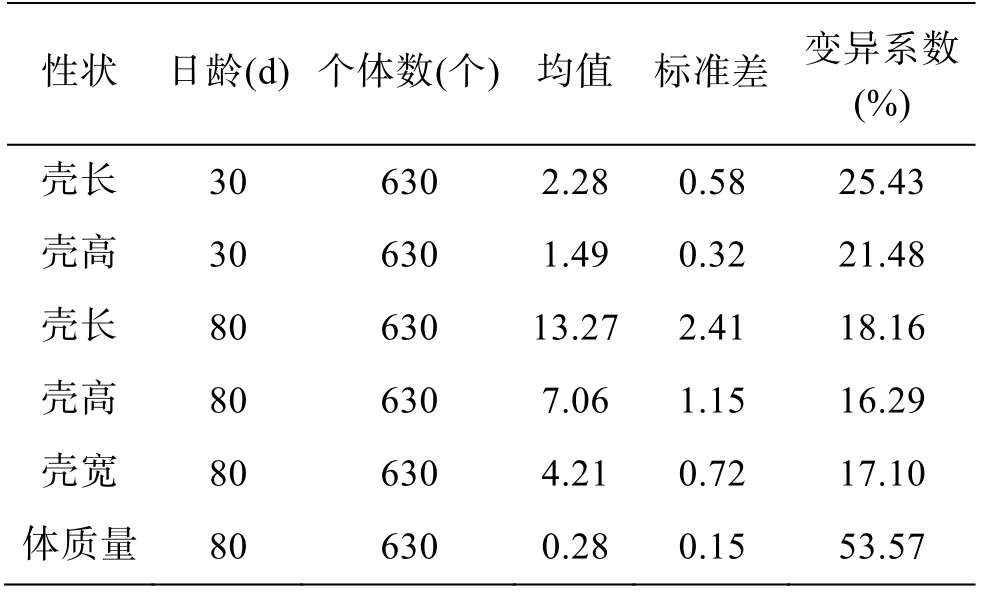
表1 施氏獭蛤生长性状参数统计Tab. 1 Parametric statistics of the growth traits of Lutraria sieboldii

表2 施氏獭蛤生长性状方差组分及遗传力Tab. 2 Variance components estimation of the growth traits and the heritability of Lutraria sieboldii
2.2 遗传参数估计
单性状分析施氏獭蛤生长性状的方差组分和估算遗传力结果见表2。30 日龄的壳长和壳高性状遗传力分别为: 0.31±0.02 和0.25±0.01, 80 日龄的壳长、壳高、壳宽和体质量性状的遗传力分别为: 0.35±0.27、0.27±0.20、0.06±0.04 和0.25±0.19, 壳宽性状遗传力为低遗传力, 其余均属中等遗传力(h2<0.2 为低遗传力;0.2≤h2≤0.4 为中等遗传力;h2>0.4 为高遗传力[4])。80 日龄的壳长和壳高性状的遗传力较30 日龄略有提高, 表明随时间增长全同胞效应降低。两性状分析施氏獭蛤生长性状间的表型相关和遗传相关结果见表3。80 日龄4 个性状间的遗传相关与表型相关均呈正相关且表现一致性, 遗传相关系数范围为 0.96~0.98,相关性均较高, 表型相关系数范围为0.77~0.96, 壳长、壳高与体质量间表型相关性较高, 结果表明在对早期施氏獭蛤体质量性状选择上, 壳长和壳高可作为主要参考性状。80 日龄壳长和壳高性状的遗传相关和表型相关均高于30 日龄, 表明在早期育种性状参考上, 80 日龄的壳长、壳高性状参考价值更高。
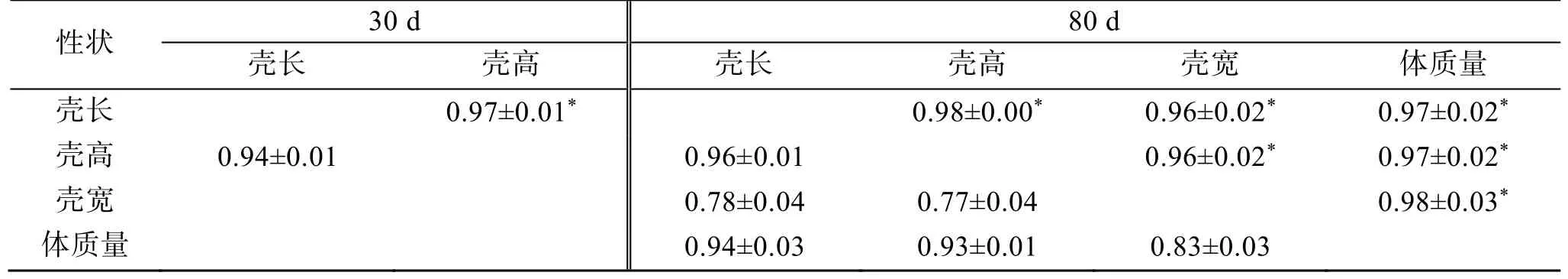
表3 施氏獭蛤生长性状表型相关和遗传相关Tab. 3 Genetic correlation and phenotypic correlation of the growth traits of Lutraria sieboldii

表4 施氏獭蛤不同性状育种值的家系排序Tab. 4 The ordered families arrangement of breeding values of different traits of Lutraria sieboldii

表5 基于表型值和育种值的个体选择比较(10%留种率)Tab. 5 Comparison of individual selection based on the phenotypic and breeding values (10% selective pressure)
2.3 育种值分析
根据施氏獭蛤不同性状个体育种值的前10 家系排序结果(表4), 30 日龄壳长、壳高性状家系相同率为90%, 80 日龄壳长、壳高、壳宽和体质量性状家系相同率为70%, 对应比较30 日龄和80 日龄的壳长、壳高性状家系相同率均为70%, 比较结果发现: 不同阶段和不同性状育种值排序存在差异, 但整体表现上呈现一致性, 且家系M7、M10 和M6 于不同阶段和不同性状育种值排序中均排前3, 综合育种性能突出。个体育种值和表型值的相关性显著, 基于表型值和育种值的个体留选比较结果(表5, 留种率10%), 留选个体相同率60.32%~71.43%, 根据表型值留选家系数量比根据育种值留选家系数量多0~2个, 根据选留个体的育种值统计比较, 发现两种方式选择结果存在差异, 30 日龄施氏獭蛤壳长、壳高育种值选择效率比其表型值选择分别提高27%和21%,80 日龄施氏獭蛤壳长、壳高、壳宽和体质量育种值选择效率比其表型值选择分别提高16%、14%、30%和13%, 根据育种值选择效率较高。
3 讨论
3.1 遗传参数
表型相关和遗传相关是进行间接选择主要参考依据, 遗传相关已经剔除了环境影响, 故能比表型相关更确切地反映两个性状间的相关程度。陶后全[5]等研究选育马氏珠母贝(Pinctada martensii)早期遗传参数得出其表型相关系数(0.747~0.921)高于遗传相关系数(0.528~0.746), 本研究中施氏獭蛤生长性状遗传相关系数(0.96~0.98)高于表型相关系数(0.77~0.96), 与马氏珠母贝早期性状相关性表现并不同,说明施氏獭蛤早期选择参考性状可靠性较高。
遗传力是遗传改良过程中至关重要的遗传参数,一般认为遗传力越大选择效果好。张守都[6]等研究“中科2 号”海湾扇贝(Argopecten irradians)幼虫期壳长遗传力时, 6 日龄壳长遗传力为0.58, 壳长遗传力较大证明选择育种对于“中科2 号”早期生长尚具有较大的改良潜力。王庆志[7]等估计了5~25 日龄长牡蛎(Crassostrea gigas)幼体壳高遗传力(0.161~0.771)和壳长遗传力(0.139~0.814), 遗传力表明以壳高或壳长为参数进行选育时, 均可达到改良生长性状的效果。本试验中, 30 日龄施氏獭蛤的壳长和壳高遗传力(0.31, 0.25)为中等遗传力, 80 日龄的壳长、壳高和体质量遗传力(0.35, 0.27 和0.25)也为中等遗传力, 预测施氏獭蛤早期对壳长、壳高和体质量性状进行选择时, 可获得较大的遗传进度。
养殖环境的不同使估算出的遗传力产生较大的差异[8-9], 不同遗传力估计方法也会导致估算结果的不一致。本试验繁育于秋季进行, 不同家系精卵成熟度存在较大差异, 从而对施氏獭蛤早期的遗传相关判断产生非遗传因素的影响。试验中施氏獭蛤附着幼体培育密度因存活率不同而存在客观差异, 造成家系间培育上的环境差异。另外, 试验所用亲本来源于多个半同胞家系混合建立的选育群体, 研究中将选育群体当作无亲缘关系进行系谱计算, 实际上部分亲本有可能是来自全同胞家系, 从而影响遗传力的估计准确性。全同胞效应其中包含了显性效应和母本效应, 全同胞效应在鱼类[10-11]中较小, 但在贝类幼体发育过程中, 存在较大的显性效应和母本效应[12], 根据全同胞家系估计遗传力时, 将全同胞效应当作加性效应一起估计会增大了遗传力估计值,ASReml 软件能较好的处理全同胞效应, 提高遗传力的估计的准确性, 而试验遗传力估计的标准误偏大则可能与测量样本数量偏低有关[13]。
3.2 育种值
BLUP 法可较准确地估测育种值, 并在水产动物育种广泛应用。孟乾[14]等利用BLUP 方法对长牡蛎“海大1 号”的育种值进行估计, 于50%选择压力下基于育种值家系选择效率比基于表型值提高3%~34%, 个体选择效率提高17%~28%。郑静静[15]等利用BLUP 方法评估60、105 和150 日龄3 个阶段中凡纳滨对虾(Litopenaeus vannamei)生长性状的育种值, 于10%选择压力下基于育种值比基于表型值选择效率提高20.06%、26.03%和12.01%。在本研究中, 于10%选择压力下, 试验30 日龄施氏獭蛤壳长、壳高育种值选择效率比其表型值选择分别提高27%和21%, 80 日龄施氏獭蛤壳长、壳高、壳宽和体质量育种值选择效率比其表型值选择分别提高16%、14%、30%和13%。结果表明, 在施氏獭蛤育种中利用育种值选择育种较表型值选择育种具有更高的优势。
3.3 生长性状早期选择的优劣
早期选择的主要优势是能够早期筛查和选种,从而降低选育成本。施氏獭蛤为开放式底播养殖品种, 养殖群体易混合, 提高了选择育种难度, 而其培育早期阶段因处在人工控制条件下生长, 环境因素可控性较高, 比较容易获取准确生长性状表型值并加以分析和家系选择, 可降低选育工作量。在不同选育策略下, 早期选择存在相关性优势, 如Gall 等[16]在罗非鱼(Oreochromis mossambicus)改良育种研究分析认为在通过体长的对体质量的间接选育应在幼鱼阶段进行, 因后期该遗传相关性降低, 本试验中80 d 施氏獭蛤壳长、壳高与体质量的表型相关(0.94、0.93)和遗传相关性(0.97、0.97)均较高, 这一现象有助于施氏獭蛤早期利用生长性状相关进行综合选育。贝类生长性状发育策略并非一成不变[17-18], 不同阶段性状生长呈一致性是早期选择的重要依据。本试验前期发育快的家系在早期生长中也一直领先,与彭慧婧[19]等研究整个阶段施氏獭蛤速生长家系发育情况相同, 表明对施氏獭蛤早期生长性状的选择具有较大可行性。
存活性状也是影响贝类产量的重要性状之一,早期选种可提前舍弃那些生长慢的家系或个体, 但无法对整个阶段存活性状进行预测。Langdon 等[20]对长牡蛎进行家系选育时, 发现一些家系虽然早期生长较快, 但生长至成贝时存活率较低, 闫喜武等[21]研究菲律宾蛤仔(Ruditapes philippinarum)家系早期生长发育发现稚贝期存活率趋于稳定且高于幼虫期,不同阶段存活性状表现不同。本试验中施氏獭蛤不同家系幼体附着后存活率差异较大(33%~74%), 而海上培育期存活率则相近(72%~91%), 且施氏獭蛤速生家系[19]在存活性状方面也无明显优势。尽管生长性状遗传参数和育种值对施氏獭蛤选育参考作用明显, 但在后续研究中还应综合存活性状对育种计划的影响。
猜你喜欢
中华实用诊断与治疗杂志(2022年1期)2022-08-31
中华实用诊断与治疗杂志(2022年1期)2022-08-31
养猪(2022年4期)2022-08-17
内蒙古统战理论研究(2021年1期)2021-06-09
海峡姐妹(2019年6期)2019-06-26
湘潮(上半月)(2019年2期)2019-05-22
中南林业科技大学学报(2019年4期)2019-04-08
森林工程(2018年1期)2018-05-14
华人时刊(2016年17期)2016-04-05
湖北畜牧兽医(2015年11期)2016-01-11
