
土壤放线菌资源及其应用
2021-03-11 00:37李鹤鸣
绿色科技 2021年2期
郑 丹,李鹤鸣,韩 力
(东北大学 生命科学与健康学院,辽宁 沈阳 110819)
1 引言
放线菌是一种极为重要的微生物资源,在自然界的分布十分广泛,因其具有复杂的次级代谢系统,可以产生诸多化学结构新颖、生物活性显著的次级代谢产物,所以对农业、医药、食品、工业、环境污染治理等生产生活的各个方面的发展均有重要意义,值得进一步深入探索研究。
土壤由固、液、气三种物质共同组成,并且相互联系又相互制约,形成了高度异质的环境,这些物质为放线菌提供了适宜的生长条件,是放线菌栖息的主要场所,无论是数量还是种类,都极为丰富,甚至在一些极端恶劣的土壤环境中也有放线菌的存在[1]。这些来自土壤的放线菌为研制和开发具有高效抗菌活性的抗生素提供了重要的资源,一些极端环境土壤中生存的放线菌还具有特殊的生物学特性和代谢途径,值得对从中开发药物、特殊酶进行研究。另外,放线菌还可以降解纤维素等有机化合物,在环境污染的生物治理中同样发挥重要作用。因此,研究开发土壤放线菌资源具有极高的科学价值、经济价值和社会价值。
本文将对普通土壤(森林土壤、草地土壤、农耕地土壤)和极端环境土壤(高盐环境、极端pH值、干旱环境、潮湿环境、极端温度、高辐射环境)几种不同生境下的放线菌资源及其在各领域的应用情况加以介绍。
2 普通土壤放线菌资源
2.1 森林土壤放线菌资源
森林植被类型丰富多样,人为干扰较少,是动植物良好的栖息繁衍地,为各类生物提供了多样化的生境。森林生态系统中的土壤可分为植物根际土壤、苔藓土壤、根区根表土壤。
李建康等人研究了秦岭山区人工纯林土壤微生物的群落特征,其结果表明根际土壤放线菌数量总是比非根际土壤放线菌数量多,主要是由于根际土壤有机质含量高,分解较快,土壤疏松,水分条件好,适合微生物的生长繁殖。而非根际土壤中所含根系较少,土层比较坚实,透气性也不如根际土壤,微生物不能充分生长和繁殖,因此微生物数量相对根际土较少。在阔叶林中放线菌数量明显高于针叶林,主要原因是放线菌最适合生长在中性或偏碱性的土壤中,而阔叶林地的土壤pH值条件适宜放线菌活动,使放线菌能够很好地生长。另外,植被类型不同、坡向不同时,土壤放线菌数量均表现为随海拔升高先升高又降低的趋势。同时,坡向对放线菌的分离效果和土壤养分的影响也很大,其原因可能是温度、光照、降水等因素综合影响土壤养分含量,导致南北坡存在较大差异[2]。
王东胜等人对秦岭主峰太白山北坡5种生境中微生物区系及拮抗放线菌资源的研究表明,56%的可培养放线菌均具有抗菌活性。虽然不同生境中均蕴藏着丰富的拮抗性放线菌资源,但其数量却存在差异,即使是相同类型的生境,处于不同海拔,其蕴藏量也不同。阔叶林根域拮抗性放线菌数量多于针叶林,拮抗性放线菌资源在低海拔土壤中的蕴藏量大于高海拔[3]。
2.2 草地土壤放线菌资源
我国北方主要草地类型分为五种:平原草甸、典型草原、草甸草原、高寒草甸和荒漠草原,土壤类型复杂多样,地带性分布明显。平原草甸的土壤因地势低平排水不畅,地下水含较高浓度可溶性盐,以及毁草开荒、草场过度放牧等人类不当活动,容易出现盐碱化;草甸草原的土壤因不合理开垦容易出现土壤风蚀化;典型草原的土壤较为肥沃;荒漠草原的土壤含水量低,容易出现水蚀和风蚀;高寒草甸的土壤在地貌、环境、地形等因素的影响下,土层较薄,温度较低,但土壤中含有丰富的有机质[4]。
赵璇在对中国北方主要草地类型土壤放线菌的研究中,分析指出了区域尺度下放线菌多样性和群落结构的变化规律及主要的环境驱动因子。研究表明:对于不同草地类型的土壤放线菌,其多样性及群落结构有很大差异。其中,平原草甸的放线菌多样性要显著低于其他四种草地类型;土壤放线菌多样性主要与土壤pH值有关,放线菌门下各纲的相对多度由土壤理化指标控制[5]。
2.3 农耕地土壤放线菌资源
农耕地土壤可分为灌溉耕地和雨养耕地,放线菌群落结构与草地土壤相似[6],其中灌溉耕地土壤的生物量、土壤肥力、有机无机物质量、土壤微生物量碳(MBC)均高于雨养耕地[7]。
有较多报道认为,耕作可造成土壤团菌体的分裂和表层土壤中有机质的损耗,相关研究指出,耕作对土壤放线菌的影响与土壤有机碳含量及酸碱性有关,另外,土壤耕作可能造成土壤结构的恶化,对于酸性和有机碳含量低的土壤,耕作可显著降低放线菌数量和多样性[8,9]。
3 极端环境土壤放线菌资源
极端环境土壤是指相对于人类和其它高等生物所能承受的最大范围而言,一系列对于一般微生物生长比较严酷的土壤条件,例如化学性质的极端:高盐、高酸、高碱、及高辐射等环境,以及物理条件的极端,如高温、低温等。某些放线菌能在极端环境中生长,在生存和适应的过程中形成了与正常环境土壤中生长的放线菌有很大差异的生理和代谢功能,如生物膜结构、酶的分子结构、生长特性、营养需求、基因的表达和调控机制等。
3.1 高盐环境土壤放线菌资源
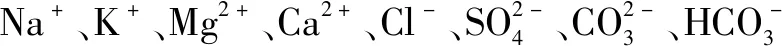
嗜盐放线菌是非常重要的一类极端微生物资源,其主要生境是富盐的碱土和海水等含盐环境。理论上,嗜盐放线菌应具有特殊的生理机制,因此,已成为近年来放线菌研究的热点之一。关统伟等对新疆塔里木盆地可培养嗜盐放线菌系统发育多样性进行研究,结果表明该地高盐环境中潜藏着丰富的新型嗜盐放线菌资源[11]。不同嗜盐放线菌对盐的耐受程度、需求程度不同,Kushner根据它们所需最适生长盐浓度的不同,将其划分为非嗜盐菌、耐盐菌、弱嗜盐菌、中等嗜盐菌、以及极端嗜盐菌[12]。尽管对于嗜盐放线菌的研究始于1975年,之后不断有新物种从含盐环境中分离出来,但由于嗜盐放线菌难以培养、营养需求特殊,相关的研究工作进展较缓慢,尤其是对陆源嗜盐放线菌的研究主要集中在物种的多样性方面,而在次生代谢产物方面的研究成果则报道较少[13]。
3.2 碱性土壤放线菌资源
在高碱性环境中能够生存的放线菌称为嗜碱放线菌。它们主要分布于pH值为10左右的高碱性极端环境中,包括盐碱土壤、碳酸盐荒漠、碱湖、碱性泉水、盐池等,如东非大裂谷的碱湖、我国新疆的盐碱湖。这些环境的碱性物质主要来源于碳酸盐。在碱性沙漠土壤、含有腐烂有机物的碱性土壤或人为造成的碱性环境中也存在嗜碱放线菌菌株。
嗜碱放线菌是极端环境中的一类特殊放线菌,具有独特的生理生化特征、基因信息及代谢途径。研究发现,虽然处于高碱环境,但嗜碱放线菌胞内pH值维持在8.0左右,依据对碱性环境中一些耐碱放线菌的研究推测主要嗜碱机制可能包括自动调节细胞质pH的机制和获得能量的机制两方面。然而,目前尚未明确真正的嗜碱放线菌的嗜碱机制,有赖于基因组学方面的进一步研究[14]。
嗜碱放线菌产生的纤维素酶可用作洗涤剂的添加剂,碱性蛋白酶则主要用于工业去污。此外,嗜碱放线菌还可以产生一些新的抗生素,如诺卡地霉素、β-内酰胺类抗生素等。
3.3 酸性土壤放线菌资源
嗜酸放线菌指广泛分布于pH值极低的酸性环境中的放线菌,尤其是酸性土壤中。它们可以分为两大类群,一类是中度嗜酸放线菌,可以在pH值为3.5或4.5至接近中性(7.5)的条件下生长,另一类是严格嗜酸放线菌,在强酸环境中生长或生存,只能生长在pH值为3.5~6.5的环境中,且利用碳源的能力强于中度嗜酸放线菌。它们可以产生蛋白酶、几丁质酶、淀粉酶等耐酸的胞外酶,参与到酸性土壤中有机物的降解循环中。此外,由于大多数真菌栖息于酸性环境,所以嗜酸放线菌也是潜在的抗真菌活性物质的生产者[15]。
在酸性和弱酸性土壤中,中度嗜酸放线菌的数量远大于严格嗜酸放线菌,其中,链霉菌是绝对优势类群。近年来,国内外的研究表明,酸性土壤中稀有的嗜酸放线菌具有丰富的生物多样性。Kim等对3株严格嗜酸放线菌进行了多相分类研究[16],发现它们在系统进化树上形成了与链霉菌属平行的单独分支,由此建立了嗜酸链霉菌新属(Streptacidiphilusgen. nov.)。
3.4 干旱环境土壤放线菌资源
在一些日照强、地表温度高、蒸发强、降水量低的干旱地区,土壤有机质含量低,环境恶劣,植被种类贫乏,生长稀疏,多为以胡杨、红柳、沙棘等为主的旱化植被,不利于微生物的生长繁殖。土壤越干旱贫瘠,放线菌的数量和种类越少,链霉菌所占比例则相对越大,可能与链霉菌的孢子化程度高、抗旱能力强、对营养的要求低有关。链霉菌的组成也相对简单,主要有六个类群(黄色类群、烬灰类群、灰褐类群、白孢类群、蓝色类群和粉红孢类群)。关统伟在对塔克拉玛干沙漠土壤中的放线菌组成和数量的研究中,分得15个属的土壤放线菌中链霉菌属占97%,其中灰褐类群、白孢类群、粉红孢类群、黄色类群约占68%[17]。
3.5 潮湿环境土壤放线菌资源
湿地沼泽或沿海滩涂土壤具有含水量高、含盐浓度较高、养分含量高及乏氧等特征,土质偏碱性,土壤中微生物以轻度耐盐菌为主,真菌数量最多,放线菌和细菌数量相对较少。放线菌主要有链霉菌属、小单胞菌属、诺卡氏菌属等,都是好氧型微生物[18]。放线菌喜欢偏碱性、有机质多的环境。潮湿土壤上层为较疏松的小块团块结构,下层则为较紧实的大块团块。随着土层加深、水分增加,土壤中氧气含量逐渐减少,厌氧微生物逐渐增多。又因土壤温度降低,有机质减少,放线菌总数量也随之减少。
水分条件是潮湿环境土壤中最关键的环境因子。在湿地系统中,水分条件一旦改变,就很容易引起盐度、pH值、土壤通气状况等其他环境因子的变化。因此,在一定范围内,放线菌的生物量、群落结构与土壤水分含量呈正相关,过高或过低都不利于土壤放线菌的生长繁殖[19]。
3.6 低温环境土壤放线菌资源
多年冻土、高寒草甸土壤等常年处在低温情况下,稳定的低温环境保存着大量的有生理代谢活性的微生物。低温微生物可分为嗜冷菌和耐冷菌2种。其中,嗜冷放线菌是可以在0 ℃及以下生长繁殖的放线菌,最适生长温度低于15 ℃,最高生长温度不超过20 ℃;耐冷放线菌在0~5 ℃可生长繁殖,最适生长温度在20 ℃以上[20]。由于低温放线菌长期生活在低温生境中,使它们形成了独特的生物学特性和适应机制,包括其特殊的基因类型、独特的质膜组成、产生具有独特功能特性的酶和抗生素等生物活性物质。同时,低温放线菌对低温环境的能量传递和物质循环也有着重要作用,值得人类探索。
3.7 地热地区土壤放线菌资源
嗜热放线菌是指放线菌目中主要生长在45~65 ℃环境的一个生态群,目前最高适应温度是70 ℃,尚未发现最适生长温度在75 ℃以上的放线菌类群[21]。适合嗜热土壤放线菌栖息的生境包括火山湖、火山口、干热河谷、温泉附近土壤等。据报道,嗜热放线菌能够产生具有重要价值的纤维素酶、淀粉酶、DNA聚合酶、脂肪酶、维生素等,也可用于处理城市垃圾、制作堆肥,在农业、医药、工业、生物技术等领域均有广泛的应用[22]。
3.8 辐射污染区土壤放线菌资源
由于某些放射性物质和放射线的存在,辐射污染区一直被认为是生命的禁区,国内外对辐射污染区微生物的研究因此甚少。我国的辐射污染区属于典型的荒漠地区,植被覆盖极少,通常此类地区中微生物种类相对单一[23]。近年来,研究人员对辐射污染区域的土壤放线菌进行了分离鉴定,发现了大量的稀有放线菌,说明存在着较为丰富的放线菌种属多样性。
4 土壤放线菌资源的应用研究
土壤放线菌能够通过菌丝体产生的胞外水解酶降解土壤中的各种有机物质,参与自然界物质循环,从而改善土壤、净化自然环境。而如今人类对土壤放线菌的应用远远超过于此,聚酮类、生物碱类等多种类型的次级代谢产物以及各种酶的产生,使放线菌已经被应用到了农业、医药行业、环境污染生物治理等人们生产生活中的各个方面。
4.1 土壤放线菌在农业当中的应用
4.1.1 农用抗生素
链霉菌产生的链霉素和土霉素可用于某些果蔬病害的生物防治,是最早用于农业的抗生素。由于这些农用抗生素使用效果良好,引起了农业研究者的广泛关注。之后,国外农业研究者们相继筛选出多氧霉素、春日霉素等防效作用较好、低毒害的新农用抗生素。尽管我国农业抗生素研究起步较晚,但仍研发出多种有效抗生素,经过几十年的发展,年产制剂量已超过8万t。其中,已实现大规模生产的农用抗生素中产值最大的品种是阿维菌素,其次是井冈霉素、赤霉素[24]。
4.1.2 重寄生
重寄生作用是指病原菌被其他微生物寄生的现象。土壤放线菌能通过重寄生侵入对农作物有害的病原菌,使菌丝发生形态上的畸变进而破裂,以达到破坏病原菌细胞的目的。放线菌侵入病原菌细胞的途径包括干扰细菌细胞壁的合成、使病原菌菌丝溶解、阻碍病原菌的蛋白质合成、使病原菌细胞质凝固等[25]。此外,重寄生作用还可以与抗生作用协同起效,即先由抗生素破坏宿主细胞细胞壁,再通过重寄生作用使放线菌侵入,从而破环病原菌细胞。农业生产中,重寄生菌作为一种控制植物病害的生物防治手段,无毒无残留、对非靶标微生物无害、防病持效期长,可用于防治“慢锈”“慢白粉”或经济价值高的作物病害[26]。
4.1.3 酶及酶抑制剂
一些土壤放线菌能够产生几丁质酶、纤维素酶、β-1,3-葡聚糖酶、β-葡糖苷酶等,可水解高等真菌、卵菌纲真菌等的细胞壁,从而破坏病原菌菌体,使其质壁分离或细胞破裂,进而死亡。如目前在农业上,可直接利用几丁质酶产生菌防治植物真菌病害[27]。此外,有些放线菌还可以通过产生几丁质酶抑制剂来抑制真菌细胞壁的合成,使病原菌死亡[28]。
放线菌作为纤维素酶的主要产生菌,在生物燃料生产、食品饲料工业等领域应用广泛,可用于处理农作物收获后的秸秆,既节约成本,又保护环境。同时,在饲料工业中,由于饲料中存在大多数动物无法分解消化的纤维素,将放线菌产生的纤维素酶加入到饲料之中,可以使牲畜将纤维素转化为更易消化吸收的葡萄糖,促进它们的良好生长[29]。放线菌产纤维素酶相较于其他纤维素酶有耐碱、耐热的特点,例如侯晓娟等从土壤中分离得到一株产碱性纤维素酶放线菌,该酶可在pH值为11的条件下保持60%以上的酶活性;宋波等筛选分离得到的产纤维素酶放线菌,其纤维素酶在70 ℃时仍具有较高活性[30]。
4.2 土壤放线菌在医药中的应用
4.2.1 抗肿瘤抗生素
放线菌次级代谢产物是开发抗肿瘤抗生素的重要资源,目前临床上使用的天然来源抗肿瘤抗生素中,超过2/3是由放线菌产生的。来源于放线菌的抗肿瘤活性物质,除了化学结构多样、生物活性显著,还有方便大规模生产、污染小等优势。
放线菌可以产生多种类别的抗生素,如蒽环类、大环内酯类、烯二炔类、糖肽类、醌类等。1975年在太平洋Easler岛土壤样品放线菌次生代谢产物中最初发现的35元环大环内酯类抗生素西罗莫司(也称雷帕霉素),在发挥抗肿瘤作用时,与目的靶蛋白结合,可抑制细胞G1至S期的进程[31]。在临床上用于乳腺癌、前列腺癌等的治疗,也有研究表明,该药有治疗子宫内膜癌的前景?[32]。在临床前肿瘤导向治疗中,十元环烯二炔类抗生素卡利奇霉素(CLM)疗效显著[33]。该抗生素由湖北武汉蛇山土壤中筛选到的稀有放线菌小单孢菌的发酵液分离得到,临床应用中以单克隆抗体为载体,具有很强的靶向性,对乳腺癌、卵巢癌等均有很好的疗效。糖肽类化合物平阳霉素与博宁霉素等同属博来霉素族抗生素,是从浙江省平阳县土壤放线菌培养物中分离得到的,可通过与DNA结合,引起DNA断裂,使DNA复制受阻而发挥其抗肿瘤作用。在临床,此类抗生素主要与其他药物联合使用来治疗恶性淋巴瘤、食管癌、鼻咽癌、结肠癌、肺癌等[34]。以丝裂霉素(Mitomycin)为代表的一类普遍使用的醌类抗肿瘤药物,多由土壤放线菌发酵产物中分离获得,抗肿瘤谱较广,结构中包含乙撑亚胺和氨甲酰脂基团,通过与DNA双链联结,抑制DNA复制,从而抑制肿瘤发展[35~37]。临床使用中结合脂质体和纳米制剂,可以提高药物的治疗指数。同样来源于土壤放线菌的多肽类化合物放线菌素D(Actinomycin D)由两个五肽环和一个平面结构的吩噁嗪酮组成,可与DNA 结合,抑制DNA复制和RNA的转录,进而抑制肿瘤生长,是临床上常用的抗肿瘤抗生素,抗肿瘤谱窄,可与其他化疗药物或放疗联合使用[38,39]。
4.2.2 药物先导化合物
土壤放线菌次生代谢产物的生物合成途径和酶反应系统具有多样性,活性基团可以与结构骨架无限组合,从而产生了一些化学结构新颖、生物活性显著的化合物,为新药研究与开发提供了丰富的模式结构和药物前体,如灵菌素、海鞘素B等[40],是新药和药物先导化合物开发的宝贵资源。
4.2.3 酶抑制剂
引起各种非感染性疾病的原因之一是人体内某些代谢酶的紊乱,因此可以利用酶抑制剂开发相应的药物。酶抑制剂可以用于治疗高血脂、糖尿病等疾病,还具有调节免疫的功效,代表药物有不可逆的β-内酰胺酶抑制剂克拉维酸、细胞表面氨肽酶抑制剂乌苯美司、假氨基糖类α-糖苷酶抑制剂阿卡波糖等。
4.2.4 免疫抑制剂
免疫抑制剂是一类通过调节机体免疫功能抑制机体异常免疫反应的药物,在临床上用于治疗移植排异或红斑狼疮等自身免疫性疾病。如从土壤放线菌发酵产物中分离出的他克莫司是一种有强大免疫抑制作用的23元环大环内酯类免疫抑制剂,临床主要用于器官移植[41]。另一种代表性药物西罗莫司,既是抗肿瘤抗生素,又具有免疫抑制活性,表现出比他克莫司更有效的免疫抑制作用,不良反应也相对较轻[31]。
4.3 土壤放线菌在环境污染治理中的应用
4.3.1 土壤重金属污染及生物修复
随着人类对于重金属的使用日趋广泛,重金属污染越来越严重,进而导致土壤质量的下降。传统的物理、化学治理措施导致的二次污染、能耗过大等问题已经无法满足当今的治理要求,而微生物修复技术成本低廉且修复彻底,因此,对重金属污染土壤的生物治理已经成为国内外的研究热点。其中,放线菌是广泛分布于土壤的优势类群,而且对于不同重金属具有吸附作用,还可以与植物共同作用,促进植物对金属离子的富集,是良好的生物吸附材料。其对重金属污染土壤的修复主要依靠自身的积累作用、溶解作用和氧化还原作用[42]。
4.3.2 土壤放线菌降解纤维素
由于大部分有机固体废物的主要成分是纤维,同时纤维素也是一种重要的可再生资源,因此无论是解决能源问题还是防止环境污染,对纤维素的生物降解的研究都有重要意义。目前,国内外对能够分解纤维素的放线菌研究报道尚少[43],但由于放线菌对于高温和各种酸碱环境等不利条件的耐受程度比其他微生物更高,所以放线菌在特殊条件下对分解纤维素有着重要作用[44]。
4.3.3 土壤放线菌菌群处理污水
在废水中,土壤放线菌可以与其他菌群组成一种有效的微生物群,活化后能快速生长繁殖,共生增殖并发挥协同作用,分解有机物,产生抗氧化物质,形成一个稳定复杂的生态系统,还能抑制有害微生物的生长繁殖,抑制硫、氮和其他物质产生的恶臭气味,激活具有净水功能的微生物、原生动物和水生植物,最终通过这些生物的综合作用达到净水处理的目的[45]。
5 总结与展望
(1) 国内外对土壤放线菌资源的研究按其生境不同可包括普通土壤(森林土壤、草地土壤、农耕地土壤)放线菌和极端环境(高盐环境、极端pH、干旱环境、潮湿环境、极端温度、高辐射环境)土壤放线菌。文中介绍了这些不同环境下的土壤概况、生态背景、理化性质、养分特征等,以及土壤放线菌的种群多样性、生态分布规律、不同因素对土壤放线菌的影响等。
(2)土壤放线菌在生产生活的各方面应用都十分广泛。在农业中,土壤放线菌的重寄生作用、酶及抑制剂的降解作用、对纤维素的分解作用等都有重要意义。在医药行业中,土壤放线菌丰富的次级代谢产物可用于抗肿瘤抗生素、酶抑制剂、免疫抑制剂等的开发生产。在环境污染治理中,土壤放线菌亦可发挥其生物修复作用。因此,开发与研究土壤放线菌资源具有极高的科学价值、经济价值和社会价值。
(3)目前在土壤放线菌资源的研究中还存在一些问题。随着对土壤放线菌研究开发的不断深入,目前对普通土壤环境中放线菌的研究开发愈发困难,发现新菌的概率越来越低,需要进一步从一些特殊生境中分离筛选新的放线菌。同时土壤放线菌分离筛选工作也陷入瓶颈,需要科研工作者结合化学、分子生物学等学科的相关技术与理论,进一步积累和不断创新,如不断开发更有效的预处理方法、抑制剂和分离培养基等。
(4)由于土壤放线菌次生代谢产物的有效成分复杂、含量低,而且对其化学组成及结构的研究过程耗溶剂、耗能、耗时、提取率低,大量的重复性实验过程使得发现新药物先导化合物的几率不断降低。应当不断提高选择性分离技术、UPLC-Q-TOF-MS去重复化技术、快速鉴定技术等,以提高从土壤放线菌中开发新化合物的效率。
猜你喜欢
现代园艺(2022年19期)2022-09-21
中老年保健(2022年5期)2022-08-24
小读者(2021年19期)2021-11-07
红领巾·探索(2021年2期)2021-08-26
学生天地(2020年14期)2020-08-25
军事文摘·科学少年(2020年2期)2020-03-19
当代陕西(2019年11期)2019-06-24
作文评点报·中考版(2019年4期)2019-02-18
儿童故事画报·自然探秘(2019年12期)2019-02-02
课外语文·中(2018年10期)2018-11-16
