
湖羊PLAG1基因5′调控区多态性及其与早期体重的关联分析
2021-02-27 04:57郭潇潇李隐侠孟春花王慧利仲跻峰曹少先
畜牧兽医学报 2021年2期
郭潇潇,李隐侠,王 悦,张 含,张 俊,钱 勇,孟春花,王慧利,仲跻峰,曹少先,3*
(1.南京农业大学动物科技学院,南京 210095;2.江苏省农业科学院畜牧研究所,南京 210014;3.江苏省农业种质资源保护与利用平台,南京 210014)
湖羊是我国著名的地方绵羊品种,具有性成熟早、产多羔、耐湿热等优点,但其生长速度仍有待提高。在湖羊的生长发育过程中,初生体重和断奶体重与后期生长高度相关,同时,初生体重还与羔羊存活力关系密切,因此开展初生体重和断奶体重与基因标记的关联研究,对于湖羊本品种选育或以湖羊为育种素材的肉羊新品种培育具有十分重要的意义。
多形性腺瘤基因1(pleomorphic adenoma gene 1,PLAG1)是多形性腺瘤基因家族成员之一,PLAG1基因在结构上包括7个C2H2锌指和1个富含丝氨酸的转录激活区[1],Kas等[2]在研究唾液腺多形性腺瘤时通过位置克隆首次发现了PLAG1基因,后来陆续报道PLAG1基因与急性髓细胞白血病等癌症有关[3-5]。近年来研究发现,PLAG1基因可调控动物的生长发育且在胚胎期表达量高,出生后表达量降低甚至低于检测限[6-8]。研究表明,PLAG1敲除导致小鼠生长迟缓和生育能力下降[9-11]。Abi Habib等[12]研究发现,HMGA2-PLAG1-IGF2通路的遗传缺陷可导致胎儿及出生后生长受限。Karim等[13]发现,14号染色体约25 Mb 位置的QTL对身高和体重有重要影响,Fink等[14]对该区域进行QTL(eQTL)定位发现,QTL区域包含的多个基因中只有PLAG1差异表达,且只有PLAG1与奶牛生长和体重的QTLs有相同的关联特征。人类和家畜的全基因组关联研究也证实了PLAG1在生长中的作用[15-20]。Zhong等[21]研究显示,PLAG1基因内或附近的单核苷酸多态性与牛的体型等经济性状有关。Li等[22]通过对牛PLAG1的多态性进行分析发现,缺失位点(g.3747444-3747462 del GAACACACAGCCCACAAGG)和替换位点(g.44966G>A)对中国牛体高、腰围、臀长、体重等生长性状有显著影响,Guo等[23]研究表明,PLAG1是影响猪肢骨长的候选基因。但湖羊PLAG1基因多态性及其与生长性能的关联研究尚未见报道。
本试验以湖羊为研究对象,对PLAG1 5′UTR和基因5′调控区进行克隆,筛选5′调控区SNPs位点,并与初生体重、断奶体重进行关联分析,为加快以湖羊为育种素材的肉羊新品种培育提供候选分子标记。
1 材料与方法
1.1 材料与试剂
456只断奶湖羊及其初生体重、断奶体重等数据资料由江苏西来原生态农业有限公司提供,试验湖羊出生于同一季度、相同饲养环境,营养水平和饲养管理一致,采集湖羊耳组织,放入冰盒带回实验室,-20 ℃冰箱保存备用。湖羊胎羊肌肉组织采自江苏省农业科学院六合基地种羊场,采集后立即投入液氮罐,带回实验室后转移到-80 ℃冰箱保存。
2×TaqMaster Mix、HiScript Ⅱ Reverse Transcriptase试剂盒均购自南京诺唯赞生物科技有限公司,DNA Marker、SMARTerRACE 5′/3′ Kit User Manual、pMD19-T均购自北京宝日医生物技术有限公司,Goldview I型核酸染色剂购自南京寿德生物科技有限公司,动物组织总RNA提取试剂盒购自北京天根生化科技有限公司,DNA凝胶回收试剂盒、TreliefTM5α Chemically Competent Cell均购自南京擎科生物科技有限公司。
1.2 DNA和RNA提取及检测
参照动物组织总RNA提取试剂盒说明书提取湖羊胎羊肌肉组织总RNA。酚/氯仿抽提法提取耳组织样DNA。紫外分光光度计检测DNA/RNA纯度和浓度,琼脂糖凝胶电泳检测其完整性。
1.3 引物设计与合成
NCBI网站查找绵羊PLAG1基因(NC_019466.2)及mRNA预测序列(XM_012183720.1),利用Primer 5.0设计5′UTR、5′RACE和5′调控区特异性引物,送至上海捷瑞公司合成。引物信息见表1。
1.4 PCR扩增
PCR反应体系为25 μL:2×TaqMaster Mix 12.5 μL,上、下游引物各1 μL,DNA模板1 μL,添加ddH2O至25 μL。PCR扩增程序:95 ℃预变性5 min;95 ℃变性30 s,退火30 s,72 ℃延伸1 kb·min-1(根据表1引物PCR扩增片段长度确定延伸时间),35个循环;72 ℃延伸5 min。引物序列及退火温度见表1,PCR产物经琼脂糖凝胶电泳、切胶,送南京擎科生物公司测序。
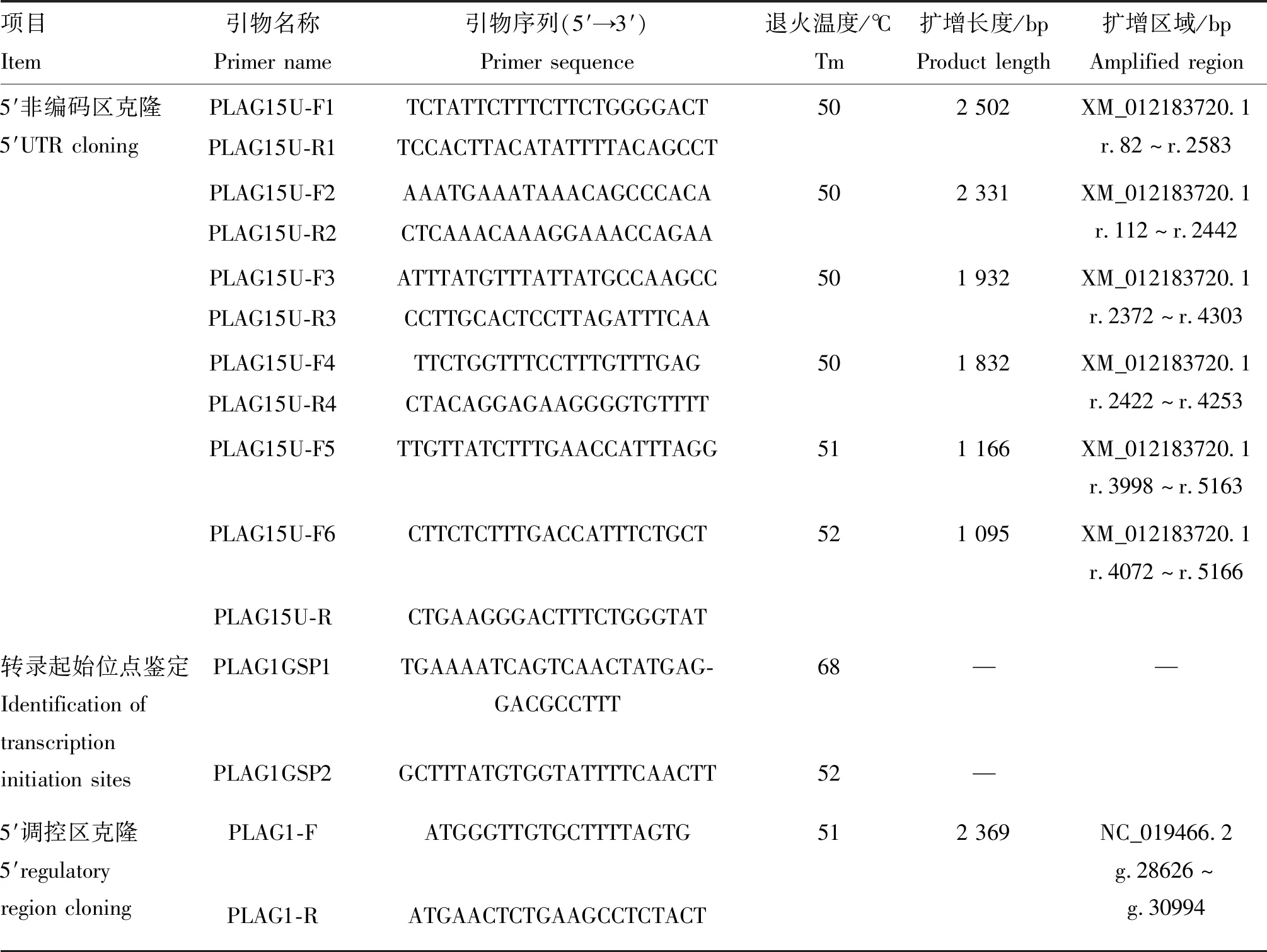
表1 特异性引物信息
1.5 5′RACE克隆
1.5.1 合成cDNA第一链 准备Buffer Mix:5× First-stand Buffer 4.0 μL、DTT 0.5 μL、dNTPs 1.0 μL 离心混匀,置冰上备用。将肌肉组织RNA模板1 μg(1.0~10 μL)、5′-CDS Primer A 1.0 μL、sterile H2O 0~9 μL加入离心管中混匀,PCR仪中72 ℃反应3 min,42 ℃反应2 min,用微型离心机离心10 s,加入1 μL SMARTerⅡ A Oligonucleotide。加入Buffer Mix 5.5 μL、RNase Inhibit 0.5 μL、SMARTScribe Reverse Transcriptase 2.0 μL混合,混匀,PCR仪中42 ℃反应90 min,70 ℃反应10 min。作为5′RACE Ready cDNA使用。
1.5.2 扩增5′非编码区末端序列 采用巢式PCR,第一轮PCR:准备A、B、C共3个PCR管,加入PCR-Grade H2O 15.5 μL、2×SeqAmp Buffer 25.0 μL、SeqAmp DNA Polymerase 1.0 μL离心混匀,分别加入5′RACE Ready cDNA 2.5 μL。A组加入10×UPM 5 μL;B组加入PLAG1GSP1 1 μL;C组加入PLAG1GSP1 1 μL、10×UPM 5 μL。每组补充ddH2O至50 μL,离心混匀。PCR反应程序:94 ℃ 30 s,72 ℃ 3 min,5个循环;94 ℃ 30 s,70 ℃ 30 s,72 ℃ 3 min,5个循环;94 ℃ 30 s,68 ℃ 30 s,72 ℃ 3 min,25个循环。反应结束后,3组分别取5 μL扩增产物进行琼脂糖凝胶电泳,观察条带是否特异及拖带。
第二轮PCR:准备D、E、F共3个PCR管,分别加入2×TaqMaster Mix 12.5 μL、C管加入反应原液1 μL。D组加入Universal Primer Short 1 μL;E组加入PLAG1GSP2 1 μL;F组加入PLAG1GSP2 1 μL、Universal Primer Short 1 μL。每组补充ddH2O至25 μL,混匀。PCR扩增程序:95 ℃预变性5 min;95 ℃变性30 s,52 ℃退火30 s,72 ℃延伸3 min,30个循环;72 ℃最终延伸5 min。反应结束后,3组分别进行琼脂糖凝胶电泳,切胶回收特异性条带。
1.5.3 TA克隆 切胶回收的DNA与pMD19-T载体连接,进行转化、摇菌,将菌液送至南京擎科公司测序。
1.6 基因分型
根据测序结果,使用Chromas进行分析,查找SNP位点。统计计算基因型频率及等位基因频率。
1.7 数据分析
PIC-CALC程序计算多态信息含量,HAPLOVIEW软件进行SNPs连锁分析,SPSS 18.0软件中One-Way ANOVA方法统计分析不同基因型、单倍型与初生体重、断奶体重的差异。数值以“平均数±标准差”表示,P<0.05表示差异显著,P<0.01表示差异极显著。
2 结 果
2.1 PLAG1预测转录本5′UTR的验证
以NCBI数据库中绵羊PLAG1预测的转录本XM_012183720.1序列为基础,设计引物,以湖羊肌肉组织cDNA为模板,分段巢式扩增,验证预测的5′UTR序列是否正确。结果显示,第一轮无目的条带,第二轮扩增出的条带与预测目的条带大小一致(图1),PCR产物测序拼接总长度为4 968 bp,并发现与预测序列XM_012183720.1一致,表明预测序列确实为PLAG1 5′UTR序列,但转录起始位点和5′UTR 的5′末端序列仍需通过5′RACE确定。
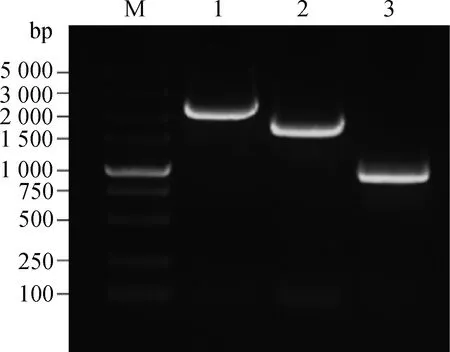
上图为第二轮巢式PCR。M.DNA相对分子质量标准;1~3.依次为5′非翻译区(5′UTR)r.112~r.2442、r.2422~r.4253、r.4072~r.5166的扩增产物
2.2 PLAG1基因转录起始位点鉴定
利用5′RACE技术,扩增PLAG1 5′UTR的5′末端序列,获得的特异性条带分别经TA克隆、测序比对,结果显示,获得5种PLAG1 5′UTR的5′末端序列,且转录起始位点均位于PLAG1基因g.30535位点(图2)。

箭头所示为PLAG1转录本转录起始位点
2.3 PLAG1基因5′调控区扩增
根据NCBI数据库PLAG1基因序列(NC_019466.2),从已经鉴定的PLAG1基因转录起始位点g.30535向前扩增5′调控区,分别在g.28626、g.30994 位点设计特异性引物扩增大小为2 369 bp的目的片段。如图3所示,表明成功扩增PLAG1基因5′调控区。
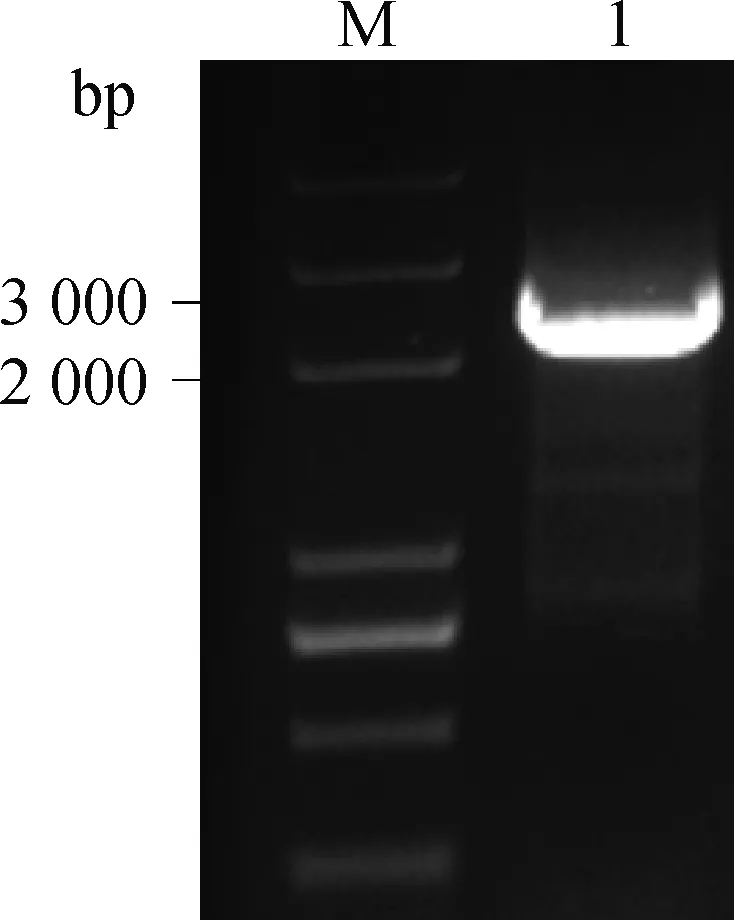
图3 PLAG1 基因5′调控区的PCR扩增
2.4 PLAG1基因5′调控区SNPs筛选
通过PCR产物测序,在湖羊群体中检测到PLAG1基因5′调控区共有15个突变位点,均具有3种基因型。测序峰图如图4所示,分别为g.28888A>T、g.28901G>T、g.28913T>A、g.29717G> C、g.29724A>C、g.29725G>A、g.29768T> C、g.29771A>G、g.30141T>A、g.30144A> G、g.30691C>A、g.30694A>T、g.30737T> C、g.30802C>T、g.30825A>G。
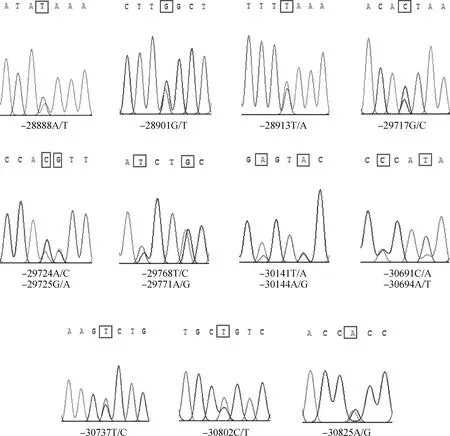
图4 PLAG1 SNPs位点杂合型测序图
2.5 PLAG1基因5′调控区群体遗传学特征
2.5.1PLAG1基因5′调控区SNPs基因分型 通过PCR产物测序,456只湖羊群体中PLAG1基因5′调控区共发现15个突变位点,15个SNPs位点的基因型频率、等位基因频率及多态信息含量见表2。结果表明,15个SNPs位点在湖羊群体中均有3种 基因型,均处于Hardy-Weinberg平衡状态(P>0.05);g.28888A>T、g.28901G>T、g.29717G>C、g.29724A>C、g.29725G>A、g.29768T>C、g.29771A> G、g.30141T>A、g.30144A>G、g.30691C> A、g.30694A>T、g.30802C>T、g.30825A> G多态信息含量(PIC)为0.09或0.10,为低度多态;g.28913T>A和g.30737T>C的PIC分别为0.34和0.33,为中度多态。

表2 PLAG1基因5′调控区SNPs基因型频率和等位基因频率
2.5.2PLAG1基因5′调控区SNPs位点单倍型分析 通过连锁突变分析,发现g.28901G>T、g.29717G> C、g.29724A>C、g.29725G>A、g.29768T> C、g.29771A>G、g.30141T>A、g.30144A> G、g.30691C>A、g.30694A>T、g.30802C> T、g.30825A>G,连锁突变相关性为100%,g.28888A>T与上述除g.28901G>T外11个 位点的连锁突变相关性为97%,g.28913T>A、g.30737T> C两者连锁突变相关性为97%(图5)。单倍型分析发现,湖羊群体中具有18种单倍型,AAGGAAGGAAGGTTAATTAACCAACCCCAA单倍型频率为40.4%,AAGGTAGGAAGGTTAATTAACCAATCCCAA单倍型频率为36.0%,属于优势单倍型。其他单倍型占比低,属于劣势单倍型(表3)。

表3 PLAG1基因5′调控区不同单倍型和单倍型频率

图5 PLAG1基因5′调控区连锁分析图
2.6 PLAG1基因5′调控区SNPs突变位点与体重关联分析
2.6.1 SNPs位点不同基因型与体重关联分析 根据SNPS基因分型获得的数据,排除没有统计意义的基因型,分别将单个位点与初生体重、断奶体重进行关联分析。如表4所示,就群体初生体重而言,g.28888A>T 位点AA型显著大于AT型(P<0.05),g.28901G>T位点GG型显著大于GT型(P<0.05),g.30802C>T 位点CC型显著大于CT型(P<0.05),g.30825A>G位点AA型显著大于AG型(P<0.05)。群体断奶体重g.30737T>C位点CC型显著小于TC型(P<0.05),且CC型

表4 SNPs位点不同基因型与湖羊体重的关联分析
2.6.2 不同单倍型与体重关联分析 将PLAG1基因5′调控区突变位点进行单倍型分析,排除样本较少的单倍型,选取样本较多的5种单倍型与湖羊初生体重、断奶体重关联分析(表5)。结果发现,AAGGTAGGAAGGTTAATTAACCAATCCCAA型初生体重、断奶体重都大于其他单倍型,且初生体重与ATGTTAGCACGATCAGTAAGCAATTCCTAG型相比达到显著水平(P<0.05),断奶体重与AAGGAAGGAAGGTTAATTAACCAACCCC-AA型相比达到显著水平(P<0.05),与其他单倍型之间差异不显著(P>0.05)。

表5 不同单倍型与湖羊体重的关联分析
3 讨 论
PLAG1是多形性腺瘤基因家族重要成员之一,属于锌指转录因子,可直接靶向调控胰岛素样生长因子2(IGF2)[24-27],参与细胞增殖。小鼠PLAG1基因敲除导致生长迟缓[28],表明PLAG1对动物生长发挥重要作用。据报道,基因可通过可变剪接产生不同的5′UTR,影响mRNA稳定性、折叠及与核糖体相互作用,从而调控基因的表达[29-33]。本研究鉴定出湖羊PLAG1基因5种5′UTR序列,这些不同的5′UTR序列可能调控PLAG1基因的表达,从而影响湖羊生长发育。
全基因组关联分析研究表明,PLAG1基因内或附近的SNPs与荷斯坦牛和胡安×泽西牛杂交种的体高等经济性状有关[34]。Littlejohn等[35]研究表明,PLAG1 ss319607402(chr14 g.23232324A>G)SNP与新生小牛体重(P=1.5×10-19)、青春期前体重(P=1.9×10-5)和日增重(P=0.035)显著相关,AA型有利于牛的生长。范家萌[36]研究发现,PLAG1基因的9个SNPs位点与猪的体长性状呈显著相关。Zhou等[37]研究表明,在牛PLAG1基因的内含子1中发现了1个19 bp的插入/缺失,19 bp缺失 与体高、胯高、胸围呈显著相关(P<0.05)。这些研究表明了PLAG1基因多态性在动物体重等经济性状中的作用,可能作为动物体重选择的候选分子标记。本研究首先鉴定了PLAG1在湖羊中有5个 5′UTR且具有相同的转录起始位点,位于g.30535。根据转录起始位点位置扩增湖羊PLAG1基因5′调控区,PLAG1基因5′调控区在湖羊群体中检测到15个SNPs位点,基因分型发现15个SNPs位点在湖羊群体中均具有3种基因型。关联分析发现,g.28888A>T位点AA型、g.28901G>T位点GG型、g.30802C>T位点CC型、g.30825A>G位点AA型湖羊群体初生体重均显著大于相应的杂合型群体(P<0.05),而这些位点群体断奶体重差异均未达到显著水平,可能是由于湖羊摄乳量等后天因素或标准差较大引起的。g.30737T>C位点CC型群体断奶体重显著小于CT型(P<0.05),而CC型群体初生体重小于CT型,但差异未达到显著水平(P>0.05),推测是由于湖羊个体存在差异所致。说明g.28888A>T位点AA型、g.28901G>T位点GG型、g.30802C>T位点CC型、g.30825A>G位点AA型有利于提高湖羊的初生体重,g.30737T>C位点CT型有利于提高湖羊的断奶体重。
据报道,牛PLAG1-CHCHD7基因间区域存在双向启动子的序列变体Q(11个重复序列CCG与G连锁)和q(9个重复序列CCG与A连锁),Q启动子活性是q启动子的1.5倍,且在胎儿期QQ型胎牛肝、骨骼肌、肌肉和大脑PLAG1表达水平高于qq型[13]。研究发现,PLAG1基因5′调控区的15个突变位点在湖羊群体中连锁,说明PLAG1表达受连锁位点的调控。本试验通过对PLAG1基因5′调控区SNPs位点进行连锁和单倍型分析发现,有18种单倍型,对具有统计意义的5种单倍型与初生体重、断奶体重进行关联分析发现,AAGGTAGGAAGGTTAATTAACCAATCCCAA型初生体重、断奶体重都大于其他单倍型,且初生体重与ATGTTAGCACGATCAGTAAGCAATTCCTAG型相比达到显著水平(P<0.05),断奶体重与AAGGAAGGAAGGTTAATTAACCAACCCCAA型相比达到显著水平(P<0.05),说明AAGGTAGGAAGGTTAATTAACCAATCCCAA型有利于提高湖羊早期体重,对湖羊羔羊PLAG1表达的调控仍有待进一步研究。
4 结 论
综上所述,湖羊PLAG1基因转录起始位点位于g.30535,其5′调控区在湖羊群体中检测到15个SNPs位点且存在连锁。其中g.28888A>T、g.28901G>T、g.30802C>T、g.30825A>G位点与初生体重显著相关(P<0.05),g.30737T>C位点与断奶体重显著相关(P<0.05)。AAGGTAGGAAGGTTAATTAACCAATCCCAA单倍型与初生体重和断奶体重显著相关(P<0.05)。这些位点可以作为候选分子标记用于湖羊生长性状的辅助选择。
猜你喜欢
今日农业(2022年2期)2022-11-16
今日农业(2022年2期)2022-11-16
现代畜牧科技(2021年11期)2021-12-21
现代畜牧科技(2021年2期)2021-03-19
现代畜牧科技(2021年2期)2021-03-19
河南畜牧兽医(2020年11期)2020-01-11
广东饲料(2016年4期)2016-12-01
新闻传播(2015年6期)2015-07-18
中国医疗美容(2015年4期)2015-04-27
中国医疗美容(2015年4期)2015-04-27
