
fth1b基因敲除对斑马鱼咽齿早期矿化的影响研究
2021-02-26 05:41周春艳郑雪丹杨德琴
华西口腔医学杂志 2021年1期
周春艳 郑雪丹 杨德琴
重庆医科大学附属口腔医院牙体牙髓病科 口腔疾病与生物医学重庆市重点实验室重庆市高等教育口腔生物医学工程重点实验室,重庆401147
铁离子的动态平衡对维持有机体的正常功能起关键作用,越来越多的研究[1-6]发现铁离子失衡在许多常见疾病中发挥重要作用,例如帕金森病、阿尔兹海默症、缺血-再灌注损伤、动脉粥样硬化、肺炎及类风湿性关节炎等。因此,细胞内精密的铁离子调控机制对生物体极为重要。铁蛋白是生物体内重要的调节铁离子浓度平衡的蛋白之一,参与多种生物过程。它由24 个亚基组成的一个内径为8 nm、外径约为12 nm 的一个中空球体状蛋白[7],内部形成矿化核约可以储存4 500 个铁原子[8]。组成铁蛋白的亚基包括2 种不同类型的多肽链:重链H 和轻链L,分别由不同的基因fth 和ftl编码[9],2 种亚基具有不同功能,L 亚基有吸取铁离子贮藏于铁蛋白中的作用[10],H亚基主要发挥铁氧化酶作用[11],将二价铁离子氧化为可溶性三价铁离子储存于铁蛋白中,可以减少亚铁离子带来的超氧化造成的细胞损伤。在不同的生物及器官内,组成铁蛋白的H和L亚基比例不同而发挥不同功能来维持生物体的铁离子整体平衡[12-14]。
fth 基因的5’端非编码区序列通常很长,包含茎环结构铁响应原件(iron responsive element,IRE),铁响应原件可以与胞质中的铁调节蛋白结合[15],从而调控铁蛋白的转录翻译,进一步影响细胞铁离子浓度的稳态,导致铁离子浓度失衡而损害机体健康。目前关于IRE 序列突变的研究[15-16]显示,IRE 的突变会影响铁蛋白mRNA 的翻译效率,导致铁蛋白重链亚基蛋白减少从而造成遗传性铁过载,但是关于fth 编码基因本身的突变未见报道。
斑马鱼是分子生物学常用的模式动物之一,其产卵量大,体外受精,胚胎透明及具有终生替换咽齿等特性也使其成为牙齿发育与再生相关研究的良好模型[17]。斑马鱼咽齿的发育模式与哺乳动物具有极高的相似性[18],早期上皮增厚形成牙板,后上皮凹陷包绕间充质,硬组织矿化形成成熟的牙齿;且组织切片染色和电镜结果显示,斑马鱼咽齿的显微结构也与哺乳动物类似,由外部的类牙釉质、类牙本质和内部空腔组成[19-20]。
有研究[21]报道,铁蛋白基因在多种动物多种细胞中均有表达。在本实验中,首次发现编码铁蛋白重肽基因fth1b 在斑马鱼咽齿部位特异性表达,并利用规律成簇间隔短回文重复序列(clustered regularly interspaced short palindromic repeats,CRISPR)/Cas9基因编辑法的独特优势[22],对该基因特异性敲除,在筛选得到的可稳定遗传突变体(mutant,MUT)中检测发现,斑马鱼咽齿早期形成并无明显缺陷,包括上皮增厚及形态形成过程均正常发生,而早期矿化过程受到一定程度的影响。
1 材料和方法
1.1 实验动物
本实验所用斑马鱼均来自于西南大学分子发育实验室,按照国际动物管理委员会的规定在标准实验室环境下饲养与繁殖。实验所用斑马鱼胚胎中加入0.003% 1-苯基-2-硫脲抑制黑色素生成,便于显微镜下观察。
1.2 实验试剂及设备
rTaq DNA 聚合酶(Takara公司,日本),多聚甲醛(paraformaldehyde,PFA)(Sigma-Aldrich 公司,美国),无水甲醇、乙醇[重庆川东化工(集团)有限公司],Anti Dig-AP、显色液(Roche 公司,德国),cas9 蛋白(NEB 公司,美国),实验设备主要包括Heraeus BK-600 立式恒温胚胎培养箱、聚合酶链式反应(polymerase chain reaction,PCR)仪及水浴锅(Eppendorf公司,德国),体视显微镜(Leica 公司,德国),共聚焦显微镜(Carl Zeiss 公司,德国)。PCR 所用全部引物及DNA 测序均由北京六合华大基因科技有限公司完成。
1.3 整胚原位杂交
收集野生型(wild-type,WT)斑马鱼56、72、96、120 hpf [受精后小时(hours post fertilization,hpf)]胚胎,4%PFA 固定过夜后无水甲醇脱水并于-30 ℃储存。按文献[23-24]描述进行原位杂交,检测基因fth1b、dlx2b的表达情况:1)将胚胎依次于75%甲醇/25%磷酸盐缓冲液含吐温20(phosphate buffer solution/tween-20,PBT)、50%甲醇/50%PBT、25%甲醇/75%PBT、100%PBT 中漂洗5 min 复水后,加入含蛋白酶K 的PBT 缓冲液消化20 min,然后加入4%PFA 固定30 min,PBT 缓冲液漂洗3次,每次5 min,固定后胚胎进入68.5 ℃水浴锅预杂交3 h,加入探针杂交液,水浴锅中过夜;2)回收探针杂交液,漂洗胚胎后加入封闭液室温下封闭2 h,然后加入含抗地高辛抗体的二抗反应液,4 ℃孵育过夜;3)回收二抗后漂洗胚胎,并将胚胎转移至24 孔板中避光显色,显色充分后于体式显微镜下观察拍照。
1.4 利用CRISPR/Cas9 基因编辑法特异性敲除fth1b基因
从Ensembl 数据库中下载fth1b 基因序列,根据基因序列选定合适的基因敲除靶位点,并根据靶位点设计合成正反向引物,然后通过PCR 合成能与靶位点特异性结合的导向RNA(guide RNA,gRNA),其中正向引物序列为:5,-taatacgactcactatagggtgaggcagaacttccaccgttttagagctagaaatagca-3,;反向引物序列为:5,-aaaaaaagcaccgactcggtgccac-3,。将合成的gRNA 与cas9 蛋白显微注射到单细胞WT胚胎(约300 枚)中,gRNA 可引导cas9 蛋白到靶位点处对基因fth1b 进行特异性剪切使其编码序列改变从而导致编码蛋白质功能缺失。
1.5 筛选MUT 及检测其咽齿相关基因的表达情况
经显微注射后的胚胎饲养至性成熟后裂解鱼鳞,测序检测,筛选出MUT F0,F0 与WT 斑马鱼交配得F1,当F1 生长至3 个月大小,裂解鱼鳞,测序检测,经检测确定的杂合MUT 与WT 斑马鱼交配得F2,性成熟后裂解鱼鳞测序检测,经检测确定得到的杂合MUT 自交得F3,性成熟后裂解鱼鳞检测,此时经检测确定得纯合MUT,是能稳定遗传的MUT fth1b-/-。通过筛选得到稳定遗传的MUT 后,分别收集48、96 hpf 的MUT 胚胎及WT胚胎通过原位杂交检测咽齿相关基因pitx2、dlx2b的表达情况。
1.6 检测MUT咽齿矿化情况
通过共聚焦显微镜直接观察及茜素红染色的方法对MUT 发育过程中咽齿矿化情况进行检测。茜素红染色:分别收集野生型与MUT 120 hpf 胚胎,4%PFA 4 ℃固定过夜后PBT缓冲液漂洗5 min,50%乙醇常温下脱水0.5 h后加入1 mL PBT和20 μL 0.05%茜素红染色液过夜染色,然后漂白液室温下漂白20 min,洗去漂白液,于50%甘油/0.25%氢氧化钾中保存及图片采集。共聚焦显微镜:在2.5 倍体式显微镜下用镊子剥离120 hpf 野生型及MUT 胚胎的心脏等组织,充分暴露咽齿,琼脂糖凝胶固定后于共聚焦显微镜下直接观察并采集图片。
2 结果
2.1 基因dlx2b、fth1b 在斑马鱼咽齿中的特异性表达情况
对dlx2b、fth1b 的表达情况检测结果显示:在56 hpf 时,咽齿部位无dlx2b 表达,在72~120 hpf各时期,dlx2b 特异性表达于斑马鱼咽齿(图1),同时,在56 hpf时,咽齿部位无fth1b表达,在72~120 hpf,fth1b 在斑马鱼咽齿表达且在全身其他部位无表达(图2),这表明fth1b 基因与目前公认斑马鱼咽齿标记基因dlx2b 表达情况一致,特异性表达于斑马鱼咽齿,提示此基因可能在斑马鱼咽齿的发育过程中发挥作用。
2.2 fth1b基因敲除MUT构建结果
经cas9 蛋白注射后,fth1b 基因被成功突变(图3)。如图3 所示,基因敲除导致在cas9 靶位点处1个碱基缺失,造成移码突变,从而导致编码序列改变,提前终止,编码氨基酸由177 个减少为16个。
2.3 fth1b-/-中斑马鱼咽齿相关基因pitx2 及dlx2b 表达情况
在fth1b-/-MUT中,对pitx2、dlx2b的表达情况检测结果显示,48 hpf时pitx2的表达与WT斑马鱼并无较大差别,均在咽齿上皮处有较强表达(图4);在96 hpf时,dlx2b的表达范围、强度与WT斑马鱼无较大差别(图4)。此结果提示,在fth1b-/-MUT中,斑马鱼咽齿的起始及上皮增厚形态形成等过程并未受到影响。
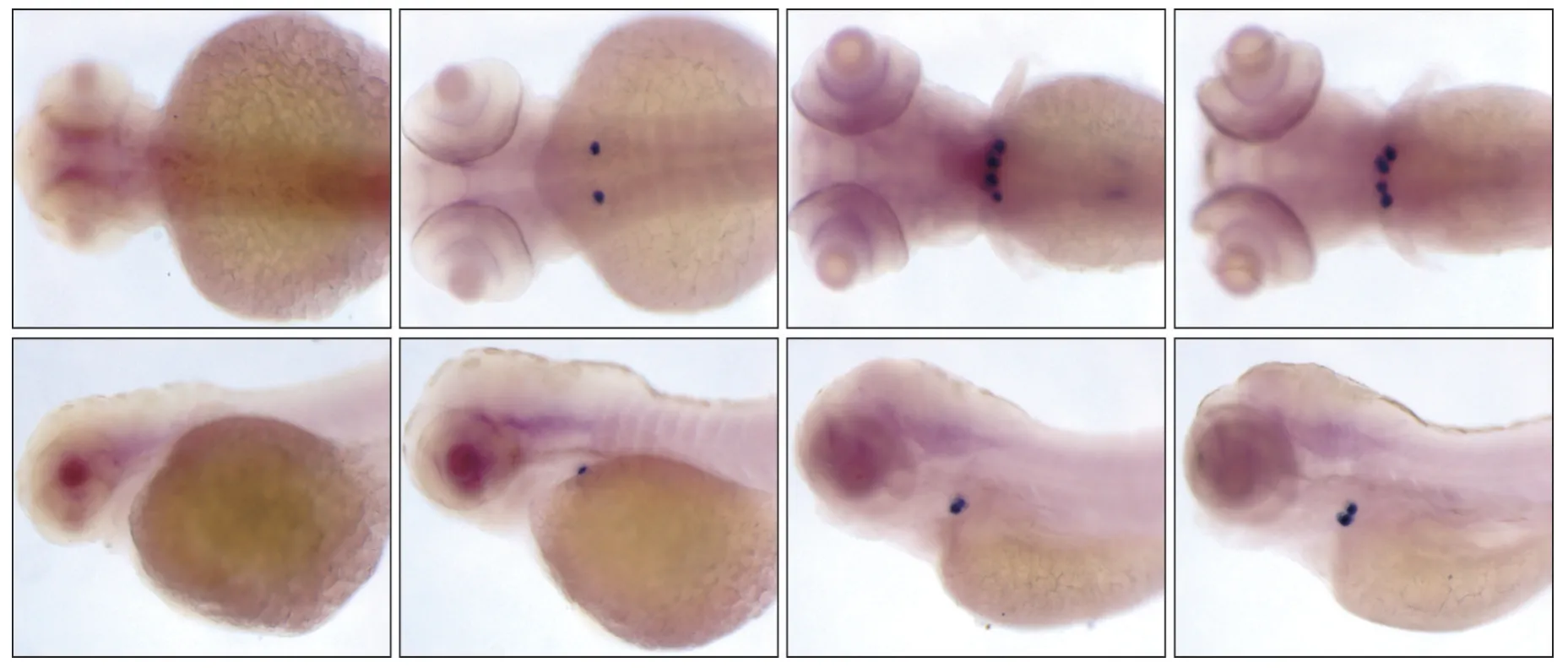
图2 56~120 hpf时基因fth1b在斑马鱼咽齿的表达情况Fig 2 The expression of gene fth1b in 56-120 hpf zebrafish embryos
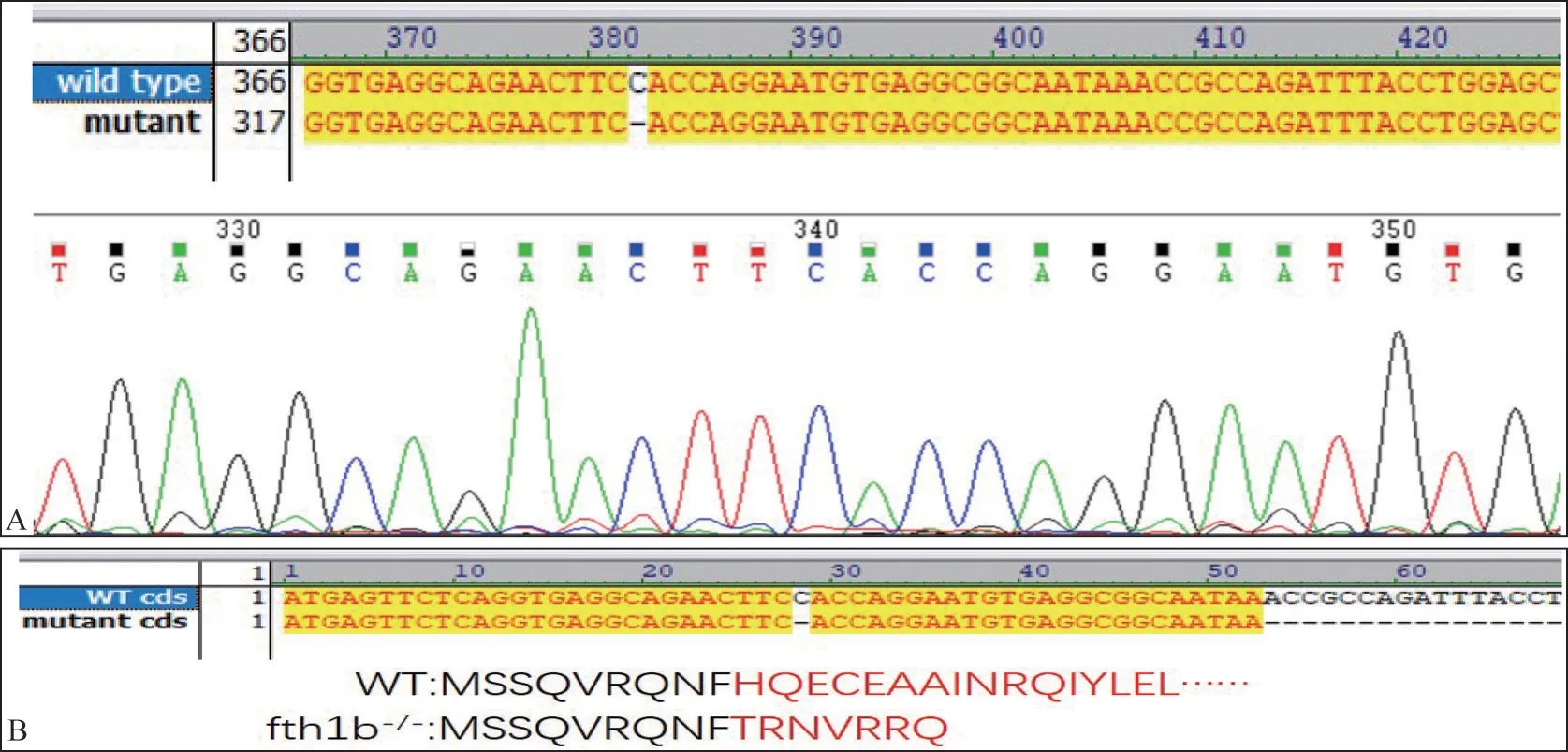
图3 WT斑马鱼及fth1b MUT的基因序列(A)和蛋白编码序列(B)Fig 3 The gene sequence(A)and protein coding sequence(B)of WT zebrafish and mutant fth1b
2.4 fth1b-/-斑马鱼咽齿矿化检测结果
通过茜素红染色结果可以观察到,在120 hpf时,WT 斑马鱼腹侧第四位置第一颗咽齿4V1已完全矿化,腹侧第三、五位置第一颗咽齿3V1及5V1处于矿化初期,仅牙尖部可见少量矿化信号,而在MUT 中,仅可见4V1矿化,3V1及5V1处未见矿化信号(图5);共聚焦显微镜下观察,MUT 斑马鱼中,咽齿的数量及形态均与野生型斑马鱼相同,矿化稍弱(图5)。因此,笔者认为在fth1b-/-MUT中,牙齿的早期发育及形态形成均正常,而早期矿化受到影响,即fth1b 基因在斑马鱼咽齿的矿化过程中发挥作用。
3 讨论
大量研究已证实铁离子浓度平衡对机体保持健康极为重要,其对血液系统的影响会造成缺铁性贫血等疾病,并且过量铁离子能通过氧化应激作用导致细胞程序性死亡,而铁蛋白作为铁离子浓度调节的重要蛋白,能够作为抗氧化剂对细胞起保护作用,并且有研究报道铁蛋白是一种有前途的药物载体,可用于肿瘤的靶向治疗。尽管对铁蛋白功能作用的研究日益增多,但其在牙齿发育中的作用及影响尚未见报道。
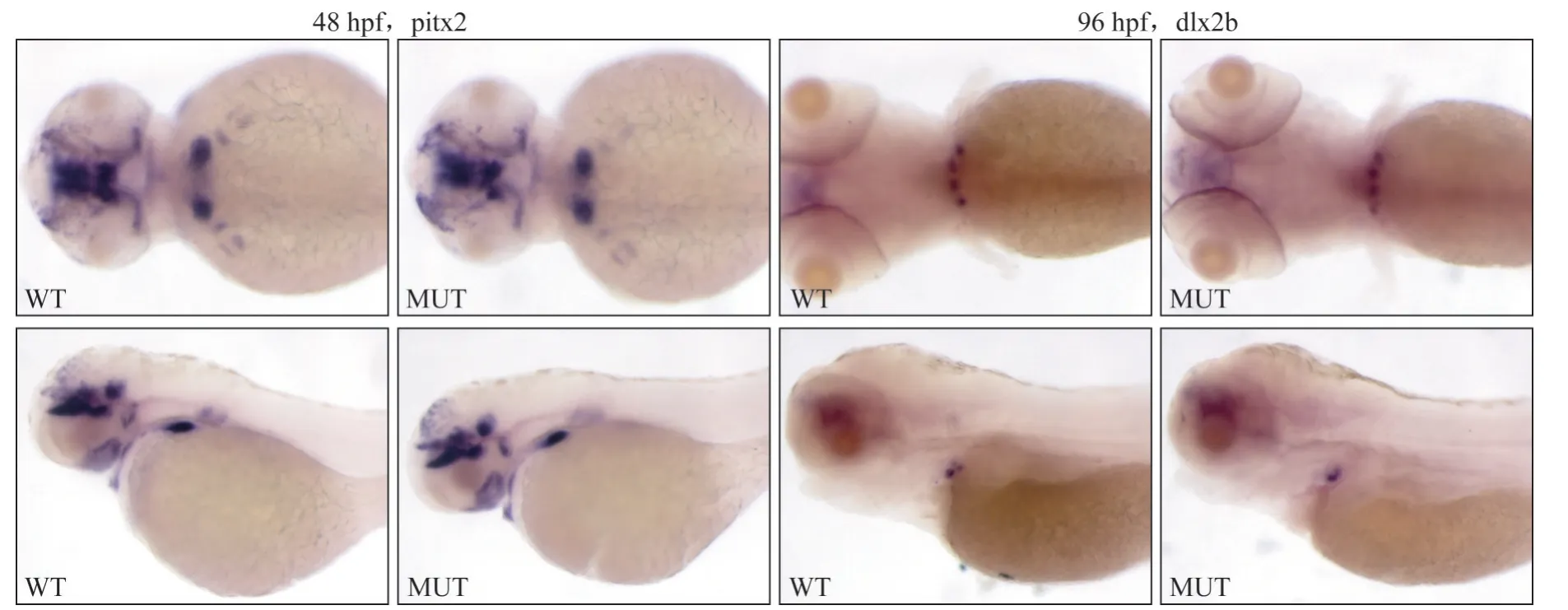
图4 基因pitx2、dlx2b在WT与MUT斑马鱼咽齿的表达情况Fig 4 The expression of gene pitx2 and dlx2b in WT and MUT embryos

图5 120 hpf时斑马鱼咽齿数量、形态及矿化情况Fig 5 The number,shape and mineralization of zebrafish teeth at 120 hpf
早期报道表明,多个基因可以特异性标记斑马鱼咽齿,其中pitx2、dlx2b 是较为常用的标记基因,pitx2 表达于咽齿上皮,是斑马鱼咽齿最早表达的基因,标志着斑马鱼咽齿起始发育;dlx2b 表达于间充质与上皮,是神经棘细胞来源。在本实验中,本课题组首次发现铁蛋白重肽基因fth1b 与dlx2b 在斑马鱼的表达模式相同,特异性表达于咽齿。fth1b 基因特异性表达于斑马鱼咽齿部位,极有可能对斑马鱼咽齿的发育过程发挥作用,因此课题组首次尝试使用CRISPR/cas9基因敲除技术特异性敲除该基因,在可稳定遗传fth1b-/-突变体中,检测发现,与野生型斑马鱼相比,突变体斑马鱼咽齿pitx2和dlx2b的表达均无较大差异,但茜素红染色及共聚焦显微镜观察显示咽齿矿化减弱,意味着突变体斑马鱼咽齿起始与形态形成过程正常而早期矿化受到影响。目前对牙齿发育的大量研究集中于牙齿的发生、形态形成及干细胞探索等方面,对于牙齿矿化的研究相对较少,但是在生活中,矿化缺陷是一种危害极大的牙齿疾病,例如遗传性乳光牙本质及釉质发育不全,牙齿极易磨损暴露牙髓,从而导致炎症发生,严重影响患者生活质量。本实验首次发现基因fth1b 在斑马鱼咽齿表达且对牙齿矿化有作用,为以后更深入探索牙齿矿化及相关疾病提供了又一证据。
利益冲突声明:作者声明本文无利益冲突。
猜你喜欢
农业机械学报(2022年7期)2022-08-08
安徽医科大学学报(2022年6期)2022-07-13
口腔医学(2021年10期)2021-12-02
现代临床医学(2021年5期)2021-11-02
小天使·二年级语数英综合(2021年8期)2021-08-16
能源工程(2021年3期)2021-08-05
食品与健康(2020年3期)2020-03-28
农产品加工(2019年8期)2019-01-06
家庭用药(2016年7期)2016-05-14
作文评点报·作文素材小学版(2016年8期)2016-03-16
