
基于matK和rbcL序列的石斛属植物亲缘关系研究
2021-02-25 09:25林晓霞李会丽郭新红印丽萍
生物学杂志 2021年1期
林晓霞,鹿 炎,李会丽,周 苹,郭新红,印丽萍
(1.湖南大学 生物学院 植物功能基因组学与发育调控湖南省重点实验室, 长沙410082;2.上海市出入境检验检疫局, 上海 200135)
铁皮石斛DendrobiumofficinaleKimura et Migo属于兰科(Orchidaceae)石斛属(DendrobiumSw.)多年生草本植物,是中国古代典籍中记载最早的兰科植物之一[1]。石斛属是兰科中最具药用价值的种属,其茎富含的氨基酸、石斛碱等成分,有抗肿瘤、抗氧化、增强免疫力、降血糖等作用[2-5]。该属约1 000种,中国有74种和2变种,其中细茎石斛D.moniliforme(L.)Sw.、铁皮石斛D.officinaleKimura et Migo、梳唇石斛D.strongylanthumRchb.f.、美花石斛D.loddigesiiRolfe、钩状石斛D.aduncumLindl.等是中药“石斛”的原植物[1]。野生的铁皮石斛生长于海拔达1 600 m的山地半阴湿的岩石上,主要分布在中国安徽西南部、浙江东部、福建西部、广西西北部、四川、云南东南部等地[6]。铁皮石斛对于其生长环境要求比较严格,喜温暖湿润气候和半阴半阳的环境,且不耐寒[7]。由于其生长缓慢、繁殖率低,以及人工过度开采,已被列为国家重点二级保护的珍稀濒危植物。铁皮石斛现已进入大规模人工栽培,但石斛属植物种间形态相似,种类多,其杂交种属也较多[8]。目前市场上铁皮石斛的栽培品种多,种质混杂,石斛属药材的质量也参差不齐。故对铁皮石斛的种质资源进行分析鉴定,对维护铁皮石斛的种质资源具有重要意义,也为市场上铁皮石斛的药材质量提供一定保障。
通过基因组特定区域有足够变异的、易扩增且相对较短的DNA序列作为分子标记[9-11],可以在物种水平上快速准确地鉴定物种[8-9]。该技术目前被广泛地应用于传统中药材的分子鉴定中[12-17]。相比于药材性状鉴定、药材理化鉴定等方法,通过DNA序列标记技术在鉴定药材上不依赖于药材性状,重复性好且准确度高。植物叶绿体基因组通过母系遗传给后代,结构简单,分子量小,序列保守,并且在不同的物种中扩增叶绿体片段的引物是通用的[7,14],使得叶绿体基因被广泛地用作分子标记应用于物种鉴定及系统进化分析中[18]。基于叶绿体matK和rbcL基因评价药用植物物种鉴定现已被应用于许多药用植物中[19-21]。有科学家基于基因matK或rbcL对兰科许多科或亚科的系统发育关系进行了研究[15, 22-24],结果显示matK和rbcL序列能很好地对兰科植物进行鉴定。本研究采用叶绿体基因matK和rbcL序列对不同地区铁皮石斛及已报道的属内近缘物种进行分析,探索石斛属进化亲缘关系,为石斛属种质资源鉴别、遗传多样性保护和遗传育种提供重要的分子依据。
1 材料与方法
1.1 材料
搜集中国6个省份的22份铁皮石斛种质资源,其中湖南省新宁县所采集的样品为野生品种,其余产地均为栽培品种,种质编号及来源地如表1所示。样品采自中国铁皮石斛主要分布地,取样时,对不同产地的植物随机取2~3个植株新鲜叶片,用酒精清洗叶片表面,再用蒸馏水洗去酒精,标记好后迅速用液氮低温冷冻,然后置于-80 ℃冰箱中保存备用。

表1 铁皮石斛采样地基本信息
1.2 方法
1.2.1 样品 DNA 的提取与扩增
将收集的材料先用CTAB-free缓冲液处理,再按照植物基因组DNA提取试剂盒步骤进行DNA的提取。植物DNA提取试剂盒购于天根生化科技(北京)有限公司。将提取的DNA进行浓度和纯度测定,调整最终质量浓度为50~100 μg/μL。matK和rbcL基因扩增引物如表2[25]。其PCR扩增反应体系为20 μL,2×Taqbuffer Mix 10 μL,ddH2O 8 μL,10 nmol/L引物F 0.5 μL, 10 nmol/L引物R 0.5 μL,DNA模板1 μL。扩增程序:95 ℃预变性5 min;95 ℃变性30 s,50 ℃退火 60 s,72 ℃延伸90 s,35个循环;最后72 ℃再延伸10 min。
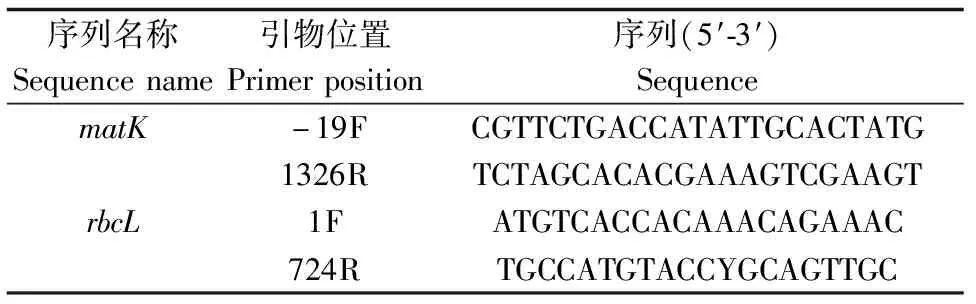
表2 PCR扩增引物
1.2.2 叶绿体基因matK和rbcL序列测定
叶绿体基因matK和rbcL序列的PCR扩增产物经质量分数为1.0%琼脂糖电泳和紫外分光光度法检测质量和浓度后,由湖南擎科生物技术服务有限公司进行双向测序。
1.2.3 数据处理
将获得的测序拼接结果,通过blastn进行序列比对。运用NCBI的Taxonomy查找铁皮石斛近缘物种的叶绿体基因matK和rbcL序列,选取了已报道的铁皮石斛近缘物种的基因序列,将其与样品测序的序列进行比对,利用MEGA 7.0软件计算序列长度、GC含量、变异位点等参数。根据Kimura 2-Parameter(K2P)模型,采用最大似然法(Maximum likelihood,ML)构建系统发育树,利用bootstrap(1 000次重复)检验各分支的支持率。在DnaSP v5中进行多态性位点、遗传多样性指数和中性检测(Fu′s Fs和Tajima′s D)。
2 结果与分析
2.1 基因扩增
成功提取了22份铁皮石斛基因组DNA,PCR扩增基因matK和rbcL后将样品进行琼脂糖凝胶电泳检测(图1和图2)。22条matK序列和rbcL序列的扩增信息如表3所示。matK和rbcL序列均扩增成功,将检测成功的样品送测序公司进行双向测序,校对拼接后,得到44条DNA序列。测序得到的序列长度与琼脂糖凝胶电泳检测的序列长度均一致。
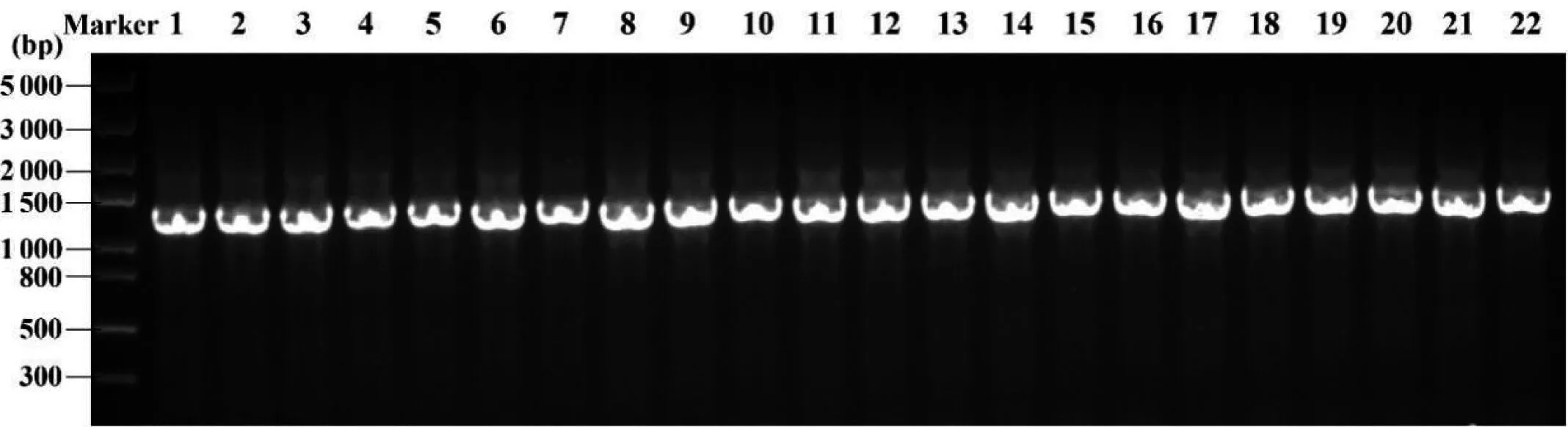
Marker:DNA分子量标准;1~22:样品叶绿体基因matK的目的片段,约为1 300 bp

Marker: DNA分子量标准; 1~22:样品叶绿体基因rbcL的目的片段,约为700 bp
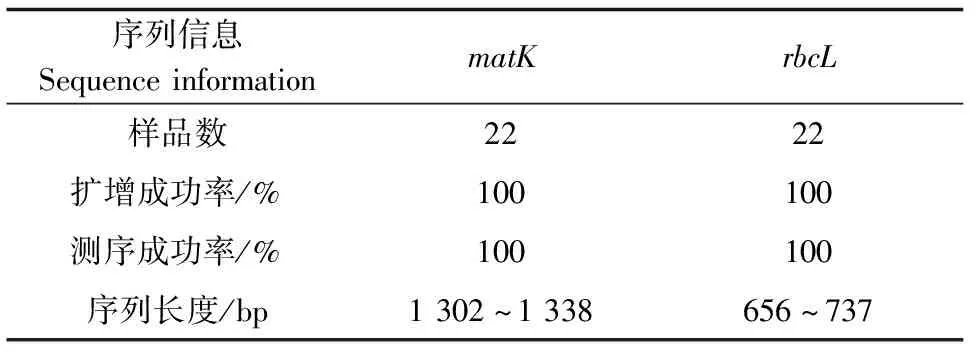
表3 matK和rbcL扩增序列信息
2.2 采集样品结合已报道的叶绿体基因 matK和rbcL序列特征分析
matK序列核苷酸构成中,AT含量为 66.8%~69.2%,GC 含量为 30.8%~33.2%,平均GC含量为32%,远低于AT含量;当空位(gap)作为缺失处理时,序列检测到变异位点数共929个,保守位点数768个,其中简约信息位点871个;rbcL序列核苷酸构成中AT碱基含量为55.6%~59.1%,GC含量为40.9%~44.4%,GC平均含量为42.9%,比AT含量略低;当空位(gap)作为缺失处理时,rbcL序列检测到变异位点数共 506个,保守位点802个,其中简约信息位点24个(表4)。
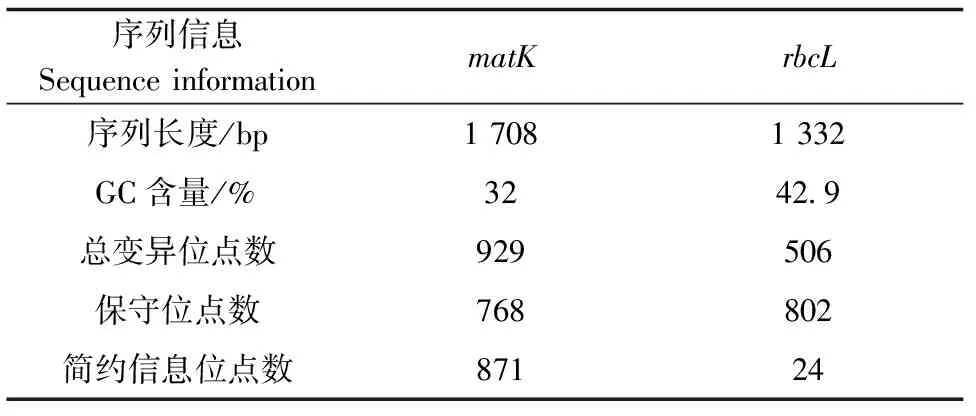
表4 matK和rbcL序列特征
2.3 采集样品结合已报道的叶绿体基因matK和rbcL序列的倍型多态性分析和中性检测
采用DnaSP 5.10[25]对matK及rbcL序列进行倍型多态性分析和中性检测[26]。倍型多态性分析包括单倍型数量(number of haplotypes,H)、单倍型多样性(haplotype diversity,Hd)、核苷酸多样性(nucleotide diversity,π)、单倍型多样性方差(variance of haplotype diversity,Vh)、单倍型多样性标准差(standard deviation of haplotype diversity,Sh)等。结果表明:石斛属matK序列共有H32个,Hd值0.842,π值0.270 16,Vh值0.002 56,Sh值0.051;rbcL序列共有H 17个,Hd值0.664,π值0.026 11,Vh值0.005 56,Sh值0.075。可见,石斛属物种具有较高的遗传多样性。通过DnaSP 5.10 对matK及rbcL序列进行中性检验分析,结果表明:matK序列的Tajima′s D值、Fu and Li′s D*值、Fu and Li′s F*值检验均为正值,在P>0.10水平上不具有显著性;rbcL序列的Tajima′s D值、Fu and Li′s D*值、Fu and Li′s F*值检验均为负值,且在P>0.10水平上具有显著性,符合中性进化模式。结果表明从物种水平上石斛属的rbcL序列进化模式符合中性进化的假设。
2.4 不同产地石斛属叶绿体基因matK和rbcL进化分析
铁皮石斛常用的拉丁学名有3个,分别为D.candidum、D.officinale和D.catenatum,中国植物志所用学名为D.officinale和D.candidum[1],研究的两个学名均有被采用;国外The Plant List网站(http://www.theplantlist.org/tpl1.1/record/kew-57343)铁皮石斛的拉丁学名为D.catenatum[27]。获取已报道的铁皮石斛近缘物种序列包括黄石斛(D.tosaense)、始兴石斛(D.shixingense)、矮石斛(D.bellatulum)和玫瑰石斛(D.crepidatum)等。采用最大似然法(ML)对所有石斛属的matK和rbcL序列进行聚类分析并构建遗传关系树,如图3和图4,图中红色线为样品所在支,蓝色线为与样品同一分支的已报道物种所在支。由图3分析可知,matK序列构建的ML进化树分为两大支,样品序列为主要一支,已报道石斛属物种为一支。样品序列与已报道的D.officinale(铁皮石斛)、D.tosaense(黄石斛)和D.acinaciforme(剑叶石斛)的matK序列为同一支,证明样品为铁皮石斛;除样品编号为AHLA3和AHLA4的序列和已报道的D.officinale、D.tosaense与其他的石斛种属处于同一支外,不同产地的铁皮石斛聚为一类且来自同一产地的居群都聚集到一起。

红色线代表采集样品所在分支;蓝色线代表与采集样品近缘的已报道物种的分支
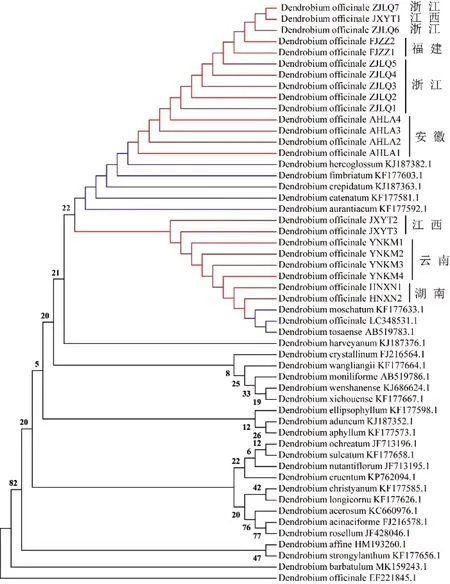
红色线代表采集样品所在分支;蓝色线代表与采集样品近缘的已报道物种的分支
对rbcL序列构建的ML进化树分析表明,同样地,不同产地的铁皮石斛聚为一类且来自同一产地的居群都聚集到一起。样品序列与几个近缘物种主要分为一大支,样品序列可以很清楚地与其他石斛属序列分开。这一大支又再分为两支,其中来源为江西、云南及湖南的样品与已报道的D.officinale、D.tosaense、杓唇石斛(D.moschatum)的序列处于同一支,而来源为福建、浙江、安徽的样品与重唇石斛(D.hercoglossum)、流苏石斛(D.fimbriatum)、玫瑰石斛(D.crepidatum)、黄花石斛(D.catenatum)和线叶石斛(D.aurantiacum)处于一支。
3 讨论
石斛属的下级分类有13组,其中石斛组的类群茎细而花小,具有重要的药用价值[28]。野生石斛的生长环境要求较高,且由于人工采摘过多,目前野生石斛的数量很少,基本都是通过无性繁殖方式进行人工栽培[29-30],品种较杂。本研究样品的采集点是目前铁皮石斛主要栽培的地方,有浙江、安徽、云南、福建、江西及湖南等6省。Asahina等[22]通过叶绿体基因matK和rbcL序列对石斛属的5个种进行鉴定,结果显示matK基因能很好地对其5个种进行分类,而rbcL由于其序列差异性小,所表现的物种鉴别能力不如matK序列。许多研究表明,rbcL具有很高的通用性,但其序列相比于其他序列保守性高,种间差异不大,用于某些物种分类鉴定的识别度不高,特别是区分中间关系密切的物种[14, 31-34]。综合其他对兰科植物研究所采用的基因,选择叶绿体基因matK和rbcL序列进行鉴定。
从遗传多样性分析以及倍型多态性分析中,matK基因表现出更高的遗传多样性(π= 0.270 16,P<0.01),同样在Fu′s Fs和Tajima′s D检验下,matK基因表现出更高的多样性(π=0.026 11,P<0.001);matK和rbcL序列Tajima′s D均显著背离0,说明石斛属的进化过程为自然选择的过程。同时,在序列分析中,matK的简约信息位点有871个,rbcL的简约信息位点有24个,matK序列的变异远远大于rbcL。综合序列分析及遗传多样性结果,可以得出matK序列相比rbcL序列具有更丰富的遗传多样性。
从利用matK所构建的分子进化树分析可知,所有样品主要聚类为一大支,且与已报道的铁皮石斛序列聚为一支,而rbcL序列所构建的进化树,除了已报道的铁皮石斛还与其他的石斛属聚为一支外,其中有杓唇石斛、重唇石斛、流苏石斛、玫瑰石斛、黄花石斛和线叶石斛。从遗传变异性、序列分析和系统进化树三者的结果分析可以得出:matK序列的变异性相对于rbcL大,但结合两个基因的分析可确定22个样品属于铁皮石斛。两个序列所构建的进化树:相同采集地的序列基本能很好地进行聚类,说明同个采集地点的铁皮石斛种质资源一致,样品来源较均一;matk序列的聚类分析:安徽省的物种属于大别山脉,其3个样品,估计受环境影响较大,存在较大的变异;rbcL序列的聚类分析:样品序列的一支又可分为福建、浙江、安徽一大支和江西、云南、湖南另一大支,表明福建、浙江和安徽3个省份的铁皮石斛和江西、云南和湖南的铁皮石斛种质来源都较均一。其中浙江来源样品ZJLQ6与福建的居群聚类结果更近,ZJLQ7与江西的居群更近,且都与浙江其他居群样品距离较远,福建、江西、浙江属于3个毗邻省份,说明浙江省份的部分铁皮石斛居群可能来源于福建和江西。从rbcL序列结果可以得出,除浙江、江西外,其他省份的铁皮石斛种质资源首先聚在一起,表明是一自然物种。但是,来源于浙江的ZJLQ6和ZJLQ7与来源于江西的JXYT1在matK和rbcL序列的进化分析中均聚集一起,说明这3个品种的铁皮石斛亲缘关系较近,该结果与原产地结果划分不完全相符;从matK序列结果可知,相同省份的铁皮石斛居群存在一定的变异及进化,这可能与不同地区的地理环境因素或者品种来源存在一定关系。
本研究通过matK和rbcL序列,结合石斛属序列数据,鉴定了中国6个不同铁皮石斛群居。该研究也为石斛属种质资源鉴别、进化及分类研究提供了参考依据。
猜你喜欢
中国甜菜糖业(2022年4期)2023-01-21
草堂(2022年4期)2022-11-11
贵州林业科技(2022年3期)2022-09-12
数学大王·中高年级(2020年12期)2020-12-31
趣味(语文)(2020年4期)2020-07-27
少儿科技(2019年9期)2019-09-10
中成药(2018年5期)2018-06-06
现代园艺(2018年2期)2018-03-15
安徽农业科学(2015年12期)2015-12-22
中国烟草学报(2012年2期)2012-04-09
